
Abstract
An increasing number of outbreaks are caused by foodborne pathogens such as Escherichia coli and Salmonella, which often harbor antimicrobial resistance (AMR) genes. We previously demonstrated the transmission of pathogens from animal operations to produce fields on sustainable farms, which illustrated an urgent need to develop and implement novel prevention methods and remediation practices such as the vegetative buffer zone (VBZ) to prevent this movement. The focus of this study was to use whole-genome sequencing (WGS) to characterize the AMR, virulence, and single-nucleotide polymorphism profile of 15 Salmonella and 128 E. coli isolates collected from small-scale dairy and poultry farms on a research station in North Carolina. Phenotypically, seven E. coli and three Salmonella isolates displayed resistance to antibiotics such as tetracycline (n = 4), ampicillin (n = 4), nalidixic acid (n = 3), chloramphenicol (n = 2), sulfisoxazole (n = 1), and streptomycin (n = 1). A single E. coli isolate was found to be resistant to five different antibiotic class types and possessed the blaTEM-150 resistance gene. Virulence genes that facilitate toxin production and cell invasion were identified. Mauve analysis of the E. coli isolates identified seven clusters (dairy-six and poultry-one) indicating that transmission is occurring from animal operations to fresh produce fields and the surrounding environment when the VBZ is denudated. This suggests that the VBZ is a useful barrier to reducing the transmission of enteric pathogens in agricultural systems. Our study demonstrates the prevalence of AMR and virulence genes on small-scale sustainable farms and highlights the advantage of using WGS to assess the impact of the VBZ to reduce the transmission of E. coli and Salmonella.
Introduction
Enterohemorrhagic Escherichia coli (EHEC) and Salmonella are major foodborne pathogens in beef, dairy, and poultry products. The Centers for Disease Control and Prevention (CDC) estimates that >2.8 million people in the United States suffer from antibiotic-resistant infections annually. One antimicrobial resistant nontyphoidal Salmonella has been linked to 212,500 infections and 70 deaths in the United States annually (CDC, 2019). Similarly, extended-spectrum beta-lactamase (ESBL) producing Enterobacteriaceae are responsible for 197,400 hospitalizations and 9100 deaths in 2017 alone (CDC, 2019). Often, the Salmonella and EHEC isolates that cause foodborne illnesses in humans also carry pathogenicity islands and virulence genes that aid their survival and replication inside of the gastrointestinal tract (Desvaux et al, 2020).
There has been a 17% increase in certified organic farms since 2016, which incorporate both organic (avoiding the use of synthetic fertilizers or pesticides) and sustainable practices (i.e., planting cover crops and integrating livestock and crop production) (United States Department of Agriculture [USDA], 2019). However, it is known that livestock such as dairy and poultry can shed AMR pathogens such as EHEC and Salmonella through feces and urine, which can then be mixed into soil amendments or spread into the nearby surface and irrigation water (Iwu et al, 2020; Glaize et al, 2020). Whole-genome sequencing (WGS) has been used to monitor the transmission dynamics of AMR pathogens more effectively in humans, retail meat, and food animals (McDermott et al, 2016). WGS allows outbreak investigators the ability to quickly compare different bacteria from clinical samples to possible food sources during an active foodborne outbreak and detect coinciding outbreaks in real time (Sekse et al, 2017).
Despite these advances in AMR surveillance, it has become apparent that new ways to prevent the transmission of AMR pathogens within integrated livestock and produce agricultural environments are needed. Previously we established a vegetative buffer zone (VBZ) and demonstrated its impact in reducing the transmission of enteric pathogens between animal operations and fresh produce fields (Glaize et al, 2021). The purpose of this study was to use WGS to (1) characterize the AMR, virulence, and single nucleotide polymorphism (SNP) profiles of 128 E. coli and 15 Salmonella isolates on two small-scale dairy and poultry farms in North Carolina, (2) determine the source of AMR E. coli and Salmonella in these systems, and (3) ascertain the impact of distance on the transmission of AMR pathogens from animal operations to fresh produce fields.
Materials and Methods
Experimental design
An in-depth explanation of the experimental design of this study can be found in Glaize et al (2021). In brief, two North Carolina small-scale sustainable poultry and dairy farms located at a research station were sampled for 2 years from 2018 to 2019. These farms claim to not use antimicrobials to treat the livestock. A VBZ was grown between the animal housing units and the produce fields to mitigate the transmission of E. coli and Salmonella (Fig. 1 in Glaize et al, 2021).

SNP trees of Salmonella isolates from dairy and poultry farms. Each tip label includes information about the isolate ID, farm, source type, distance from the VBZ, serovar, and MLST. The “Source” column is color coded by source type: air (white), barrier (gray), soil (light gray), produce (dark gray), and feces (black).
The E. coli and Salmonella isolates discussed in this article were recovered from three sample types collected from five sources: fecal matter, fresh produce (tomatoes or romaine lettuce), and environmental (i.e., soil, barrier, and air). Each farm had three air sample stations and fresh produce fields located 10, 61, and 122 m away from the VBZ and downwind of the animal housing units (Fig. 1 in Glaize et al, 2021). A separate air sample station was located at the horticulture plots of the farm upwind from both the dairy and poultry units and served as our air control.
Phenotypic characterization of AMR profile
The phenotypic AMR profile of E. coli (n = 128) and Salmonella (n = 15) isolates was determined using the broth microdilution Gram-negative National Antimicrobial Resistance Monitoring System (NARMS) CMV3AGNF plate (Trek Diagnostic Systems, Cleveland, OH) as described by Glaize et al (2020). The isolates were exposed to a panel of 14 preselected drugs as follows: cefoxitin (FOX; 0.5–32 μg/mL), azithromycin (AZI; 0.12–16 μg/mL), chloramphenicol (CHL; 2–32 μg/mL), tetracycline (TET; 4–32 μg/mL), ceftriaxone (AXO; 0.25–64 μg/mL), amoxicillin/clavulanic acid (AUG2; 1/0.5–32/16 μg/mL), ciprofloxacin (CIP; 0.015–4 μg/mL), gentamicin (GEN; 0.25–16 μg/mL), nalidixic acid (NAL; 0.5–32 μg/mL), ceftiofur (XNL; 0.12–8 μg/mL), sulfisoxazole (FIS; 16–256 μg/mL), trimethoprim/sulfamethoxazole (SXT; 0.12/2.38–4/76 μg/mL), ampicillin (AMP; 1–32 μg/mL), and streptomycin (STR; 32–64 μg/mL).
The NARMS breakpoints were used to determine the tested isolates' susceptibility and resistance status (NARMS, 2019). Isolates resistant to three or more classes of antimicrobials are classified as multidrug resistant (MDR).
DNA isolation and WGS
An in-depth explanation of the DNA isolation and WGS protocols is available in Glaize et al (2020; 2021). In brief, the DNA libraries were prepared using the Nextera XT kit (Illumina, San Diego, CA). These libraries were then sequenced on an Illumina MiSeq (Illumina) using 250 or 300 bp paired-end reads (MiSeq reagent kit, version 3). Sequenced isolates for E. coli (Accession: PRJNA293225) and Salmonella (Accession: PRJNA293224) were submitted to the National Center for Biotechnology Information GenomeTrakr database (Supplementary File S1).
Bioinformatic analysis
The bioinformatic pipeline used to analyze the E. coli and Salmonella isolates is described by Glaize et al (2020 and 2021) and Hull et al (2021). In summary, de novo genome assembly was completed using Shovill version 1.1.0 and SPAdes version 3.14.1 (Seemann, 2020c; Prjibelski et al, 2020). The QUAST program, version 5.0.2, was used to check the sequence quality of each isolate (Mikheenko et al, 2018). The program ABRicate version 1.0.0 was then used to screen for AMR, ESBL, and virulence genes using the CARD, MEGARes, VFDB, and Ecoli_VF databases (
The multilocus sequencing typing (MLST) was determined using the command-line tool MLST version 2.19.0 (Jolley and Maiden, 2010; Seemann, 2020b). E. coli pathotype categories were determined using the recommendations outlined by Robins-Browne et al (2016). E. coli phylogroups (A, B1, B2, C, D, E, F, and G) for the E. coli isolates were determined using the web-based tool ClermonTyping version 1.4.0 (Beghain et al, 2018; Clermont et al, 2019). ParSNP version 1.5.6 from the Harvest suite was used to create the SNP core-genome phylogeny tree (Treangen et al, 2014). Dendroscope version 3.7.4 was used to visualize and edit the phylogenetic tree (Huson et al, 2012; Huson et al, 2007). The multiple alignment tool Mauve, version 2.4.0, was used to assess the homology of genomic sequences (Darling et al, 2004). Annotations were done using the phytools package version 0.7–80 with R version 4.04 and RStudio version 1.4.1106 (R Core Team, 2021; Revell, 2012).
Results and Discussion
Phenotypic and genotypic AMR profile of Salmonella and E. coli
A total of 15 Salmonella and 128 E. coli isolates were recovered from 57 environmental samples collected from the dairy (Salmonella = 9; E. coli = 111) and poultry (Salmonella = 6; E. coli = 15) farms as well as the control air station (Salmonella = 0; E. coli = 2) (Table 1). Three Salmonella (20%; n = 3/15) and seven E. coli (5.5%, n = 7/128) isolates were found to be resistant to antibiotics (Table 1). The incidence of Salmonella resistance to nalidixic acid isolated from poultry farms is similar to what has been previously reported. For example, Velasquez et al (2018) found a 23% incidence of resistance to nalidixic acid in Salmonella isolates recovered from a poultry farm (Velasquez et al, 2018).
Number of Escherichia coli and Salmonella Isolates by Distance, Source, and Farm Type
Bold indicate the total number of isolates recovered from a source at different distances.
A total of 143 Escherichia coli (n = 128) or Salmonella (n = 15) isolates were recovered from 57 fecal, produce, and environmental samples.
VBZ, vegetative buffer zone.
In total, six different AMR profiles were observed: NAL (n = 3), TET (n = 2), Amp (n = 2), TET AMP (n = 1), CHL (n = 1), and CHL TET FIS AMP STR (n = 1). The frequency of resistance was highest in TET (3.6%; n = 4/111), AMP (3.6%; n = 4/111), and NAL (50%; n = 3/6) (Table 2). One E. coli isolate recovered from a VBZ sample from the dairy farm was found to be MDR and with resistance to chloramphenicol, tetracycline, sulfisoxazole, ampicillin, and streptomycin. This outcome is aligned with a report from Innes et al (2021), which found a lower prevalence of MDR contamination of retail meats from organic processing plants than the conventional ones in the United States.
Escherichia coli and Salmonella Antimicrobial Resistance in Dairy and Poultry Farms
The two E. coli isolates from the control air station is not included in this table.
Antimicrobials: FOX, cefoxitin; AZI, azithromycin; CHL, chloramphenicol; TET, tetracycline; AXO, ceftriaxone; AUG2, amoxicillin/clavulanic acid; CIP, ciprofloxacin; GEN, gentamicin; NAL, nalidixic acid; XNL, ceftiofur; FIS, sulfisoxazole; SXT, trimethoprim/sulfamethoxazole; AMP, ampicillin; STR, streptomycin. Resistance breakpoints: FOX, ≥32 μg/mL; AZI, ≥16 μg/mL; CHL, ≥32 μg/mL; TET, ≥32 μg/mL; AXO, ≥64 μg/mL; AUG2, ≥32/16 μg/mL; CIP, ≥4 μg/mL; GEN, ≥16 μg/mL; NAL, ≥32 μg/mL; XNL, ≥8 μg/mL; FIS, ≥256 μg/mL; SXT, ≥4/76 μg/mL; AMP, ≥32 μg/mL; and STR, ≥64 μg/mL. MICs are expressed as μg/mL.
MICs, minimum inhibitory concentrations.
All of the AMR E. coli were recovered from fecal (n = 2) and environmental (i.e., barrier [n = 2] and air [n = 3]) samples collected from the dairy farm between April and May of 2019 (Table 1). Salmonella Agona and Salmonella Give were retrieved from dairy fecal samples, while Salmonella Typhimurium and Salmonella Newport were recovered from poultry, produce, and soil samples (Fig. 1A, B). Based on WGS analysis, two genes that can confer resistance to aminoglycosides [aac(6′)-laa (n = 3) and acc(6′)-ly (n = 12)] and one to fosfomycin (fosA7 [n = 6]) were detected in the Salmonella isolates. Point mutation gyrA_D87G was detected in 100% of Salmonella Typhimurium isolates. Resistance genes to phenicol (floR), sulfisoxazole (sul2), and aminoglycosides [aph(3″)-Ib and aph(6)-Id] were detected in the MDR E. coli isolate (Fig. 2).

SNP tree of Escherichia coli isolates from a dairy farm. Each tip label includes information about the isolate ID, farm, source type, distance from the VBZ, O-antigen, H-antigen, MLST, and phylogroup. Isolates recovered from the dairy farm were classified into four phylogroups from top to bottom: A, B1, C, and D. The “Source” column is color coded by source type: air (white), barrier (gray), soil (light gray), produce (dark gray), and feces (black). Isolates that are either phenotypically resistant (black) or susceptible (white) to ampicillin (AMP; 1), chloramphenicol (CHL; 2), sulfisoxazole (FIS; 3), streptomycin (STR; 4), or tetracycline (TET; 5) are shown in the third group of columns. The presence (gray) or absence (white) of the point mutations uhpT_E350 (1), rpoB_V146F (2), cyaA_S352T (3), and parE_I355T (4) is shown in the fourth group of columns. Finally, the presence (black) or absence (white) of antimicrobial resistant genes aph(6)-Id (1), aph(3″)-Ib (2), tet (3), fosA7 (4), floR (5), and sul2 (6) is indicated in the fifth group of columns. MLST, multilocus sequencing typing; SNP, single-nucleotide polymorphism; VBZ, vegetative buffer zone.
WGS analysis revealed that all E. coli and Salmonella isolates sequenced carried genes that conferred resistance to beta-lactamases. The gene marA was detected in all the 128 E. coli isolates, and the encoded MarA protein acts as a global regulator of the MDR efflux pump AcrAB and reduces the permeability to beta-lactams. A single E. coli isolate possessed the gene blaTEM-150 . This is the same isolate that was determined to be MDR by an antimicrobial susceptibility testing. A similar result was reported by Nüesch-Inderbinen et al (2015) who recovered 12 ESBL-producing Enterobacteriaceae from ready-to-eat produce such as tomatoes, spinach, and precut chives. All 15 Salmonella isolates carried the E. coli-specific ampH gene and the marA efflux pump regulator gene. A total of 12 Salmonella isolates possessed the ramA gene, which encodes the AcrAB efflux pump's regulator protein.
Virulence profile of Salmonella and E. coli
According to the Virulence Factor Database (VFDB), all Salmonella isolates possessed the virulence genes mig-14, mgtC, and spvB, which confer resistance to antimicrobial peptides, aid in magnesium transport, and produce an intracellular toxin, respectively. The genes rck (mediates cell invasion) and sodCI (a stress protein) were only found in Salmonella Typhimurium isolates from fresh produce samples from the poultry farm. Similarly, the three Salmonella Give isolates from dairy fecal samples were found to have the cdtB gene, which produces the cytolethal distending toxin (CDT). However, the cdtA and cdtC genes, which help the CDT bind to target cells, were not detected. Of the 128 E. coli isolates, 2 isolates were determined to be atypical enteropathogenic E. coli (aEPEC). This was due to the absence of bfp and the presence of eae, paa, ler, east1, espA, espB, espD, espF, espG, espH, and tir. These isolates were recovered from the dairy VBZ in April 2019.
Overall, both the antibiotic-free farms on the research station had a low prevalence of AMR and virulence genes present in the Salmonella and E. coli isolates. The genotypic profiles of these isolates suggest that they have the potential to confer to different medically important drug classes such as aminoglycosides, tetracyclines, and aminopenicillins. For example, a single E. coli isolate recovered from the dairy VBZ was MDR and possessed the bla TEM-150 resistance gene. Furthermore, most Salmonella isolates from both the dairy and poultry farms contained ampH, an E. coli-associated beta-lactamase gene, and PBP2, which encodes for a protein that can bind to beta-lactam antibiotics and render them ineffective. It should be stated that the genotypic profile of isolates does not always correlate to phenotypic resistance or susceptibility to antibiotics (Doyle et al, 2020).
However, there is an extensive amount of evidence that both Salmonella and E. coli can act as reservoirs of AMR and virulence genes and disseminate them into the environment (Iwu et al, 2020). Recent publications have suggested that there may be a correlation between the presence of antibiotic resistance genes and virulence factors within a bacterium (Cepas and Soto, 2020; Ewers et al, 2021; Higgins et al, 2020). This correlation can be positive, negative, or neutral depending on the species, antibiotics, and virulence factors assessed. Higgins et al (2020) found that the distribution of virulence genes between susceptible and AMR Salmonella isolates from clinical samples was not statistically significant (Higgins et al, 2020). However, the distribution frequency did become significant when comparing susceptible and MDR Salmonella.
As more high-quality WGS data become available, it will soon be possible to further illuminate the relationship and fitness cost of AMR genes and virulence factors in Salmonella and E. coli (Seske et al, 2017; Carlos et al, 2020, Iramiot et al, 2020).
Phylogroups and confirming the transmission of E. coli
A total of five phylogroups were identified in the E. coli isolates. The majority (93.75%; n = 120/128) of these isolates were characterized as phylogroups A (n = 34) and B1 (n = 86), while the remainder were sorted into phylogroups B2 (n = 1), C (n = 3), and D (n = 3). The two E. coli isolates recovered from the control air station belonged to phylogroup B1. All of the isolates retrieved from dairy and poultry fecal samples were a part of phylogroup B1 (n = 10). The isolates categorized in phylogroups B2, C, and D were from produce and soil samples.
The three phylogroup C isolates were recovered from two romaine lettuce samples located 61 m away from the dairy VBZ. Similarly, one phylogroup D isolate was retrieved from a romaine lettuce sample located 122 m away from the dairy VBZ. The other phylogroup D isolates (n = 2) were retrieved from two soil samples located 122 m away from the dairy VBZ. Finally, the one phylogroup B2 isolate was recovered from a soil sample located 10 m away from the poultry VBZ.
A total of 7 E. coli groups (dairy = 6; poultry = 1) were identified based on Mauve SNP analysis, comprising 31 E. coli isolates. Isolates sorted into these groups belonged to phylogroups A and B1 and were recovered from fresh produce, fecal, and environmental (i.e., soil and barrier) samples between April and May 2019. The assembled E. coli poultry SNP tree identified an isolate recovered from a poultry fecal sample that closely matches the SNP patterns of two E. coli isolates that were collected from soil samples located 10 m away from the VBZ (group 1; Fig. 3A). The Mauve analysis found that the genomes of these three isolates possessed similar core genome and SNP alignment (Fig. 3B). This group documents the transmission of E. coli between poultry feces and agricultural soil used for growing produce.
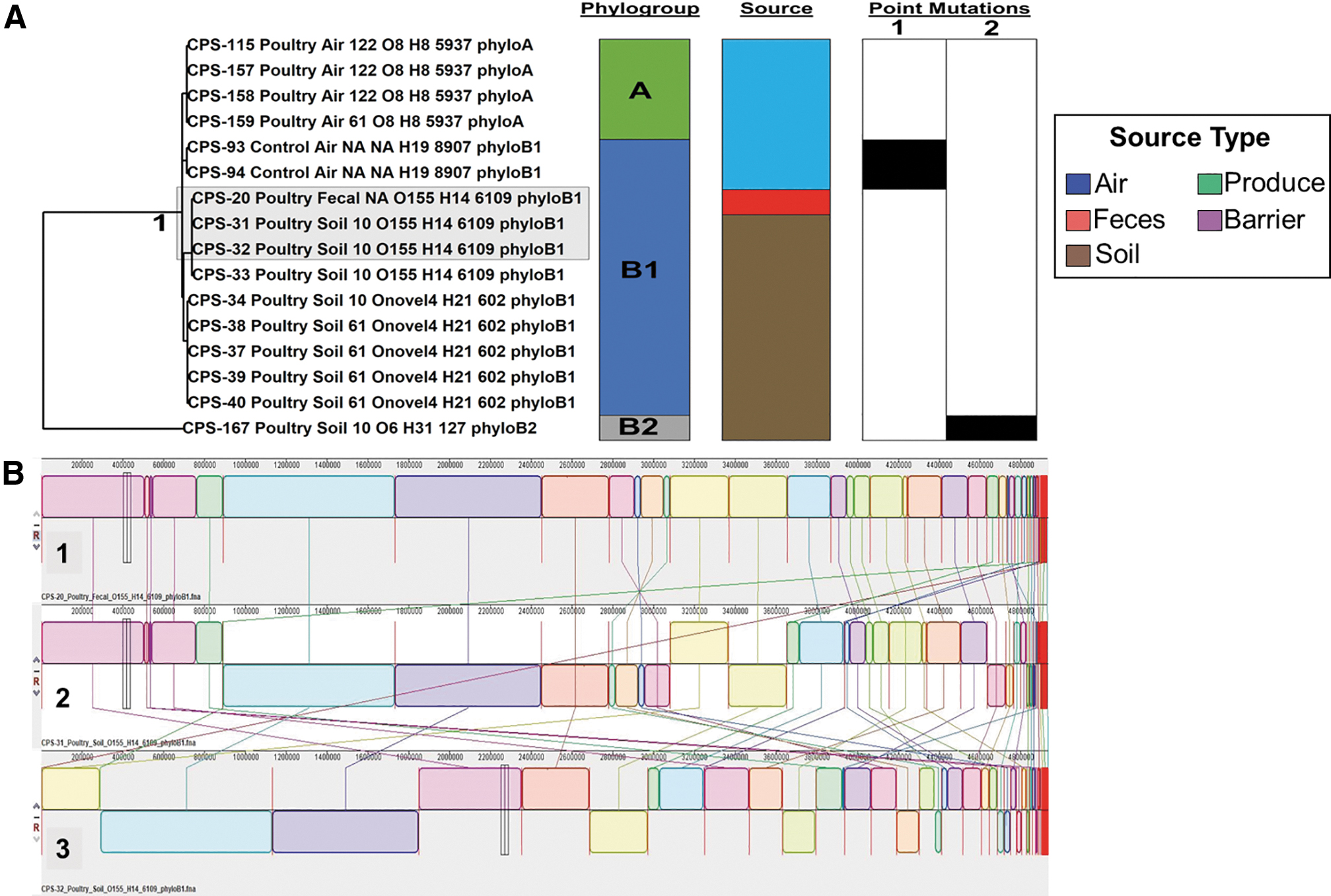
SNP tree and alignment of E. coli isolates from a poultry farm.
The same trend can be found in the dairy E. coli SNP tree (Fig. 2). In group 6, two E. coli isolates from two dairy fecal samples have the same serotype (O168:H8), MLST (718), and phylogroup (B1) as six isolates recovered from one barrier (n = 2) and two produce (n = 4) samples. They also share a similar core genome and SNP pattern (Fig. 4F). These two produce samples were collected 10 and 122 m away from the dairy VBZ. Similarly, an E. coli isolate from a barrier sample in group 2 had the same SNP and molecular characterization profile as isolates from a dairy fecal sample (n = 3) (Figs. 2 and 4B). This group supports the possibility that the VBZ can block E. coli transmission between dairy fecal samples and environmental air, soil, and fresh produce samples.
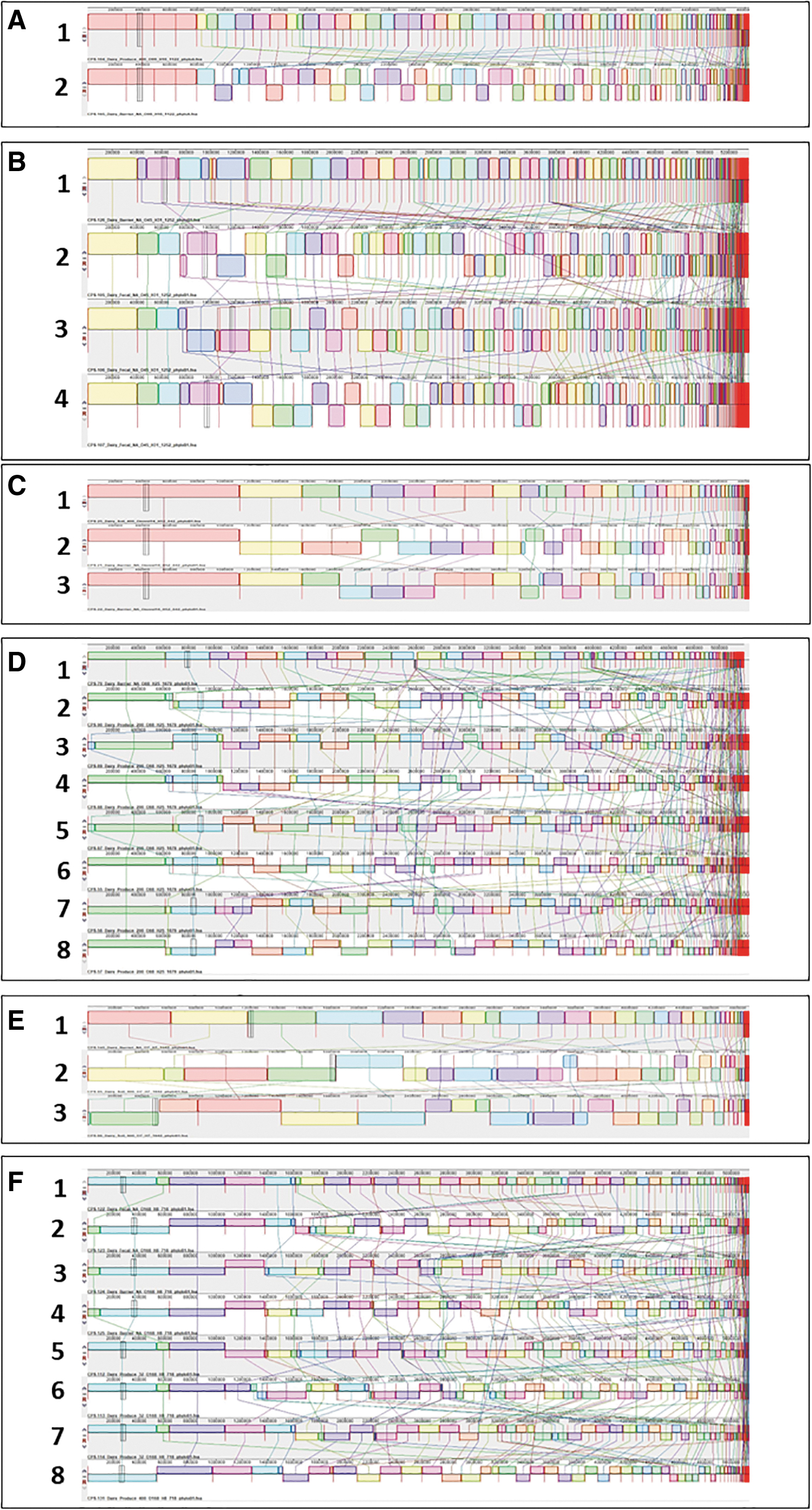
SNP alignment of E. coli isolates from a dairy farm.
It is also important to note that the Mauve analysis of the two control E. coli isolates did not match the core genome or SNP patterns of the E. coli isolates recovered from the poultry or dairy farms.
According to the dairy E. coli SNP tree, many barrier isolates are related to isolates retrieved from fresh produce (groups 1 and 4) or environmental soil (groups 3 and 5) samples (Fig. 2). The produce samples from these groups were grown at 61 and 122 m, and the positive soil samples were located 122 m from the dairy VBZ. It is important to note that the dairy VBZ did not provide side coverage from the wind or dairy pasture to the 61 and 122 m plots (Fig. 1 in Glaize et al, 2021). As a result, the produce was not fully protected by the VBZ and was exposed to contaminated unfiltered air. In fact, more produce, soil, and air samples situated away 61 and 122 m from the VBZ were positive for E. coli and Salmonella (Table 1).
Publications by Jeamsripong et al (2019) and Theofel et al (2020) have also reported a similar increase in bacterial contamination of fresh produce that is directly downwind and exposed to animal operations (Jeamsripong et al, 2019; Theofel et al, 2020). Taken together, these groups demonstrate that the VBZ can reduce the transmission of E. coli from dairy fecal and environmental air samples to fresh produce fields when adequately covered.
Finally, the majority of recovered E. coli and Salmonella isolates were retrieved between April and May of 2019. During this period, the back row of the VBZs, composed of poplar trees, were still in the process of leafing out. Once the poplar trees were close to full capacity a month later, the number of isolates recovered from the dairy and poultry farms dropped. It suggests that the VBZ cannot completely block the transmission of foodborne pathogens until the leaves have fully emerged. This also demonstrates how effective the VBZ is at reducing the transmission of E. coli and Salmonella.
Conclusion
Overall, the transmission of Salmonella and E. coli from dairy and poultry units to fresh produce fields is occurring. These pathogens are AMR and, at least one, MDR, despite the farms' claims of not using antimicrobials, and can be recovered from fresh produce, livestock fecal samples, and the environment. However, the VBZ, at its fullest capacity, is an effective way of reducing the transmission of these pathogens from animal operations to fresh produce within a distance of 125 m from animal activities. The information generated from the study can be used to assess the transmission dynamics of AMR foodborne pathogens on small-scale farms and evaluate prevention methods that can reduce the transmission of these pathogens on agricultural farms. This study highlights the need for more effective ways of decontaminating agricultural soil between production cycles to reduce the safety risk of AMR pathogens in our food network.
Footnotes
Authors' Contributions
S.T. and E.G.-R. conceived of the presented idea and obtained funding to support this project. A.G. and L.R. conducted the field analysis and prepped the confirmed isolates for WGS. A.G., D.H., and A.H.M.v.V. performed the bioinformatic analysis. A.G. and S.T. wrote the article. S.T. and E.G.-R. supervised the findings of this work. All authors discussed the results and contributed to the final article.
Disclaimer
The contents are solely the responsibility of the authors and do not necessarily represent the official views of North Carolina State University.
Ethics Approval/Exemption Statement
No ethics approval was required for our study since it did not involve people, medical records, human samples, or live animals.
Source of Support
Center for Produce Safety, Specialty Crop Block Grant Program (USDA).
Disclosure Statement
No competing financial interests exist.
Funding Information
This study on antimicrobial resistance in Salmonella and Escherichia coli on small-scale farms was supported by the North Carolina State University College of Veterinary Medicine. The whole-genome sequencing was completed by the FDA GenomeTrakr program-funded grant 1U18FD00678801.
Supplementary Material
Supplementary File S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.