
Abstract
Low-moisture foods (LMF) have clear advantages with respect to limiting the growth of foodborne pathogens. However, the incidences of Bacillus species in LMF reported in recent years raise concerns about food quality and safety, particularly when these foods are used as ingredients in more complex higher moisture products. This literature review describes the interlinked pathways of sporulation and biofilm formation by Bacillus species and their underlying molecular mechanisms that contribute to the bacteriums' persistence in LMF production environments. The long-standing challenges of food safety and quality in the LMF industry are also discussed with a focus on the bakery industry.
Introduction
Low-moisture foods (LMF) are either naturally low in moisture or produced by subjecting foods to dehydration or desiccation, addition of solutes (salt or sugar), among others. They are shelf-stable from a microbiological perspective as the phenotypes necessary to support growth and proliferation are not expressed in these matrices (Beuchat et al., 2013). Baking protocols are associated with loss of water, so the remaining moisture (∼38%, a w circa 0.95–0.96) primarily comes from the interaction of low water activity ingredients (a w < 0.85) such as flour, herbs, spices, seeds, dried milk, protein powders, and nuts during processing (Schmidt and Fontana Jr., 2007; Cauvain and Young, 2008; Anon., 2010; Ishida and Steel, 2014; Gurtler et al., 2019). It has been established that LMF provide a conducive environment for long-term survival of microorganisms without the necessity for them to grow within a food matrix to be considered a risk (Bourdichon et al., 2021).
Studies suggest that more than one million food-associated illnesses per year are caused by bacterial toxins, including Bacillus (Jessberger et al., 2020); however, the latter is often underreported because of usually mild and self-limiting symptoms (Dierick et al., 2005). A recent study designated Bacillus species as one of the leading causes of food poisoning (Glasset et al., 2016). Although outbreaks of foodborne illness traced to LMF have been recognized for some time, these are now being increasingly reported due to improved detection methods (Logan, 2012; Choi and Kim, 2020). The Centers for Disease Control described 619 confirmed outbreaks of Bacillus-related poisoning from 1998 to 2015 (McDowell et al., 2022). A total of 105 outbreaks in 2005–2007 in the European Union caused by Bacillus toxins were linked to a range of LMF (EFSA, 2009). The European Commission Rapid Alert System for Food and Feed issued 28 alerts for the presence of Bacillus species in LMF during 2015–2021 (EC, 2021).
The genus Bacillus consists of Gram-positive bacterial species belonging to phylum Firmicutes, which display a wide range of genotypic and phenotypic characteristics (Postollec et al., 2012; Berendsen et al., 2016; Soni et al., 2016). Their genomes have low G + C content having less than 50% guanine (G) and cytosine (C) bases in their DNA (Traag et al., 2013), facilitating a unique ability to temporarily and reversibly transform into highly resistant structures called endospores under stressful conditions (Piggot and Hilbert, 2004; Voundi et al., 2017; Ulrich et al., 2018).
Bacillus species are ubiquitous and have been frequently reported in raw materials including flour, brewer's yeast, bread improvers, spices, and herbs (Pepe et al., 2003; Smith et al., 2004; Logan, 2012; Lavermicocca et al., 2016). In addition, they can also form three-dimensional colony biofilms allowing them to persist in adverse conditions (Lavermicocca et al., 2016), resulting in the contamination of food products (Bridier et al., 2013). The resistance and heterogeneous response of Bacillus species to preservation treatments such as heat, radiation, organic acids, among others, pose major challenges in predicting their spoilage and disease-causing potential (Eijlander et al., 2011).
While baking kills most vegetative bacteria (Lavermicocca et al., 2016), and the water activity of the bread crust is normally low enough to limit microbial growth, conditions in the loaf center and storage at ambient temperatures are more suited to support the growth of Bacillus species (Cauvain, 2017). The fermentation before baking, such as sponge dough for 4–8 h to overnight as well as sourdough starters stably maintained for years, could facilitate Bacillus spores (Deák, 2003; Cauvain, 2015; Landis et al., 2021, personal communication with artisan bakery associated with this research). It is apparent that to control Bacillus species in LMF, understanding their survival mechanisms is key. Therefore, this review aims to explore the underlying molecular mechanisms of sporulation and biofilm formation expressed by Bacillus species, which contribute to their ecological niche persistence.
Survival Mechanisms of Bacillus Species in LMF Built Production Environments
The natural and unique tendency of Bacillus species to adapt to stresses, accomplished by altering their gene expression to develop into endospores, is interlinked with their ability to form biofilms with sporulating cells often embedded within them, either as Bacillus-only biofilms or as heterogeneous biofilms with microorganisms from other genera. When compared with planktonic cells, Bacillus spores and biofilms exhibit an enhanced resistance to processing treatments and exposure to low water activity confers cross-tolerance to other stresses, including resistance to biocides, UV irradiation, heat tolerance, among others (Simões et al., 2010; Madigan et al., 2014; Remize, 2016).
Sporulation and biofilm formation could be described as the two main survival mechanisms of Bacillus species in LMF production environments. Earlier studies have proposed sporulation as a culminating feature of biofilm formation wherein motile cells transit to matrix-producing cells ultimately developing into spore-producing cells (Vlamakis et al., 2008).
Sporulation and contributing environmental conditions
Sporulation, the process of transforming from vegetative bacteria into endospores, is a survival strategy triggered by unfavorable environmental conditions generated during food processing, such as low moisture/water activity in case of LMF, in which the former alters its genotypic expression to form dormant endospores (Stragier and Losick, 1996; Tan and Ramamurthi, 2014; Gauvry et al., 2017). Endospores are tough, refractile, dehydrated, and metabolically inert structures, highly resistant to physicochemical agents and preservation treatments, including osmotic pressure, extreme heat or cold, starvation, action of biocides, UV radiation, and which once formed can remain dormant for long periods (Brown, 2000; Setlow et al., 2006; Black et al., 2007; Wei et al., 2010; Postollec et al., 2012; Madigan et al., 2014; De Bellis et al., 2015; André et al., 2017; Ulrich et al., 2018).
When water becomes available as occurs during food preparation and reconstitution, spores germinate into vegetative cells leading to spoilage or food poisoning (Baril et al., 2012; Logan, 2012; Markland et al., 2013; Hertwig et al., 2015; Oh et al., 2015; Fogele et al., 2018; Huang et al., 2020). Subsequent storage at ambient temperature further provides a suitable environment to promote their germination and outgrowth (Jaquette and Beuchat, 1998; Lavermicocca et al., 2016; Cauvain, 2017).
Some Bacillus endospores can survive baking processes (Lindsay and Flint, 2009; Valerio et al., 2015). DT values (time required at temperature T to achieve 90% reduction in spore count) reported previously are in line with survival at standard sterilization processes used (121°C) in the food industry; an average D 120 = 0.33–45.7 s was recorded for Bacillus subtilis (Berendsen et al., 2015b), D 110 = 0.3–0.7 min for Bacillus cereus (Stoeckel et al., 2013), D 120 = 1.3–1.9 min for Bacillus thermoamylovorans (Berendsen et al., 2015a), and D 135 = 0.02 min for Bacillus licheniformis (Janštová and Lukášová, 2001).
Furthermore, Bacillus species demonstrate an increased resistance to heat in low a w conditions (a w circa 0.34) to as high as D 120 = 2 h (Fan et al., 2020). The resistance demonstrated by endospores underpins their ability to persist in LMF production environments. Even though it is known that sporulation can occur at multiple stages along the complex food chain (Turner et al., 2006; King et al., 2007; Heyndrickx, 2011; Di Pinto et al., 2013; Ijabadeniyi and Pillay, 2017; Sánchez-Maldonado et al., 2018), the precise sporulation niches along the farm-to-fork continuum are challenging to identify (Gauvry et al., 2017). Moreover, the conditions required for spore formation in their natural habitat are often unknown; therefore, estimating germination and outgrowth is complicated within food production settings (Wells-Bennik et al., 2016).
Sporulation cycle in Bacillus species
The main stimulus required to trigger sporulation is starvation. Formation of an endospore from vegetatively growing Bacillus follows a series of well-defined steps (summarized in Fig. 1). Under stressful conditions, an asymmetrically positioned septum is created followed by the mother cell engulfing the smaller part or forespore. This leads to the development of a thick peptidoglycan layer around the chromatin network, known as the cortex. Sporulation culminates with lysis of the mother cell and release of a mature spore (Doi, 1989; Errington, 2003; Setlow, 2003; Black et al., 2007). The time to complete the sporulation cycle varies, ranging from ∼8 to 10 h in B. subtilis (Zhang and Mathys, 2019).
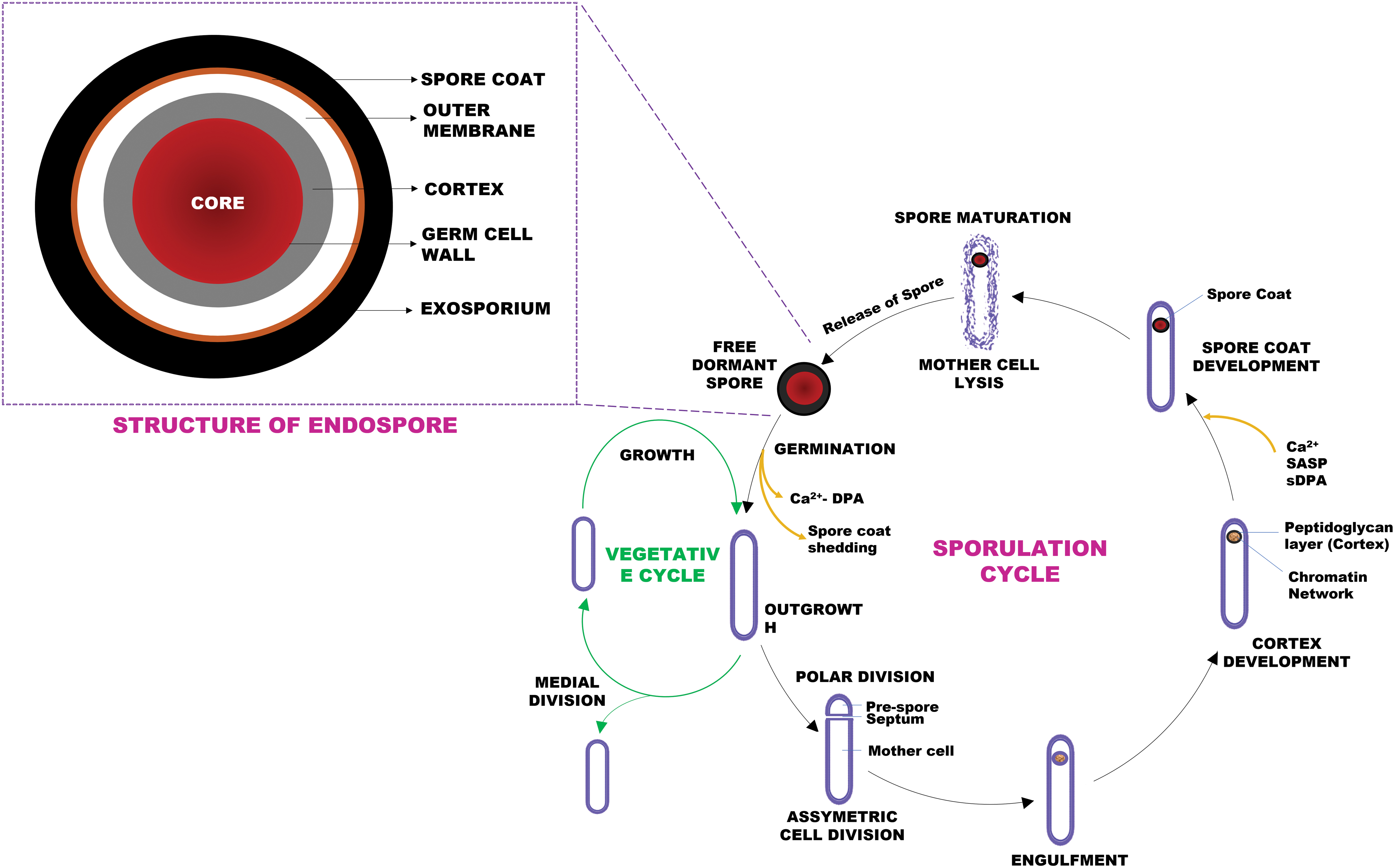
A schematic showing the features associated with the structure of a typical Bacillus endospore along with the key stages involved in the sporulation cycle. The sporulation cycle begins following an environmental cue, with the development of an asymmetric septum to produce a mother cell and a forespore. This step is then followed by the mother cell engulfing the forespore, after which the peptidoglycan cortex is assembled around the chromatin and the spore coat develops during late sporulation. As the spore matures, the mother cell lyses to release it into the environment. The multiple layers and partially dehydrated central core of the spore impart resistance while the exosporium (absent in Bacillus subtilis) aids in the attachment to surfaces. The core is protected by the cortex and spore coat, which consists of an inner coat, outer coat, and crust. These spores are capable of germinating and resuming vegetative growth in response to nutrients.
The external layer on spores produced by B. cereus and Bacillus megaterium is a proteinaceous exosporium that plays an important role in surface attachment (Stewart, 2015). B. subtilis does not express an exosporium (Setlow, 2014a), rather it has a triple-layered, semipermeable proteinaceous structure consisting of an outer coat (abundant in electrons), an inner coat (thinner hence termed a crust), and an electron-diffuse undercoat, which together confer resistance to chemicals and enzymes while providing structural integrity and facilitating surface attachment (Setlow et al., 2006; Black et al., 2007; McKenney et al., 2013).
Cortex forms the layer below the exosporium or spore coat and acts as a protective cover for the core. The central core consists of ribosomes, inert enzymes, and small acid-soluble binding proteins (SASPs) that bind tightly to DNA (chromatin network) and protect it from ultraviolet radiation, desiccation, and dry heat (Doi, 1989; Stragier and Losick, 1996; Setlow et al., 2006; Madigan et al., 2014; Soni et al., 2016; Ulrich et al., 2018). The low water content (25–55% of wet weight) and high levels of pyridine-2,6-dipicolinic acid (DPA) in the spore coat along with α/β-type SASPs provide resistance against heat and chemicals (Setlow et al., 2006; Madigan et al., 2014; Setlow, 2014b).
During germination or spore revival, described in the context of B. subtilis, Ca2+ and dipicolinic acid complex (Ca2+-DPA), representing 10% of the dry weight of the endospore, is released to rehydrate the core followed by disintegration of the peptidoglycan layer or cortex lysis and expansion of the germ cell wall. SASPs function as a carbon and energy source for the outgrowth of a new vegetative bacterial cell from the endospore during germination.
Upon rehydration, enzyme activity is restored, RNA and other macromolecules synthesized, and the spore coat disintegrates, leading to outgrowth and re-entry into the vegetative growth cycle (Errington, 2003; Setlow, 2003; Moir, 2006; Eijlander et al., 2014; Madigan et al., 2014). Although metabolically inactive, spores can sense their surroundings and rapidly respond to the presence of appropriate nutrients by germinating and resuming the vegetative state, occasionally expressing a pathogenic phenotype due to the production of toxins thus compromising shelf-life and food safety (Setlow, 2003; Postollec et al., 2012; Markland et al., 2013; André et al., 2017; Voundi et al., 2017).
Spore germination can be triggered by nutrients in the food matrix (amino acids:
Some Bacillus spores have fewer GR on their inner membrane, and therefore, the germination is either not induced or occurs slowly (Setlow, 2003). Such spores are known as superdormant spores, and their germination can only be induced by heat treatments 8–15°C higher than the normal activation temperatures (60–100°C) or by exposure to pressures of around 500 MPa or utilizing the germination-inactivation strategy by triggering the germination of spores by exposure to a pressure of 150 MPa at 37°C in the first step followed by their elimination with a milder inactivation step (Ghosh and Setlow, 2009; Wei et al., 2010; Markland et al., 2013; Zhang and Mathys, 2019; Delbrück et al., 2021).
Interestingly, under food processing conditions, bacterial cells form biofilms before sporulation, by forming sessile communities encased in a protective self-produced extracellular matrix (ECM) (Vlamakis et al., 2008; López et al., 2009a; Faille et al., 2014; Oh et al., 2015; Galié et al., 2018). These biofilms eventually act as a nidus for sporulation and subsequent release into the production environments (Hamon and Lazazzera, 2001; Wijman et al., 2007).
Biofilm formation and some of their functional features
Formation of biofilms by bacteria is a survival mechanism offering self-defense with inherent tolerance against desiccation, mechanical strength against liquid flows in pipelines, resistance against antimicrobial compounds and biocides, and adaptation to common food processing stresses such as temperature variations, radiation, among others (Kobayashi and Iwano, 2012; Oh et al., 2015). Those biofilms formed by Bacillus on surfaces, such as food preparation equipment, conveyor belts, tanks, vats, pipes, corners, cracks, crevices, gaskets, valves, and transport vehicles, are not only difficult to prevent but are also hard to eliminate through various cleaning interventions.
These bacterial formations constitute a risk toward recontamination of food processing lines along with the final product, while confounding food processing/preservation protocols (Bailey and von Holy, 1993; Giffel et al., 1997; Simmonds et al., 2003; Auger et al., 2006; Brooks and Flint, 2008; Kobayashi and Iwano, 2012; Faille et al., 2014; Farakos and Frank, 2014; Gopal et al., 2015; Fink et al., 2017; Møretrø and Langsrud, 2017; Galié et al., 2018). In case of bakery facilities, equipment including mixers, moulders, and slicers, among others, may be particularly challenging to clean as there is high possibility of dough and flour to build up, aiding in biofilm formation and transfer to the finished products (Brouillette and Haley, 2019).
In addition to cross-contamination, biofilms also contribute to the corrosion of pipelines and tanks, shortened time between cleaning, process downtime, reduced system/equipment efficiency, or even obstruction (Ryu and Beuchat, 2005; Cappitelli et al., 2014). The adhered bacterial communities may be resident or transient with a biotransfer potential (Wirtanen et al., 1996; Verran, 2002; Pereira et al., 2008; Møretrø and Langsrud, 2017).
Studies demonstrate that Bacillus modifies its biofilm composition and architecture in terms of volume, wrinkled network, and spatial expansion, as a stress-adaptive response (Stanley et al., 2003; Majed et al., 2016; Gingichashvili et al., 2019). Biofilms exhibit quorum sensing (QS) while existing in close proximity to augment cell-to-cell communication as well as nutrient and genetic exchange using chemical signaling molecules/autoinducers (Vlamakis et al., 2008; Cappitelli et al., 2014; Faille et al., 2014; Madigan et al., 2014; Akbas, 2015; Oh et al., 2015; Galié et al., 2018). This feature enables the development of unique spatiotemporal patterns both in single population Bacillus biofilms and in those with mixed populations including members such as Staphylococcus, Listeria, Pseudomonas, and others (Auger et al., 2006; Hsueh et al., 2006; Jahid and Ha, 2014; Mielich-Süss and López, 2015; Majed et al., 2016).
Stages of biofilm development in Bacillus species
Biofilm formation occurs following a series of well-defined sequential steps (summarized in Fig. 2) beginning with attachment (reversible and irreversible), cell-to-cell adhesion, expansion, maturation, and dispersal (Cappitelli et al., 2014; Akbas, 2015; Coughlan et al., 2016). At the initiation of biofilm formation, motile cells with flagellae differentiate into nonmotile cells to form microcolonies, which then adhere to each other and to the surface by secreting a slimy matrix composed of extracellular polymeric substances (EPS)—cellulose, proteins, lipids, and extracellular DNA—forming a single layer or three-dimensional structure essential for biofilm integrity (Branda et al., 2006; Marvasi et al., 2010; Vlamakis et al., 2013; Akbas, 2015; Peng et al., 2020).
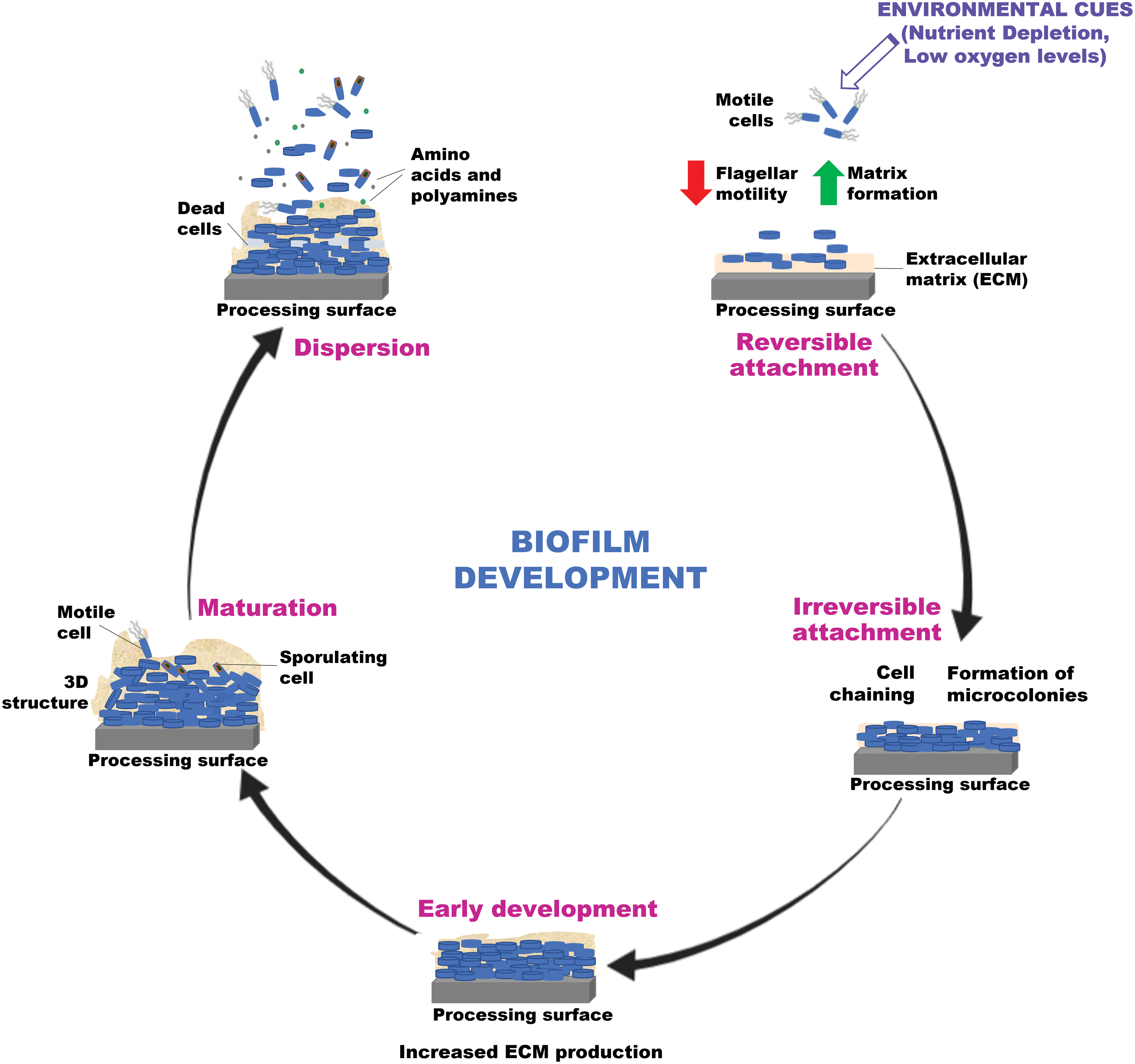
A schematic showing the five steps involved in the development of Bacillus biofilms on food processing surfaces. Biofilm development occurs in five stages beginning with attachment (reversible and irreversible) of bacteria to a surface, cell-to-cell adhesion, expansion, maturation, and finally disassembly of the community. Motile cells expressing flagella differentiate into chains of nonmotile matrix-producing bacterial cells that attach to the surface and a protective ECM starts to develop around these cells. Upon maturation, some matrix-producing cells sporulate while the ECM production enhances. This further leads to a community of matrix-producing cells, sporulating cells, and dead cells while small molecules such as
Bacillus species exhibit appendages on their exosporium surface for surface attachment that functions to embed them in the EPS (Klavenes et al., 2002; Van Houdt and Michiels, 2010). Motile cells positioned on the bottom layer of biofilm contribute to the expansion of the bacterial community, whereas matrix producers in the core primarily act to produce the ECM and maintain its rigidity (Vlamakis et al., 2008). This matrix also provides shelter to the inner cells, contributing to the recruitment of nutrients and adhesion to the surface (Alvarez-Ordóñez et al., 2019). The biofilm often elaborates extensive wrinkles to increase the surface-to-volume ratio providing for better access to oxygen (Mielich-Süss and López, 2015).
Upon maturation, some Bacillus cells secrete
Furthermore, as a defense mechanism in natural settings, a subpopulation of B. subtilis differentiate into cannibals (competent cells) to ensure that the matrix producers thrive using the nutrients released by lysed cells sensitive to environmental stress, ultimately increasing matrix production and delaying sporulation (López et al., 2009b). Thus, motile cells, nonmotile matrix producers, competent cells, QS cells, and sporulating cells all coexist in a biofilm (López et al., 2009a).
Despite the research undertaken to date, the food industry continues to be challenged in controlling (Bacillus) biofilm development and sporulation because of the complex nature of their formation in niche environments.
Molecular Regulation of Sporulation and Biofilm Formation in Bacillus Species
The molecular mechanisms underpinning biofilm formation, disassembly, and sporulation in B. subtilis as a model system have been extensively studied. The relevant molecular pathways involved in the sporulation cycle and biofilm formation in Bacillus species are summarized in Figure 3.

A schematic figure showing a representation of the regulatory pathways functioning in biofilm (matrix) formation and sporulation in Bacillus species. The schematic representation presents the key components for biofilm formation and sporulation in Bacillus species denoted by transcriptional activation (), transcriptional repression ( ), and indirect regulation (
), and indirect regulation ( ,
,  ) along with their phenotypic expression. The master regulator, Spo0A (stimulating the synthesis of σ
H), under stress, is activated by a multicomponent phosphorelay consisting of histidine kinases KinA, KinB, KinC, and KinD and proteins Spo0F and Spo0B—first, at low concentrations via the activation of the regulatory protein SinI to develop ECM encoded by epsA-O, tapA-sipW-tasA, and slrR. AbrB and SinR act as repressor pathways inhibiting transcription of ECM genes. SinI binds to SinR so the latter is unable to block target promoters allowing transcription of target genes. The expression of sinI and σ
H is repressed by the AbrB regulator. Biofilm activation is further promoted when DegU∼P (under the action of ComP-ComA) produces BslA encoding hydrophobic biofilm coat. On reaching a quorum, ComA, Spo0F, and DegU are indirectly repressed. Finally, the biofilm containing cells disperse in the environment probably under the action of σ
B while some enter the sporulation cycle. If starvation conditions persist, sporulation is promoted as a result of higher concentrations of active Spo0A. Once asymmetric division occurs, a cascade of sigma factors is triggered resulting in a discrete expression profile at each step of sporulation. Spo0A∼P works by activating the transcription of several key sporulation-specific genes, particularly the spoIIE, spoIIA, and spoIIG genes as well as successive actions of specific sigma factors σ
F (forespore), σ
E (mother cell), σ
G (engulfed spore), σ
K (mother cell with endospore), and σ
A (germinated cell under the action of germination receptors GerA, GerB, GerK, and GerD releasing Ca2+-dipicolinic acid). ECM, extracellular matrix.
) along with their phenotypic expression. The master regulator, Spo0A (stimulating the synthesis of σ
H), under stress, is activated by a multicomponent phosphorelay consisting of histidine kinases KinA, KinB, KinC, and KinD and proteins Spo0F and Spo0B—first, at low concentrations via the activation of the regulatory protein SinI to develop ECM encoded by epsA-O, tapA-sipW-tasA, and slrR. AbrB and SinR act as repressor pathways inhibiting transcription of ECM genes. SinI binds to SinR so the latter is unable to block target promoters allowing transcription of target genes. The expression of sinI and σ
H is repressed by the AbrB regulator. Biofilm activation is further promoted when DegU∼P (under the action of ComP-ComA) produces BslA encoding hydrophobic biofilm coat. On reaching a quorum, ComA, Spo0F, and DegU are indirectly repressed. Finally, the biofilm containing cells disperse in the environment probably under the action of σ
B while some enter the sporulation cycle. If starvation conditions persist, sporulation is promoted as a result of higher concentrations of active Spo0A. Once asymmetric division occurs, a cascade of sigma factors is triggered resulting in a discrete expression profile at each step of sporulation. Spo0A∼P works by activating the transcription of several key sporulation-specific genes, particularly the spoIIE, spoIIA, and spoIIG genes as well as successive actions of specific sigma factors σ
F (forespore), σ
E (mother cell), σ
G (engulfed spore), σ
K (mother cell with endospore), and σ
A (germinated cell under the action of germination receptors GerA, GerB, GerK, and GerD releasing Ca2+-dipicolinic acid). ECM, extracellular matrix.
Nutrient deprivation triggers several cellular responses including increased motility and chemotactic responses in a search for nutrients while attempting to utilize by-products of growth and other carbon sources to prevent starvation. If these initial responses fail to provide the required nutrients, stress-adaptive responses are triggered (Doi, 1989; Baril et al., 2012). At the center of this regulatory network is the highly conserved Spo0A regulon (Hamon and Lazazzera, 2001; Fujita et al., 2005; Traag et al., 2013), the activity of which is regulated by a phosphorylation network (Stragier and Losick, 1996; Errington, 2003; Piggot and Hilbert, 2004; Liaqat et al., 2013).
The spo0A gene is expressed primarily during the exponential phase of growth (Doi, 1989), a low activation level of which induces matrix (biofilm) production while a higher activation level favors sporulation. Therefore, as summarized in Figure 4a and b, when Spo0A is initially phosphorylated, biofilm formation commences as a consequence of matrix gene expression, and upon maturation, Spo0A∼P accumulates in certain cells to activate sporulation (Setlow, 2003; Fujita et al., 2005; Vlamakis et al., 2013; Gauvry et al., 2017).

A schematic figure showing a representation of the key molecular factors interlinked in biofilm (matrix) formation and sporulation in Bacillus species:
The phosphorelay cascade begins with the phosphorylation of Spo0F by kinases KinA, KinB, KinC, KinD, resulting in the transfer of a phosphoryl group to Spo0B, which then phosphorylates Spo0A (Spo0A∼P), the active form of the response regulator (Errington, 2003; Piggot and Hilbert, 2004; Vlamakis et al., 2008; Eijlander et al., 2014; Mielich-Süss and López, 2015). Once Spo0A is phosphorylated, the expression of a sigma factor (σ H) is stimulated (López et al., 2009a), which then functions to activate the transcription of other genes required for Spo0A phosphorylation (kinA and spo0A). In this way, the phosphorylation of Spo0A (Spo0A∼P) switches on a cycle that stimulates both its synthesis and its phosphorylation (Chung et al., 1994).
Furthermore, both σ H and Spo0A have been reported to initiate biofilm development in B. subtilis (Hamon and Lazazzera, 2001; Ren et al., 2004). It has been reported that σ B induces the expression of Spo0E whose gene product then inactivates Spo0A∼P by dephosphorylation highlighting the two dominant and probably mutually exclusive adaptive stress responses—biofilm formation and sporulation—within the regulatory network that influences the fate of B. subtilis (Reder et al., 2012; Rodriguez Ayala et al., 2020).
Of the four kinases that phosphorylate Spo0A, KinA and Kin B are essential for spore formation, KinC for biofilm formation while KinD plays a dual role (Hamon and Lazazzera, 2001; Kobayashi, 2008). KinC senses the potassium cation (K+) leakage under stress and triggers the production of ECM to maintain the bacteria within the biofilm network (López et al., 2010). KinB is activated when the electron transport chain is impaired, as would happen during low oxygen availability. KinA triggers the onset of spore formation in response to impaired respiration by sensing a reduction in reducing power via nicotinamide adenine dinucleotide (NAD+/NADH) levels in the cytoplasm (Piggot and Hilbert, 2004; Kolodkin-Gal et al., 2013; Liaqat et al., 2013; Tojo et al., 2013; Mielich-Süss and López, 2015). KinD acts as a checkpoint to sense the production of extracellular polysaccharides, thereby facilitating bacteria to undergo sporulation when a critical threshold is reached (Aguilar et al., 2010). Therefore, depending on which kinase is stimulated, different levels of Spo0A activation occur resulting in a distinctive outcome.
A lower threshold of Spo0A∼P is required for ECM gene expression via activation of the regulatory protein SinI (Fujita et al., 2005). Expression of sinI is repressed by the AbrB regulator, thus negatively regulating biofilm formation by repressing the expression of genes involved in intercellular adhesion (Hamon and Lazazzera, 2001). Both abrB and sinI respond at low levels of Spo0A∼P (Veening et al., 2006). Spo0A represses abrB and AbrB represses σ H, therefore, until a threshold of Spo0A is reached, AbrB represses σ H keeping Spo0A levels low (López et al., 2009a; Reder et al., 2012).
AbrB also represses a variety of genes that are activated at an early stage of sporulation, including spoVG, spo0E, and spo0H (Fujita et al., 2005). It is further reported that AbrB and Spo0E together act as a checkpoint to prevent premature commitment to sporulation (Shafikhani and Leighton, 2004). In addition, the σ B regulatory cascade is reported to control the expression of the transcription factor sinR, which in turn influences whether or not bacteria residing in the biofilm remain in and continue to grow or colonize new niches through biofilm dispersal (Rodriguez Ayala et al., 2020).
SinR, a small DNA-binding transcription factor, acts centrally in the process of cell transitioning between planktonic and sessile states (Kearns et al., 2005; Kobayashi, 2008). Under conditions that favor planktonic growth, SinR activates genes required for flagellum synthesis (hag) while represses biofilm-related operons epsA-O (encodes extracellular polymeric substances or EPS synthesis), tapA-sipW-tasA (encodes protein components that provide structural integrity to the matrix), and slrR (activates biofilm matrix gene expression) (Kearns et al., 2005; Branda et al., 2006; Chu et al., 2008; Kobayashi, 2008; Kampf et al., 2018).
SinR negatively regulates spo0A and is necessary for competence (Mandic-Mulec et al., 1995). At the initiation of biofilm formation, SinR is sequestered by its antagonists SinI and SlrR (slrR expressed in the presence of high levels of SinI), allowing the expression of the eps operon and the production of proteins required for exopolysaccharide synthesis (Vlamakis et al., 2008; López et al., 2009a; Liaqat et al., 2013; Mielich-Süss and López, 2015; Kampf et al., 2018). It is proposed that SlrR determines motility at low activation levels and biofilm formation at high activation levels by downregulating motility functions (Chai et al., 2010).
Furthermore, in B. subtilis, biofilm formation relies on the assembly of a fibrous framework formed by action of TasA, a protein that functions to express amyloid-like fibers protruding from the cell wall (Romero et al., 2014). TapA (TasA anchoring/assembly protein) is required for securing the fibers to the cell wall, while SipW functions as a dedicated signal peptidase and also induces expression of the tapA and eps operons. Interestingly, TapA (formerly annotated as YqxM) is reported to be induced by the action of SlrR to promote biofilm formation; however, the latter is not required for the expression of eps operon (Chu et al., 2008; Kobayashi, 2008).
Moreover, the absence of TapA has been correlated with a reduction in the level of TasA in the biofilm matrix (Romero et al., 2014). The molecular structure of the EPS remains to be elucidated; however, it is known that eps-defective mutants develop flat colonies and fragile pellicles (Branda et al., 2006; Nagórska et al., 2010; Peng et al., 2020). EpsE-mediated inhibition of motility ensures the flagellar motor switch protein FliG functions to switch off motility when matrix production occurs, thereby initiating biofilm formation (Vlamakis et al., 2013).
An additional protein component of the matrix is BslA, secreted during the final stages of biofilm maturation, self-assembles into a hydrophobic layer on the biofilm securing its integrity (Kobayashi and Iwano, 2012). It is possible that EPS mediates hydrophobicity indirectly as a consequence of currently undefined interactions with BslA and that the hydrophobic properties of the biofilm raincoat protein BslA might be involved in TasA fiber formation (Hobley et al., 2015). Together, epsA-O, tapA-sipW-tasA, and bslA are referred to as ECM biosynthetic genes (Pisithkul et al., 2019). Also, sfp required for production of lipopeptide surfactin is essential for swarming that is, movement of a large mass of bacteria on solid surfaces, latter recognized as a preliminary step in biofilm formation in wild-type bacteria (Nagórska et al., 2010). The srfA operon is required for the production of surfactin, competence establishment, and conditionally required for sporulation (Nakano et al., 1991; Kobayashi, 2007, 2008; Mielich-Süss and López, 2015).
Two other proteins, YmcA and YlbF, probably modulate the interaction between SinR and SinI and influence the high-level expression of the eps operon (Kearns et al., 2005; Kobayashi, 2008). It is further suggested that YmcA and YlbF interact with a third protein, YaaT, to form a tripartite complex, which in turn regulates the activity of Spo0A by stimulating the phosphorelay, probably by interacting with Spo0F and Spo0B (Carabetta et al., 2013). Additionally, the phosphodiesterase YmdB in close association with SinR controls the bistable switch between expression of matrix production and downregulation of motility gene expression, including the flagellum protein (hag) and autolysin proteins (Branda et al., 2006; Mielich-Süss and López, 2015; Kampf et al., 2018). The fact that ymdB mutants appear to readily acquire suppressor mutations that restore biofilm formation suggests selective pressure for this behavior (Ryan-Payseur and Freitag, 2018).
Under conditions where nutrient availability is low, but sufficient to allow metabolic activity, low levels of Spo0A trigger the expression of two operons: sporulation-killing factor (skf) and sporulation-delaying protein (sdp) as a mechanism to prolong biofilm formation. In the presence of potential competition by other bacteria within the same niche, the former produces bacteriocins to reduce the susceptible cell population, thereby enabling cannibalization for nutrients, which in turn acts to promote the growth of matrix-producing cells, while briefly delaying sporulation (López et al., 2009a; González-Pastor, 2011; Vlamakis et al., 2013).
In addition to Spo0A, the two-component system DegS-DegU influences biofilm formation by activating ECM (Pisithkul et al., 2019). Modulated by DegS and DegQ, a low level of response regulator DegU∼P (usually consistent with the planktonic state) activates transcription of flagellar genes, whereas a high level represses them via reduction of σ D activity required for transcription of flagellar genes, secretion of exoproteases, and biofilm formation (Kobayashi, 2007). DegU∼P favors bslA transcription in floating biofilms (Kobayashi and Iwano, 2012) and poly-γ-glutamate (pgs) transcription in surface-adhered biofilms (Stanley and Lazazzera, 2005). It has been also shown that DegU indirectly affects Spo0A∼P, shortening the time for phosphorylation, required to trigger the synthesis of the ECM (Marlow et al., 2014).
The expression of DegQ is regulated by another two-component quorum signal response system, ComP-ComA (Kobayashi, 2008). Being a QS regulated process, activation of ComA∼P (hence DegQ) occurs upon reaching a threshold cell density (Simões et al., 2010). Studies have shown that QS in Bacillus species is regulated through the comQXPA locus, mediated by small peptides that control DNA uptake and sporulation (Dogsa et al., 2014), ComA confirmed as the most conserved protein in the QS locus in B. licheniformis (López et al., 2009b).
The transmembrane histidine kinase sensor protein (ComP) interacts with the ComX pheromone leading to phosphorylation of the ComA response regulator/transcription factor, which in turn drives transcription of the srf operon hence epsA-O operon (Esmaeilishirazifard et al., 2017). The phosphorylated form of ComA (ComA∼P) can also induce the expression of the proteins RapA, RapB, and RapE in B. subtilis, which specifically block the phosphorelay route for Spo0A (López et al., 2009a). It was recently postulated that the conserved comER gene might play a role in the regulatory circuit that controls Spo0A activation in B. subtilis and B. cereus (Setlow, 2014a; Yan et al., 2016).
B. subtilis also demonstrates biofilm regulation via luxS-mediated QS system that produces chemical signal molecules called autoinducers (AI-2), the expression of which might be repressed by SinR and Spo0A (Lombardía et al., 2006). A recent study reported on a cross-pathway regulation wherein the biofilm activators SlrR and ComK function to block competent cells from simultaneously becoming matrix producers by repressing the key biofilm regulatory gene sinI (She et al., 2020). Another study suggested that the ComQXPA QS system acts as a switch that contributes to the stochastic sporulation initiation and through this pathway promotes bet-hedging behavior.
This finding is important for quorum-quenching strategies that function to down-modulate biofilm development by inhibiting QS signaling (Ryan-Payseur and Freitag, 2018; Špacapan et al., 2020). Furthermore, the ability of Bacillus species to switch and thus maintain a subpopulation of swimmers through SinR regulation provides the bet-hedging that guarantees bacterial survival (Ryan-Payseur and Freitag, 2018). Studies suggest that ComX inhibits matrix production, especially the TasA protein, and promotes sporulation during the early stages of biofilm development in the wild type (Špacapan et al., 2020). This, on one hand, may enable bet-hedging behavior, but on the other, may burden cells with the metabolic costs of early sporulation initiation.
It has been further demonstrated that the tricarboxylic acid (TCA) cycle is rapidly upregulated during early biofilm development (12–16 h) driven by a decrease in the activity of CcpA and CodY, whereas the fatty acid synthesis pathway is downregulated characterized by reduced expression of plsX and plsC (Pisithkul et al., 2019). CcpA represses a gene that either decreases the rate of attachment of cells to a biofilm or increases the rate of detachment of cells from the biofilm (Stanley et al., 2003). CodY is also involved in repressing the transcription of kinB, hence spo0A (Piggot and Hilbert, 2004; Tojo et al., 2013; Eijlander et al., 2014).
As biofilms age, they disassemble because constituent cells release their amyloid fibers as a result of the incorporation of
Sporulation is the last resort response that Bacillus can deploy to survive under adverse conditions (Rodriguez Ayala et al., 2020). The capacity to form sporulating biofilms is variable among Bacillus species ranging from as low as 0.01% to 10% of the attached recoverable population (Ren et al., 2004; Ryu and Beuchat, 2005; Wahlen et al., 2018) to more prolific strains producing 50–90% spores within biofilms (Wijman et al., 2007; Faille et al., 2014).
Spo0A∼P initially accumulates to intermediate levels in matrix producers leading eventually to the expression of genes involved in sporulation (She et al., 2020). Spo0A∼P, the positive regulator, works by activating the transcription of several key sporulation-specific genes, particularly the spoIIE, spoIIA, and spoIIG genes as well as successive actions of specific sigma factors σ F, σ E, σ G, σ K, and σ A. After asymmetric division, σ F is activated in the forespore and σ E in the mother cell. This leads to engulfment of the forespore when σ G becomes active in the engulfed spore and σ K in the mother cell with engulfed endospore, finally to spore maturation and release (Doi, 1989; Stragier and Losick, 1996; Fujita et al., 2005; Madigan et al., 2014).
Once the nutrient conditions resume, the spore germinates by releasing Ca-dipicolinic acid (Ca2+-DPA) in the presence of GR—GerA, GerB, GerK, and GerD, finally activating σ
A in the germinated cell and resuming vegetative cycle. Germination is triggered upon exposure to an array of nutrients or conditions activating the GR in the inner spore membrane. In B. subtilis, GerA receptor induces germination in response to
SpoVA proteins form a channel in the spore inner membrane for release of DPA and CLEs—CwlJ and SleB, degrade the peptidoglycan cortex layer leading to complete rehydration of the spore core, after which the enzyme activity within the spore protoplast resumes (Moir, 2006; Setlow, 2014b). This allows RpmE and Tig to resume protein synthesis, TCA cycle, and essential cellular activities, thus cell size starts increasing (Sinai et al., 2015).
Conclusions
Although LMF are traditionally considered safe, the concerns raised by the recent outbreaks and recalls connected to LMF cannot be overlooked. The natural tendency of Bacillus species to adapt to various environmental stresses by altering their gene expression and physiology remains a significant challenge for the food industry. The processes of sporulation and biofilm development are intertwined rather than linear independent pathways initiating with matrix production in the early stages, followed by transition to sporulation under conditions of extreme stress.
The conditions in the food processing plant can act as a switch between the two interrelated genetic pathways and also likely favor the survival, selection, and development of extremely resistant strains with high enzymatic activity, tolerance to a wide range of pH and temperature. It remains to be elucidated whether or not biofilm formation by Bacillus species could be associated with the production of toxins in mono- and/or multispecies forms. Therefore, detailed studies closer to the persistence niches of Bacillus species in the food chain are imperative to understand the possible synergy between sporulation, germination, biofilm formation, and toxin production at a molecular level.
Footnotes
Authors' Contributions
S.L. wrote the review with inputs from D.M.M. S.F. and A.G.M.S. supervised and edited the article. All authors read and approved the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This project (NOSPORES-DFI) was funded by grant-aid under the Food Institutional Research Measure project no. 14/F/845, administered by the Department of Agriculture, Food and the Marine (DAFM), Ireland.