
Abstract
Shigella sonnei is a species of Shigella, and the infection rate of S. sonnei is increasing year by year. Eugenol is an active ingredient in clove essential oil and is a generally recognized as safe (GRAS)-certified food ingredient. The mechanism of inhibition of S. sonnei by eugenol has been investigated in this study. The minimum inhibitory concentration of eugenol against both S. sonnei ATCC 25931 and S. sonnei CMCC 51592 was 0.5 mg/mL and minimum bactericidal concentration (MBC) for both strains was 0.8 mg/mL. The inhibition effect of eugenol against S. sonnei was due to increased levels of reactive oxygen species in cells, changed cell membrane permeability, and induced cell membrane dysfunction, for instance, cell membrane hyperpolarization and intracellular ATP concentration drops. The results of confocal laser scanning microscope and field emission scanning electron microscopy showed that eugenol leads to decreased cell membrane integrity, resulting in changed cell morphology. Moreover, eugenol inactivated S. sonnei in Luria-Bertani (LB) broth and lettuce juice. These results indicated that eugenol could inactivate S. sonnei and has the potential to control S. sonnei in the food industry.
Introduction
Shigella can be divided into four major species: Shigella flexneri, Shigella sonnei, Shigella dysenteriae, and Shigella boydii (Chisenga et al., 2021). The proportion of deaths due to diarrhea caused by S. sonnei is increasing year by year and it has become an urgent problem for developing countries (Shad and Shad, 2021). S. sonnei is a rod-shaped, facultatively anaerobic Gram-negative bacterium and commonly linked to fecal contamination from human sources (Ranjbar and Farahani, 2019).
It can contaminate almost any food, including salads, fruits and vegetables, meat, dairy products, and cooked foods (Soffer et al., 2017). There have been reports of foodborne illness outbreaks caused by eating lettuce contaminated with S. sonnei (Bennett et al., 2018). It is spread through drinking water or food contaminated with the feces of an infected person (Zarei et al., 2019). Every year, about 200,000 people are infected by S. sonnei, and children under 5 years of age are the main affected group (Shad and Shad, 2021).
Several studies have been carried out to control Shigella in food and food industry. Organic acids and ozone, alone or in combination, were found to have an antibacterial effect on Shigella spp. (Singla et al., 2011). Further more, ultraviolet-C radiation has been investigated for its antibacterial activity against S. sonnei and is extensively employed in the research (Rajkowski, 2007). However, there are few studies on the control of S. sonnei by natural antimicrobials.
The use of plant-derived active substances as antimicrobial agents of growth of foodborne pathogenic and spoilage bacteria has become a hot topic in food research (Tako et al., 2020). Eugenol (4-allyl-2-methoxyphenol) is a bioactive ingredient of plant origin, mainly derived from clove, cinnamon, and pepper (Joardar et al., 2020). Eugenol is considered nonmutagenic and noncarcinogenic and generally recognized as safe (GRAS) by the U.S. Food and Drug Administration (SY/FL, GRAS-184.1257).
Eugenol has a variety of biological activities such as antioxidant, anticancer, antiviral, and nerve cell protection (Jahangir et al., 2021). In addition, its bacteriostatic activity has been reported. The study by Yadav et al. (2015) showed that biofilm formation of Staphylococcus aureus was inhibited by eugenol at 0.43 mg/mL. Ashrafudoulla's study indicated that eugenol at 1.07 mg/mL showed an inhibitory effect on Vibrio parahaemolyticus in Luria-Bertani (LB) broth containing 2% (v/v) NaCl, and the biofilm formed by V. parahaemolyticus on the surface of crab shells was eradicated by eugenol at 6.40 mg/mL (Ashrafudoulla et al., 2020).
Devi et al. (2010) studied the antibacterial mechanism of eugenol on Salmonella typhi, but only focused on the effect of eugenol on cell membranes. Similarly, Niu et al. (2019) studied the antibacterial mechanism of eugenol on Escherichia coli and found that its mechanism of action was mainly to cause physical and morphological changes in the cell membrane and to directly bind to genomic DNA through the minor-groove mode. However, the antibacterial effect and mechanism of eugenol against S. sonnei have not been investigated.
The study aimed to determine the antibacterial activity and inhibition mechanism of eugenol against S. sonnei. In addition, the inactivation effect of eugenol on S. sonnei in contaminated lettuce juice was determined.
Materials and Methods
Reagents
Eugenol (high performance liquid chromatography [HPLC] ≥98%, CAS, 97-53-0) was purchased from Chengdu Desite Biotechnology Co., Ltd., (Chengdu, China). LB agar and broth were purchased from Land Bridge Technology Co., Ltd., (Beijing, China). Before each assay, eugenol was solubilized in dimethyl sulfoxide (DMSO), and the final concentration of DMSO in all sample solutions was 0.1% (v/v). DMSO at 0.1% had no significant effect on the growth of S. sonnei (data not shown). All other chemicals and reagents were of analytical grade.
Bacterial stains and growth conditions
S. sonnei ATCC 25931 was obtained from American Type Culture Collection (ATCC, Manassas, VA) and the strain CMCC 51592 was purchased from the National Center for Medical Culture Collection (CMCC, Beijing, China). S. sonnei ATCC 25931 was used for the study of antibacterial mechanisms. S. sonnei strains stored at −80°C were streaked onto LB agar at 37°C for 18 h. S. sonnei strains were inoculated into 30 mL of LB broth and cultivated at 130 rpm at 37°C for 14 h. Bacteria were washed with phosphate-buffered saline (PBS, pH 7.2). The suspension concentration was adjusted to ∼3 × 108 colony-forming units [CFUs]/mL.
Determination of minimum inhibitory concentration and minimum bactericidal concentration
The minimum inhibitory concentration (MIC) of eugenol against S. sonnei was determined by the broth microdilution method (CLSI, 2018). The bacterial suspension (3 × 108 CFUs/mL) was diluted to 5 × 105 CFUs/mL in LB broth. Suspension and eugenol solution were added in equal volumes (100 μL) to a 96-well microtiter plate to give final dilutions of 0 (control), 0.25, 0.40, 0.50, 0.80, 1.00, and 1.60 mg/mL eugenol. LB broth supplemented with 0.05% (v/v) DMSO was used as the black background control.
The LB broth with added DMSO, but not inoculated with bacteria, was set as a black background control. Then, the optical density (OD)600nm of the sample was measured. The sample was incubated in a 37°C incubator for 24 h, and OD600nm was measured. MIC is defined as the minimum concentration of eugenol, which is the lowest concentration that resulted in ΔOD600nm (final OD600nm value minus initial OD600nm value) of ≤0.05.
For minimum bactericidal concentration (MBC) determination, the bacterial suspension was coated on LB agar and cultured at 37°C for 48 h. The MBC was defined as the lowest antimicrobial concentration corresponding to a 3-log reduction of viable cells.
Growth curve assay
Growth curve determination was carried out as described by Kang et al. (2020). Equal volumes of eugenol solution were mixed with the S. sonnei suspension (2 × 106 CFUs/mL) to achieve final concentrations of 1000 (2MIC), 500 (MIC), 250 (1/2MIC), 125 (1/4MIC), 62.5 (1/8MIC), 31.3 (1/16MIC), 15.7 (1/32MIC), 7.8 (1/64MIC), and 0 (control) μg/mL. LB broth with 0.05% (v/v) DMSO was set as a black background control.
The samples were incubated in the Bioscreen C Automated Microbiology Growth Curve Analysis System (Labsystems, Helsinki, Finland) at 37°C, with OD600nm values measured at 1-h intervals for 24 h.
Inactivation effect of eugenol on S. sonnei in LB broth
S. sonnei suspension (5 × 106 CFUs/mL) was treated with eugenol at concentrations of 0 (control), 0.25, 0.50, and 0.80 mg/mL and incubated at 37°C. Samples were spread-plated after being taken at 0, 1, 3, 6, and 9 h for serial dilution in PBS (pH 7.2). The plates were incubated at 37°C for 24 h before counting S. sonnei colonies. Five 200-μL aliquots of undiluted samples also were spread-plated onto five LB plates to achieve a detection limit of 1 CFU/mL.
Inactivation effect of eugenol on S. sonnei in lettuce juice
The antibacterial effect of eugenol on S. sonnei in lettuce juice was tested by the assay method described by Rossana et al. (2019), with slight modifications. Lettuce was bought from the local market (Yangling, China) and homogenized into juice, and then the juice was irradiated (6 kGy) for 2 s to remove the initial bacteria. S. sonnei ATCC 25931 and S. sonnei CMCC 51592 cells were added into the lettuce juice (5 × 106 CFUs/mL). Eugenol was added with final concentrations of 0 (control), 0.50, 0.60, and 0.80 mg/mL to the cell suspension.
The samples were treated at 25°C for 0, 1, 3, 6, and 9 h. The bacteria were diluted by buffered peptone water (BPW), plated onto LB agar, and incubated at 37°C for 24 h. After incubation, the total bacterial count was recorded.
Effect of eugenol on endogenous reactive oxygen species production
The change of ROS in S. sonnei was quantified using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA; Beyotime Institute of Biotechnology, Shanghai, China) by Hyun et al. (2020), with slight modifications. Thirty milliliters of bacterial solution (1 × 107 CFUs/mL) was incubated with 150 μL of 5 μM DCFH-DA at 37°C for 10 min. The cells were centrifuged at 12,000 × g for 10 min after being treated with eugenol (control, 1/8MIC, 1/4MIC, and 1/2MIC) for 10 min.
The cells were washed twice with PBS and suspended by PBS (12,000 × g, 10 min, 4°C). Then, the fluorescence intensity was measured at emission and excitation wavelengths of 525 and 488 nm, respectively, using a multimode enzyme reader platform (Spark®; Tecan, Salzburg, Australia).
Effects of eugenol on extracellular malondialdehyde production
As described by Cao et al. (2021a), the change of malondialdehyde (MDA) in S. sonnei was quantified using the lipid peroxidation MDA assay kit with slight modifications. S. sonnei strains were treated for 10 min at 37°C with eugenol (control, 1/8MIC, 1/4MIC, and 1/2MIC). Then, cells were centrifuged at 8000 × g for 10 min at 4°C. The supernatant (300 μL) was mixed with the MDA working solution (12,000 μL) at a concentration of 0.67% (w/v), boiled at 100°C for 1 h, and then cooled to room temperature.
Determination of the MDA content was done by measuring OD at 450, 532, and 600 nm.
Observation based on confocal laser scanning microscopy
As described by Yang et al. (2021), the influence of eugenol on cell membrane integrity was verified using a confocal laser scanning microscopy (CLSM), with some modifications. S. sonnei strains were washed twice in 0.85% (w/v) NaCl solution and the concentration was adjusted to 3 × 108 CFUs/mL. Eugenol (control, 1/2MIC, MIC, and 2MIC) was added to the bacterial solution before incubating at 37°C for 10 min.
After culture, bacteria were centrifuged at 10,000 × g at 4°C for 5 min and resuspended with 0.85% NaCl (w/v) solution. SYTO 9 and PI dyes were mixed and the dyes were added into the bacterial suspension in the dark for 5 min. Then, a confocal laser scanning microscope (CLSM; A1; Nikon, Tokyo, Japan) was used at 500 × magnification.
Field emission scanning electron microscopy-based observation
Field emission scanning electron microscopy (FESEM) was used to observe the morphological changes, as described by Shi et al. (2016) with some modifications. S. sonnei strains were washed twice in PBS and the concentration was adjusted to 3 × 108 CFUs/mL. Eugenol (control, MIC, 2MIC, and 4MIC) was added into all samples and incubated at 37°C for 2 or 4 h. Cells were initially immobilized with 2.5% (v/v) glutaraldehyde and stored overnight at 4°C.
The cells were then fixed again with glutaraldehyde for 5 h at 4°C. Then, cells were eluted with 30%, 50%, 70%, 90%, and 100% alcohol gradient for 10 min. Finally, samples were dried and fixed on supports. After vacuum spraying, the cells were observed using a field emission scanning electron microscope (S-4800; Hitachi, Tokyo, Japan).
Measurement of intracellular ATP concentrations
Intracellular ATP concentrations were measured as described by Gong et al. (2021), with slight modifications. S. sonnei suspension (3 × 108 CFUs/mL) was mixed with eugenol (control, MIC, and 2MIC). The samples were placed at 37°C for 10 min, and individual samples were subjected to plate counting. Then, samples were placed in an ice bath to maintain the stability of ATP. After lysing, the samples were immediately subjected to 100°C for 2 min to remove ATPase.
The supernatant was centrifuged at 8000 × g for 5 min at 4°C and kept in an ice bath. Determination was carried out using an ATP assay kit, according to the manufacturer's instructions (Beyotime Institute of Bioengineering). The supernatant and the prepared ATP standard solution were mixed with 100 μL of working suspension and then added into white, 96-well microtiter plates. Luminescence of each well was measured using a multimode microplate reader (Spark; Tecan).
Measurement of membrane potential
The influence of eugenol on S. sonnei membrane potential was previously described by Yang et al. (2019). S. sonnei strains were diluted in PBS (3 × 108 CFUs/mL) and incubated in a black 96-well plate at 37°C and cultured 30 min. One micromolar bis-(1,3-dibutyl barbituric acid) trimethine oxonol [DiBAC4(3); Molecular Probes, Eugene, OR] was added to the bacterial suspension at 37°C in the dark for 30 min.
Eugenol was added to each well, and the final concentration of eugenol was 0, MIC, or 2MIC. The fluorescence intensity was measured in the multimode, microplate reader platform (Spark; Tecan) at emission and excitation wavelengths of 515 and 492 nm, respectively.
Statistical analysis
All experiments were replicated three different times, each time the analysis was performed in duplicates. Data were analyzed using one-way analysis of variance. IBM SPSS statistical software was used to determine the significance of differences in means. Significance is indicated as follows: *p < 0.05 and **p < 0.01.
Results
MIC and MBC
The MIC of eugenol was found to be 0.50 mg/mL for S. sonnei ATCC 25931 and CMCC 51592 in this study. Bacteria treated with eugenol at 0.5 mg/mL could grow on LB agar, and the bacterial amount decreased by more than 3 log after 0.8 mg/mL eugenol treatment. Therefore, the MBC of eugenol was found to be 0.80 mg/mL for S. sonnei ATCC 25931 and CMCC 51592.
Growth curve
The effect of eugenol on the growth of S. sonnei is shown in Figure 1. It can be seen that eugenol at 500 and 1000 μg/mL completely inhibited the growth of bacteria. After 4 h of treatment of bacteria with 1/64MIC–1/2MIC concentration of eugenol, the growth rate was reduced and the OD600nm value was different compared with the control. Compared with the control, the OD600nm of bacteria decreased in a concentration-dependent manner after treatment with different concentrations of eugenol.
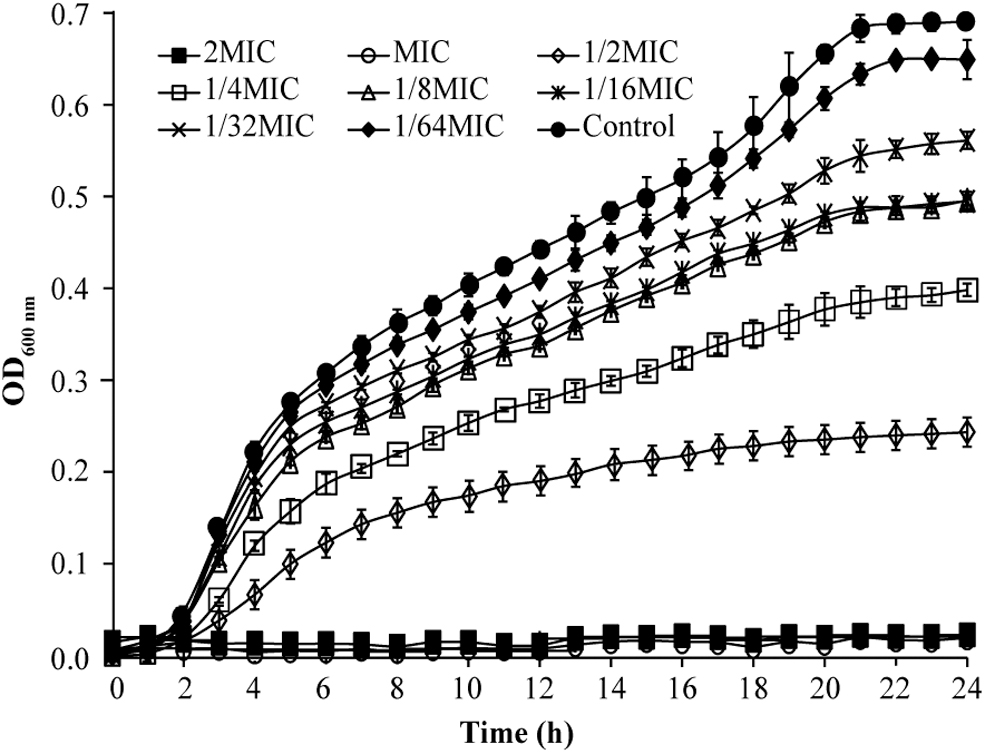
Effects of eugenol on the growth curve of Shigella sonnei ATCC 25931.
Inactivation effect of eugenol on S. sonnei in LB broth
As illustrated in Figure 2A, the starting bacterial load for each group was 6.9 log CFUs/mL. The bacterial number of the control increased by 1.4 log CFUs/mL after 9 h. The bacterial counts of S. sonnei exposed to 0.25 and 0.50 mg/mL of eugenol for 9 h were reduced by 1.0 and 1.9 log CFUs/mL, respectively. Cell counts of cultures exposed to eugenol at 0.80 mg/mL were below detectable levels (1 CFU/mL) at 0.5 h.

Inactivation of Shigella sonnei by eugenol in LB broth
Inactivation effect of eugenol on S. sonnei in lettuce juice
As shown in Figure 2B, the initial population of S. sonnei in the lettuce juice was about 6.7 log CFUs/mL. The number of cells in the control was retained at the original level for 9 h. Cell counts of S. sonnei exposed to eugenol at 0.50 mg/mL for 9 h decreased by 0.80 log CFUs/mL. After treatment with eugenol 0.60 and 0.80 mg/mL, the number of total viable cell counts decreased below detectable levels (1 CFU/mL) at 6 and 0.5 h.
Effects of eugenol on the intracellular ROS level in S. sonnei
The effect of eugenol on the intracellular ROS level in S. sonnei is shown in Figure 3. It can be seen that the ROS level significantly (p < 0.01) increased with the increase in eugenol concentration. ROS levels in cells treated with eugenol at 1/8MIC, 1/4MIC, and 1/2MIC were 5-, 8-, and 11-fold higher, respectively, compared with untreated (control) cells.
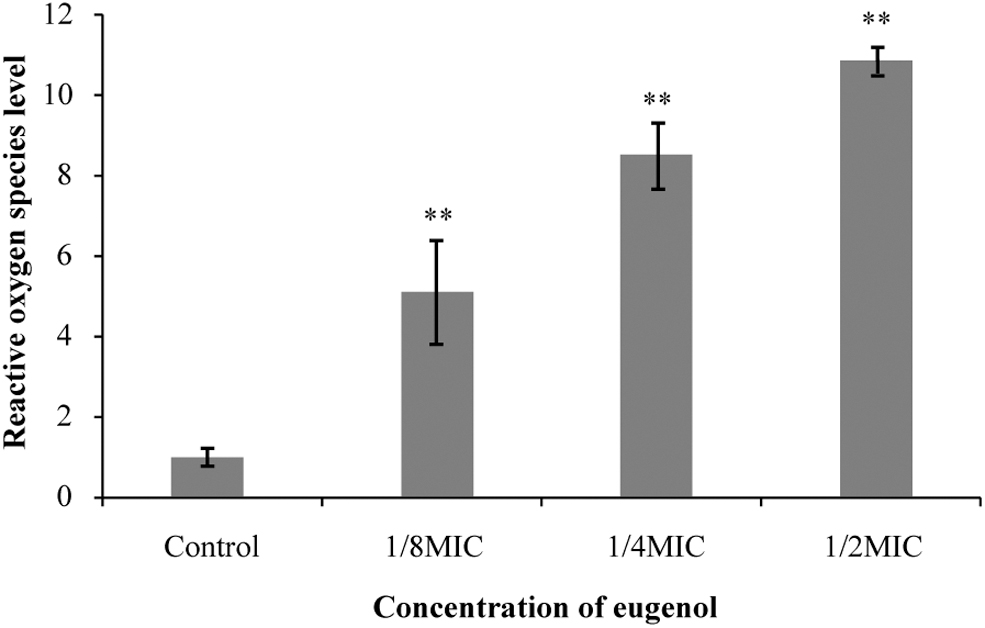
Effects of eugenol on intracellular reactive oxygen species levels of Shigella sonnei. **p < 0.01 versus the control.
Effects of eugenol on the extracellular MDA content in S. sonnei
The effect of eugenol on the extracellular MDA content of S. sonnei is shown in Figure 4. The bacteria that were not treated with eugenol had almost no MDA production. There was no significant difference in the extracellular MDA content of bacteria treated with 1/8MIC of eugenol compared with the control. The MDA content, respectively, increased to 0.06 ± 0.02 and 0.17 ± 0.05 mg/mL in cells treated with eugenol at 1/4MIC and 1/2MIC. With increase of eugenol concentration, the extracellular MDA content of S. sonnei significantly increased.
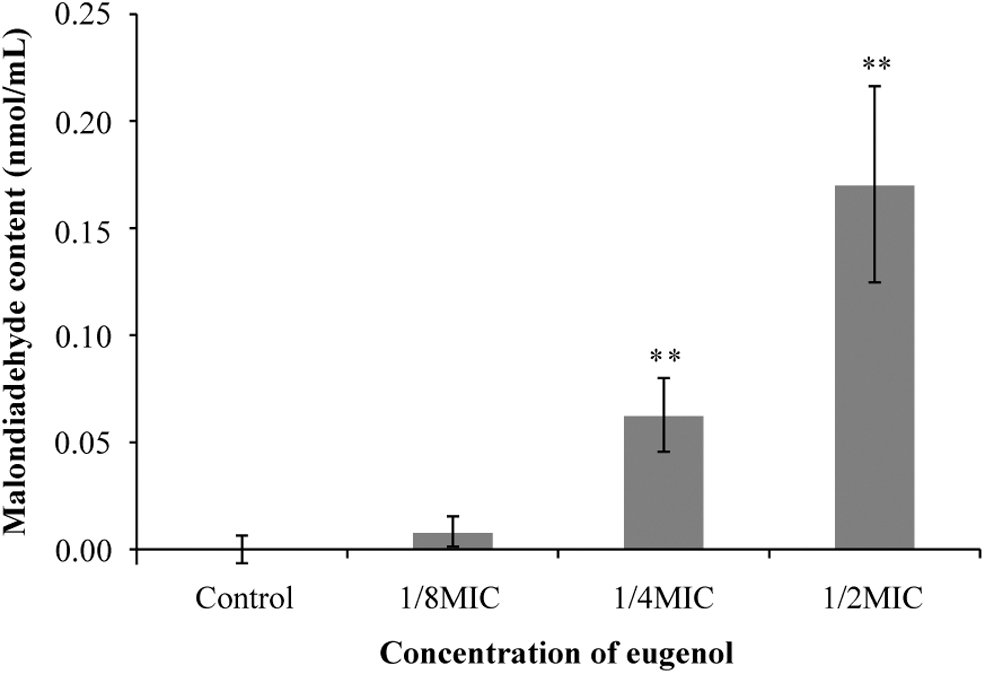
Effect of eugenol on the extracellular malondialdehyde content of Shigella sonnei. **p < 0.01 versus the control.
Effects of eugenol on the cell membrane in S. sonnei
The effect of eugenol on cell membrane integrity of S. sonnei is shown in Figure 5. Untreated control cells fluoresced green (Fig. 5A), which indicated that the cell membranes of untreated cells were intact. The cells treated with MIC of eugenol showed some red fluorescence (Fig. 5B). After treatment with 2MIC of eugenol, most cells fluoresced orange, indicating that eugenol reduced the integrity of the cell membrane (Fig. 5C).
CLSM-based observation of changes in the membrane integrity of untreated Shigella sonnei cells
All cells treated with eugenol at 4MIC showed complete red fluorescence, indicating that cell membrane integrity was disrupted (Fig. 5D). The results signified that eugenol exerts the membrane damage effect in a concentration-dependent manner.
Effects of eugenol on cell morphology in S. sonnei
FESEM analysis showed changes in cell morphology when S. sonnei cells were exposed to eugenol (Fig. 6). Untreated S. sonnei cells had not been altered in morphology and had a smooth surface (Fig. 6A, E). Cells were wrinkled after treatment with eugenol at MIC, with a rough surface and obvious depressions. Cells treated with eugenol at 2MIC showed substantial surface collapse on the cell membrane (Fig. 6C, G). When exposed to eugenol at 4MIC, cells displayed large surface collapse and abnormal cell debris (Fig. 6D, H).

FESEM-based observations (10,000 × magnification) of untreated Shigella sonnei at 2 h
Effects of eugenol on intracellular ATP concentrations in S. sonnei
The results showed a linear relationship between relative luminescence units and ATP concentration (R 2 = 0.9994, the standard curve is not shown). The concentration of ATP was 1.08 ± 0.08 μmol/1012 CFUs in the control. The intracellular ATP concentrations of S. sonnei treated by eugenol at MIC and 2MIC decreased by 0.76 and 0.83 μmol/1012 CFUs after 10 min, respectively.
Compared with the control, the intracellular ATP concentration of eugenol-treated S. sonnei decreased significantly (p < 0.01), and the decrease in ATP concentration showed a dependence on eugenol concentration (Fig. 7).
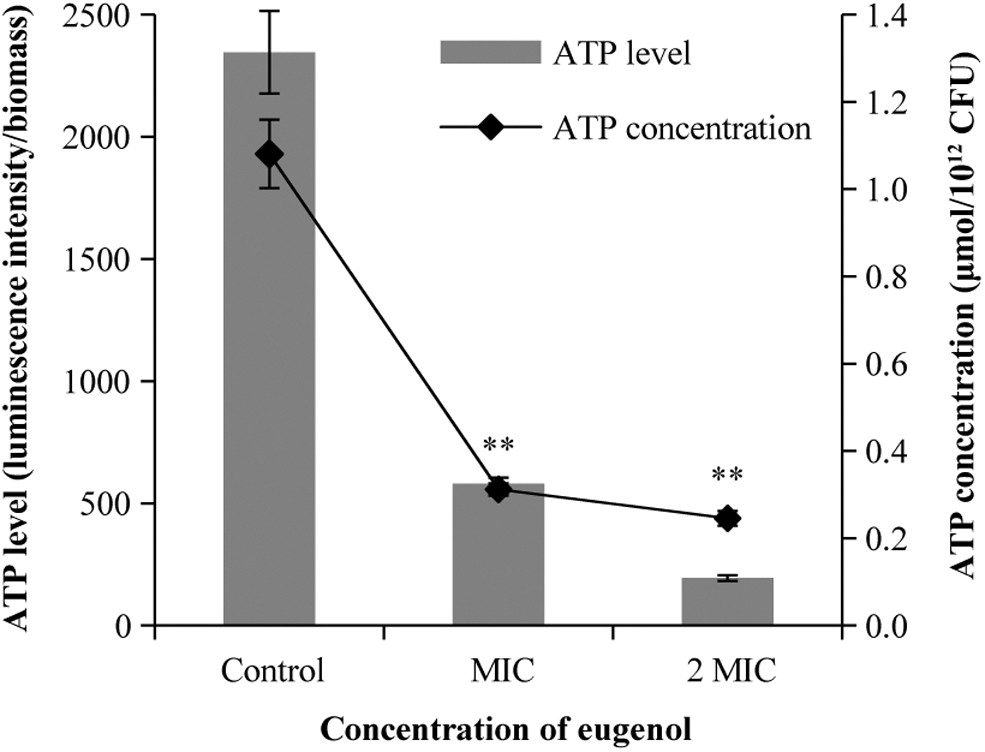
Effects of eugenol on intracellular ATP production in Shigella sonnei. **p < 0.01 versus the control.
Effects of eugenol on membrane potential in S. sonnei
As shown in Figure 8, the cell membranes of S. sonnei treated with eugenol showed hyperpolarization (negative fluorescence value), and this hyperpolarization became more obvious as the concentration of eugenol and incubation time increased.
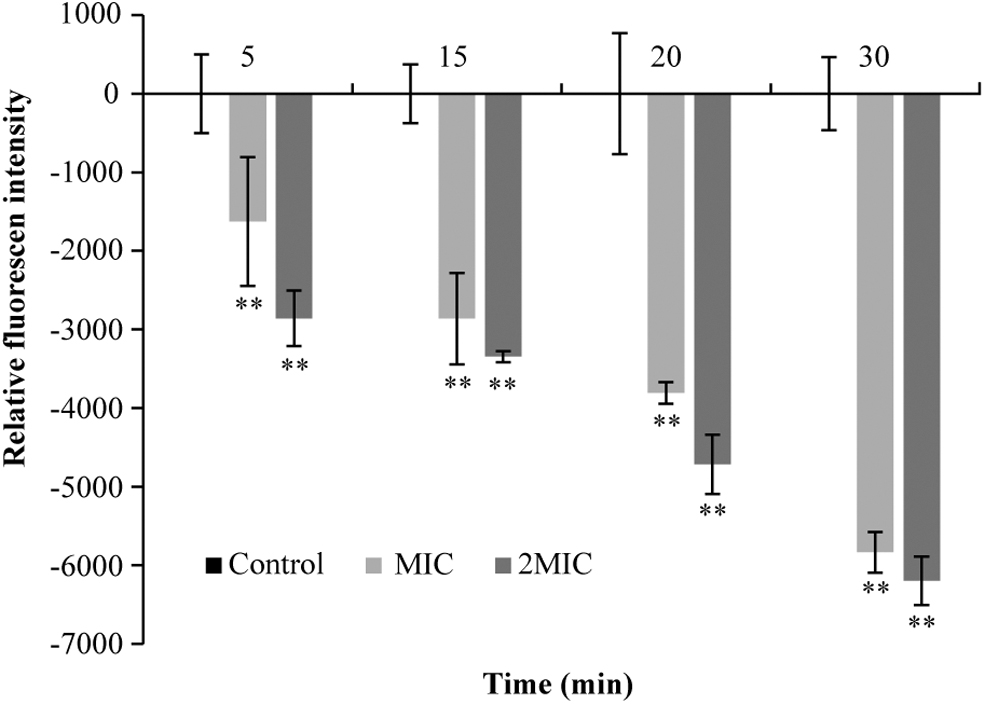
Effects of eugenol on the membrane potential. **p < 0.01 versus the control.
Discussion
In this study, the results showed that the MIC of eugenol against S. sonnei was 0.50 mg/mL. The inhibitory effect of natural active substances on S. sonnei was rarely mentioned in previous studies. Bagamboula et al. (2004) reported that the MIC of basil essential oil against S. sonnei was 1.00 mg/mL, while that of oregano essential oil 0.5% (w/v) was 5.00 mg/mL. Ashrafudoulla et al. (2020) reported that the MIC of eugenol against V. parahaemolyticus was 1.00 mg/mL. The MIC of eugenol against Streptococcus pneumoniae was 0.6 mg/mL (Hu et al., 2018).
The inactivation assays of eugenol on S. sonnei in LB broth indicated that eugenol had an antibacterial effect (Fig. 2A). Fan et al. (2021) showed that after treatment with thymoquinone at 0.80 mg/mL, the initial concentration of S. flexneri cells dropped from 8.6 log CFUs/mL to about 1.0 log CFU/mL within 24 h. In addition, our results also showed that S. sonnei was reduced to below the limit of detection (1 CFU/mL) with eugenol at 0.60 mg/mL within 6 h (Fig. 2B).
Although eugenol had an antibacterial effect on S. sonnei in both LB broth and lettuce juice, the effect was different in the two media. The difference in the antibacterial effect of eugenol in different media may be due to a reaction of chemicals that reduce the antibacterial effect of eugenol.
ROS levels of S. sonnei increased significantly with increase in eugenol concentration (p < 0.01) (Fig. 3) in this study. A study showed that Aspergillus niger was treated with 50 μg/mL cinnamaldehyde for 30 min and the level of intracellular ROS increased significantly (Qi et al., 2020). ROS production can increase oxidative stress and irreversible destruction of biomolecules and cause cell death (Ma et al., 2022).
The MDA content, a proxy for the degree of lipid oxidative damage, significantly increased after S. sonnei treatment with eugenol (Fig. 4). Similarly, Chen et al. (2021) found that Lindera glauca fruit essential oil extract at a concentration of 0.156 mg/mL can increase the intracellular MDA content of S. flexneri. Studies have shown that eugenol increased the intracellular ROS level and extracellular MDA content in S. sonnei. MDA accumulates in large quantities, leading to cell death (Lee and Lee, 2017).
After eugenol treatment, a decrease in the cell membrane integrity of S. sonnei was observed using CLSM, with a concentration-dependent effect on eugenol (Fig. 5). Tian et al. (2021a) observed the impact of thymol on the integrity of the cell membrane of Cronobacter sakazakii by laser confocal technology. Similarly, Tian et al. (2021b) found that protocatechualdehyde can decrease the integrity of the cell membrane of Yersinia enterocolitica.
Studies have shown that ROS is related to membrane damage caused by lipid peroxidation, and destruction of biomolecular elements may cause cell membrane damage (Vatansever et al., 2013). The decrease in cell membrane integrity of eugenol-treated cells may be the result of elevated MDA content.
The results of FESEM showed that S. sonnei treated with eugenol changed from rod-shaped bacteria to amorphous bacteria, and the surface of the bacteria collapsed (Fig. 6). He et al. (2022) showed that the cell appearance of Listeria monocytogenes was blurred and the surface collapsed or even ruptured after treatment with linalool at a concentration of 1.5 μL/mL for 4 h. Guo et al. (2020) found that 0.2 mg/mL coenzyme Q0 treatment of C. sakazakii led to obvious depressions. The collapse of the cell may be due to lipid oxidation of the cell membrane after eugenol treatment of cells.
The intracellular ATP content was found to be decreased after S. sonnei strains were treated with eugenol (Fig. 7). Kang et al. (2020) showed that treatment with ferulic acid at 2 mg/mL for 30 min caused a decrease in the intracellular ATP content of S. flexneri. Song et al. (2020) found that citrus essential oil treatment at 0.5 mg/mL on S. aureus for 30 min decreased the intracellular ATP content. Studies have shown that the decrease in intracellular ATP content may be due to changes in cell membrane permeability, causing intracellular ATP leakage. Another way is to increase hydrolysis of ATP through proton power driven by ATPase (Kang et al., 2018; Gyorgy et al., 2020).
Hyperpolarization of the membrane potential of S. sonnei cells was induced by eugenol and showed a concentration-dependent effect (Fig. 8). Qian et al. (2020) used DiBAC4(3) to probe whether the cell membrane potential showed hyperpolarization in Klebsiella pneumoniae treated with eugenol. Previous studies have shown that membrane potential exhibits hyperpolarization due to disruption of pH homeostasis and ion efflux. ATP and membrane potential together explained the change of membrane permeability and dysfunction (Plasek et al., 2013).
Conclusions
In summary, this study confirmed that eugenol had antibacterial activity against S. sonnei. The intracellular ROS level in S. sonnei was increased by eugenol, resulting in oxidative damage to the cell membrane. Meanwhile, addition of eugenol caused intracellular ATP concentration decrease, cell membrane hyperpolarization, membrane integrity decrease, and cell morphology alteration.
These findings suggest that eugenol can be applied as a prospective antibacterial agent to prevent S. sonnei contamination of lettuce juice. Further study should be performed to investigate the effect of eugenol on the sensory quality of lettuce juice.
Availability of Data and Materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval
This study does not present experiments with humans or animals.
Footnotes
Authors' Contributions
R.S. was involved in conceptualization, investigation, formal analysis, project administration, writing—original draft, and writing—review and editing. X.B. was involved in resources, investigation, data curation, and visualization. Xiaoxiao Liu was involved in resources, data curation, and investigation. L.S. was involved in data curation, resources, and methodology. Xue Liu was involved in visualization, formal analysis, and resources. X.Z. was involved in methodology, supervision, and software. D.G. was involved in visualization, formal analysis, and validation. Y.W. and Y.C. were involved in supervision and project administration. C.S. was involved in project administration, supervision, and funding acquisition.
All authors have read and agreed to the published version of the manuscript.
Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This work was supported by the National Natural Science Foundation of China (32272445 and 31801659), the Key Research and Development Program of Shaanxi Province, China (2021NY-125), and the Natural Science Foundation of Guizhou Province (Qiankehe Foundation-ZK [2022] General 008).