
Abstract
Antimicrobial-resistant bacteria isolated from food animals pose a major health threat to the public on this planet. This study aimed to determine the susceptibility profiles of Escherichia coli isolated from cattle and pig fecal samples and investigate the molecular characteristics of extended-spectrum β-lactamase (ESBL)-producing E. coli using gene identification, conjugation, and Southern blot approach. Overall 293 E. coli were recovered from cattle (120 isolates) and pigs (173 isolates) in 7 provinces of Korea during 2017–2018. Ampicillin, chloramphenicol, streptomycin, and sulfisoxazole resistance rates were the highest in pigs' isolates (>60%, p ≤ 0.001) compared to that in cattle (3–39%). Multidrug resistance (MDR) was higher in pig isolates (73%) than in cattle (31%), and the MDR profile usually includes streptomycin, sulfisoxazole, and tetracycline. Resistance to critically important antimicrobials such as ceftiofur, colistin, and ciprofloxacin was higher in weaners than those from finishers in pigs. The qnrS gene was detected in 13% of the pig isolates. Eight isolates from pigs and one isolate from cattle were identified as ESBL-producers and ESBL genes belonged to bla CTX-M-55 (n = 4), bla CTX-M-14 (n = 3), and bla CTX-M-65 (n = 2). Notably, the bla CTX-M-65 and qnrS1 genes were found to be carried together in an identical plasmid (IncHI2) in two isolates from finisher pigs. The bla CTX-M-carrying isolates belonged to phylogenetic groups B1 (n = 4), B2 (n = 2), A (n = 2), and D (n = 1). The bla CTX-M genes and non-β-lactam resistance traits were transferred to the E. coli J53 recipient from seven bla CTX-M-positive strains isolated from pigs. The bla CTX-M genes belonged to the IncI1α, IncFII, and IncHI2 plasmids and are also associated with the ISEcp1, IS26, IS903, and orf477 elements. These findings suggested the possibility of bla CTX-M-carrying E. coli transmission to humans through direct contact with cattle and pigs or contamination of food products.
Introduction
Antimicrobial resistance has become a worldwide risk both in human and veterinary medicine. The frequent use of antimicrobials in animals and humans exerts selective pressure and contributes to the emergence of antimicrobial-resistant commensal bacteria. Thus, the level of resistance in commensal bacteria such as Escherichia coli is considered a good indicator of the selection pressure exerted by the use of antimicrobials in food-producing animals (Lei et al, 2010; Marshall and Levy, 2011). In addition, the bacterium contributes to a reservoir of resistance genes, which can spread to other commensals and a wide range of pathogenic bacteria through the horizontal transfer of mobile genetic elements (Marshall and Levy, 2011).
The widespread use of third-generation cephalosporins and another beta (β)-lactam antibiotics for treatment and prophylaxis of multiple diseases in food-producing animals has contributed to the emergence of extended-spectrum β-lactamase (ESBL)-producing E. coli (Reuland et al, 2014). Among the ESBLs, CTX-M (Cefotaximase-Munich) type β-lactamase enzymes are the most widespread, heterogeneous, and complex group of ESBL enzymes (Ramos et al, 2020). More than 238 members of the CTX-M family have been detected in different bacterial species. The most common CTX-M types in E. coli isolated from food-producing animals include the CTX-M-1 group (CTX-M-1 and CTX-M-15) and CTX-M-9 group (CTX-M-9 and CTX-M-14) in European countries, the CTX-M-2 group in South America and Japan, the CTX-M-9 group in China, and the CTX-M-1 group in the United States and North Africa (Day et al, 2019; Ramos et al, 2020).
In the Republic of Korea (Korea), bla CTX-M-1, bla CTX-M-14, and bla CTX-M-15 were the most frequent CTX-M gene types in E. coli isolated from cattle, chickens, and pigs (Jo and Woo, 2015; Tamang et al, 2013a; Tamang et al, 2013b). Recently, several studies have identified bla CTX-M-55 and bla CTX-M-65-carrying E. coli isolates from food-producing animals in Korea (Na et al, 2019; Park et al, 2019; Song et al, 2020a; Song et al, 2020b).
Although many studies are describing the antimicrobial resistance profiles of E. coli and the molecular characteristics of bla CTX-M-carrying E. coli from food-producing animals in Korea, continuous surveillance of the diversity of bla CTX-M genes is vital to understanding the evolution of the bla CTX-M genes and to prevent the dissemination of bla CTX-M-carrying E. coli in the food chain. Therefore, in the present study, we aimed to identify the antimicrobial resistance and molecular characteristics of ESBL-producing E. coli isolated from cattle and pigs in Korea.
Materials and Methods
Collection of samples
A total of 121 and 174 fecal swab samples were collected from cattle and pigs, respectively. Samples were obtained from 13 cattle and 12 pig farms (10–15 samples per farm) located in seven provinces of Korea in 2017 and 2018. However, the authors do not have information about the history of antimicrobial use in farms and the number of animals considered for this study. The samples were collected from young and adult age groups of animals from pig and cattle farms. We have collected samples from weaners (4–10 weeks) and finishers (6 months) pigs and calves (<6 months) and adult cattle (>2 years). No ethical approval was deemed necessary for this study. Collected samples were placed in ice-cooled containers, and immediately transported to the laboratory for further processing.
Isolation of E. coli
Isolation and identification of E. coli were performed as previously described by Song et al (2020a). In brief, swab samples were streaked on Eosin Methylene Blue agar (Becton Dickinson, Sparks, USA) and incubated for 20–24 h at 37°C. Suspected colonies with metallic sheen cultured again on MacConkey Agar (Becton Dickinson). Then suspected colonies with pink appearance were selected from each plate and subcultures on Tryptic Soy Agar (Becton Dickinson). E. coli was then identified by matrix-assisted laser desorption and ionization-time-of-flight mass spectrometry (MALDI-TOF; Biomerieux, Marcy L'Etoile, France). Only a single isolate per sample was considered for further processing.
Antimicrobial susceptibility testing
About 17 antimicrobials, including amoxicillin/clavulanic acid, ampicillin, cefepime, cefoxitin, ceftazidime, ceftiofur, chloramphenicol, ciprofloxacin, colistin, gentamicin, meropenem, nalidixic acid, streptomycin, sulfisoxazole, tetracycline, and trimethoprim/sulfamethoxazole, were tested for their antimicrobial susceptibility profile by the broth microdilution method (CLSI, 2018) using commercially available Sensititre plates KRNVP5F (Thermo Trek Diagnostics, Waltham, MA, USA). E. coli ATCC 25922 and E. coli ATCC 35218 were used as quality reference strains. Minimum inhibitory concentration (MIC) was interpreted according to the Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI VET01; CLSI 2018). However, streptomycin and ceftiofur MIC were interpreted according to the National Antimicrobial Resistance Monitoring System (NARMS, 2014). For the colistin MIC, the European Committee on Antimicrobial Susceptibility Testing guidelines has been followed (EUCAST Version 8.1. 2018). Multidrug resistance (MDR) was defined as resistance to three or more antimicrobial subclasses.
Identification of ESBL-producing E. coli and antimicrobial resistance genes
A double-disk synergy test was performed to identify ESBL production among ceftiofur-resistant isolates using cefotaxime–cefotaxime/clavulanic acid and ceftazidime–ceftazidime/clavulanic acid disks (Dickinson) (CLSI, 2018). A PCR assay and sequencing analysis were performed to identify isolates that carried bla CTX-M genes, genes encoding for AmpC families, and plasmid-mediated quinolone resistance (PMQR) genes, as previously described by Song et al (2020a).
Conjugation experiment
Conjugation was performed by the broth-mating method using sodium azide-resistant E. coli J53 as the recipient strain (Tamang et al, 2007). Transconjugants were selected on Muller–Hinton agar (Becton Dickinson), supplemented with sodium azide (150 μg/mL; Sigma) and cefotaxime (2 μg/mL; Sigma).
Characterization of ESBL-producing E. coli
The major plasmid incompatibility groups of E. coli strains carrying the bla CTX-M genes were determined by using a multiplex PCR-based replicon typing kit (Diatheva, Fano, Italy). PCR and Sanger sequencing were performed to investigate the genetic environment of the bla CTX-M genes using primers and PCR conditions described by Saladin et al (2002) and Eckert et al (2006). In addition, chromosomal DNA was digested with XbaI (TaKaRa Bio, Inc., Shiga, Japan), and pulsed-field gel electrophoresis (PFGE) analysis of the bla CTX-M-positive isolates was performed, as described by Gautom (1997). Further, phylogenetic grouping was performed by multiplex PCR according to the method of Clermont et al (2000).
Southern blot
Plasmids DNAs were extracted from bla CTX-M- and qnrS1-positive transconjugants using the Plus SV Minipreps DNA Purification Kit (Promega Corporation, MD, USA). The DNAs were subjected to agarose gel electrophoresis and then transferred onto a positively charged nylon membrane (Roche Diagnostics GmbH, Mannheim, Germany). A Digoxigenin DNA Labeling and Detection Kit (Roche Diagnostics) was used for hybridization analyses, according to the manufacturer's instruction.
Statistical analysis
The analysis of the antimicrobial resistance rates were performed using Rex software (Version 3.0.3; RexSoft, Inc., Seoul, Korea). A chi-square test was used to compare the observed resistance rates between young and adult age groups, and pigs and cattle. p-Values <0.05 were considered significant.
Results
Antimicrobial susceptibility
We identified 293 E. coli isolates from cattle and pigs: 120 isolates from cattle and 173 from pigs. Overall, E. coli isolates from pigs exhibited high resistance rates to most of the tested antimicrobials compared with those isolated from cattle (Table 1). Ampicillin, chloramphenicol, streptomycin, sulfisoxazole, and tetracycline resistance rates were high (>60%) in pig isolates, while cattle isolates presented very low or moderate resistance rates (3–39%) to these antimicrobials. The ciprofloxacin (33% vs. 1%), nalidixic acid (38% vs. 11%), gentamicin (22% vs. 2%), and trimethoprim/sulfamethoxazole (36% vs. 1%) resistance rates in pig isolates were significantly (p < 0.001) higher than that of cattle isolates. We observed ceftiofur resistance in 5% (8/173) of pig and 1% (1/120) of cattle isolates.
Minimum Inhibitory Concentrations (μg/mL) and Antimicrobial Resistance Rates for Escherichia coli Isolated from Pigs and Cattle
MIC50 and MIC90 concentrations (μg/mL) at which 50% and 90% of the isolates were inhibited, respectively.
Antimicrobial resistance rate.
Statistically significant association (p < 0.05).
MDR, multidrug resistance; MIC, minimum inhibitory concentration; ND, not determined.
On the contrary, a very low proportion (<5%) of cattle and pig isolates demonstrated resistance to the tested cephalosporins, amoxicillin/clavulanic acid, and colistin, while all the isolates were susceptible to ceftazidime and meropenem.
A slight difference was noted between the resistance rates of most of the tested antimicrobials isolated from young (weaned pigs and calves) and adult (finisher pigs and >2–year-old cattle) animals. The ciprofloxacin and nalidixic acid resistance rates were significantly (p < 0.001) high in isolates from weaner pigs (45% and 51%, respectively) compared with those from finishers (17% and 23%, respectively). We found no significant difference between the resistance rates of isolates recovered from adult cattle and calves, However, tetracycline and nalidixic acid resistance rates were relatively high in isolates from adult cattle (42% and 14%, respectively) compared with those from calves (34% and 6.4%, respectively) (Table 1; Supplementary Table S1).
In this study, 42% of cattle and 85% of pig isolates were resistant to at least one antimicrobial agent (Tables 2 and 3). The level of MDR was significantly higher in pig isolates (73%, 127/173) than in cattle isolates (31%, 37/120). The most frequent MDR patterns in cattle and pig isolates usually include resistance to streptomycin, sulfisoxazole, and tetracycline.
Frequent Resistance Patterns in Escherichia coli Isolated from Pigs (N = 173)
MDR (resistance to three or more antimicrobial subclasses).
AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; FEP, cefepime; FIS, sulfisoxazole; GEN, gentamicin; MDR, multidrug resistance; NAL, nalidixic acid; STR, streptomycin; SXT, trimethoprim/sulfamethoxazole; TET, tetracycline; XNL, ceftiofur.
Frequent Resistance Patterns in Escherichia coli Isolated from Cattle (N = 120)
MDR (resistance to three or more antimicrobial subclasses).
AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; FIS, sulfisoxazole; GEN, gentamicin; MDR, multidrug resistance; NAL, nalidixic acid; STR, streptomycin; SXT, trimethoprim/sulfamethoxazole; TET, tetracycline; XNL, ceftiofur.
Detection of PMQR gene
Among the PMQR gene, the qnrS gene was detected in 2% (2/99) and 27% (20/74) of isolates recovered from weaner and finisher pigs, respectively. In addition, we found two isolates from adult cattle that carried either the qnrS or the aac (6′)-lb-cr gene.
Identification of ESBL-producing E. coli and antimicrobial resistance gene carriage
Eight isolates from pigs (three from finishers and five from weaners) and one isolate from adult cattle were identified as ESBL-producers (Table 4). The ESBL-producing phenotypes isolated from finisher pigs were associated with bla CTX-M-65 (n = 2) and bla CTX-M-14 (n = 1), while isolates recovered from weaner pigs carried bla CTX-M-55 (n = 4) and bla CTX-M-14 (n = 1). All but one of the bla CTX-M-carrying isolates from pigs were recovered from the same farm in Gangwon province. The only ESBL-producing isolate from adult cattle belonged to bla CTX-M-14.
Molecular Characterization of Extended-Spectrum β-Lactamase-Producing Escherichia coli Isolated from Pigs and Cattle
The underlined resistance markers were transferred to the recipient Escherichia coli J53 strain by conjugation.
CHL, chloramphenicol; CIP, ciprofloxacin; FIS, sulfisoxazole; FOX, cefoxitin; GEN, gentamicin; NAL, nalidixic acid; PMQR, plasmid-mediated quinolone resistance; STR, streptomycin; SXT, trimethoprim/sulfamethoxazole; TET, tetracycline; XNL, ceftiofur; +, transferred; −, Not transferred.
Characteristics of bla CTX-M-harboring E. coli
The bla CTX-M-harboring isolates exhibited resistance to multiple non-β-lactam antimicrobials (Table 4). The bla CTX-M-65 and qnrS1 genes were carried together in two isolates from finisher pigs. Transfer of the bla CTX-M genes to recipient E. coli J53 was demonstrated in seven of the bla CTX-M-positive strains isolated from pigs (three bla CTX-M-55, two bla CTX-M-14, and two bla CTX-M-65). Also, we noted the cotransfer of non-β-lactam antibiotic resistance traits along with bla CTX-M genes.
We identified three plasmid replicon types in the transconjugants from pigs: IncI1α, IncFII, and IncHI2. IncI1α and IncHI2 were the most common plasmid replicon types (Table 4). The bla CTX-M-harboring transconjugants presented diverse genetic environments, namely bla CTX-M-65-IS903 (n = 2), bla CTX-M-14-IS903 (n = 2), ISEcp1-IS26-bla CTX-M-55-orf477 (n = 2), and each of the ISEcp1-bla CTX-M-14-IS903, ISEcp1-bla CTX-M-55-orf477, and IS26-bla CTX-M-55-orf477 elements. The Southern blotting analysis confirmed that the bla CTX-M-65 and qnrS1 genes were located on a single plasmid (IncHI2 ≈ 70 kb) in both transconjugants (Fig. 1). In addition, strain17-S-L-F-15 carried an additional conjugative plasmid (≈60 kb).
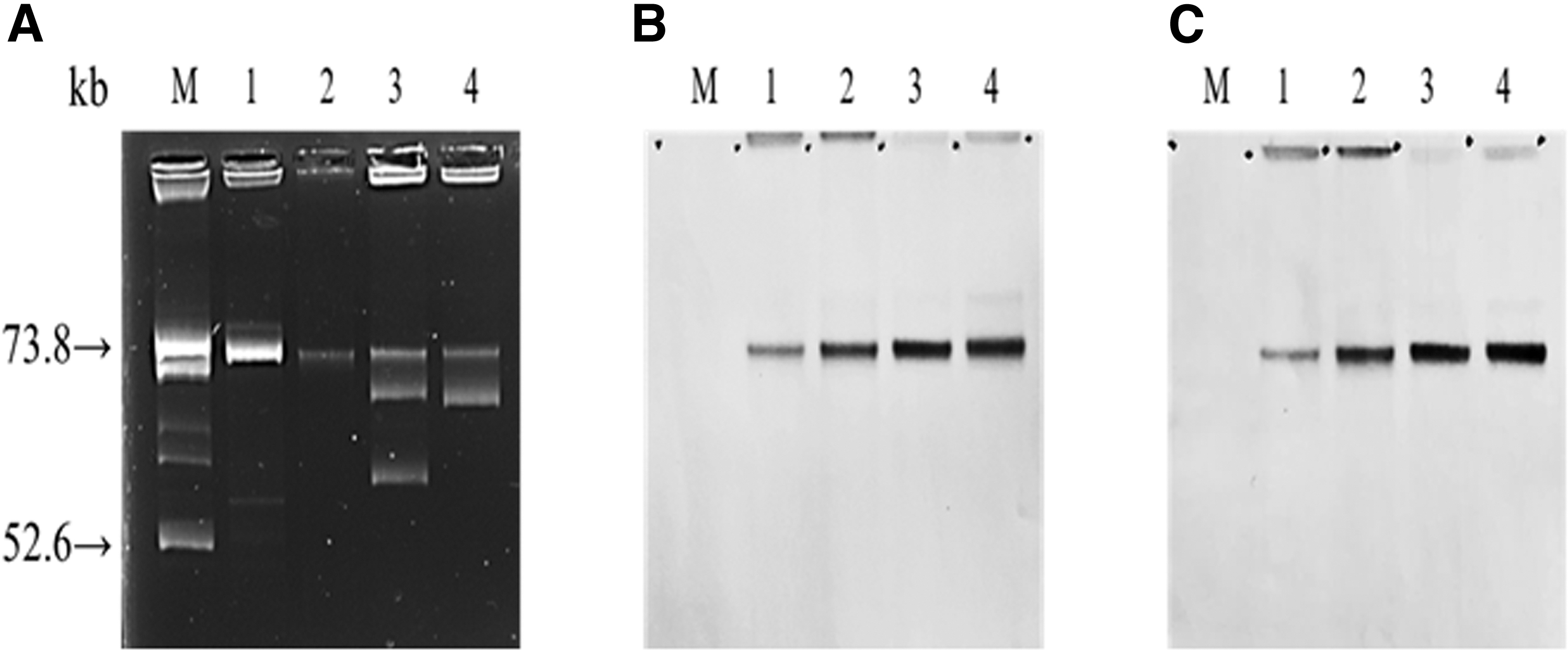
Plasmid profiles of two bla
CTX-M-65 and qnrS1-positive clinical isolates and their transconjugants.
PFGE analysis of the bla CTX-M-carrying strains demonstrated eight arbitrary pulsotypes (Supplementary Fig. S1). Except for the two bla CTX-M-55-carrying strains from weaner pigs in farm L (17-S-L-F-2 and 17-S-L-F-6), all strains exhibited heterogeneous PFGE profiles. Phylogenetic analysis demonstrated that four bla CTX-M-carrying isolates belonged to subgroup B1 (three bla CTX-M-55 and one bla CTX-M-14), subgroup A (two bla CTX-M-14), subgroup B2 (two bla CTX-M-65), and subgroup D (bla CTX-M-55).
Discussion
Investigating antimicrobial resistance of indicator commensal bacteria from food-producing animals provides important data on the occurrence of resistance (EFSA, 2015). In agreement with previous reports in Korea (Do et al, 2020; Do et al, 2019; Kyung-Hyo et al, 2020) and other countries (Donkor et al, 2012; Hayer et al, 2020; Kidsley et al, 2018; Zhang et al, 2017), a significantly higher proportion of E. coli isolates from pigs were resistant to ampicillin, chloramphenicol, streptomycin, sulfisoxazole, and tetracycline. However, consistent with previous reports in other countries (Dolejská et al, 2008; Enne et al, 2008; Ho et al, 2011; Lim et al, 2014), cattle isolates presented low or moderate resistance rates to these antimicrobials.
Conversely, 23–39% of E. coli isolates from pigs in this study were resistant to ciprofloxacin, gentamicin, nalidixic acid, and trimethoprim/sulfamethoxazole. Several studies in Asia (Do et al, 2020; Kyung-Hyo et al, 2020; Lei et al, 2010; Xu et al, 2015) and Europe (de Jong et al, 2012; Enne et al, 2008) reported highly variable (0–95%) and relatively low resistance rates (≤11%) to ciprofloxacin, gentamicin, nalidixic acid, and trimethoprim/sulfamethoxazole in E. coli isolated from pigs and cattle, respectively. The high antimicrobial resistance rates coincide with the heavy use of antimicrobials in pig industry (APQA, 2020). Another possible reason may be the treatment regimens between pigs and cattle (group vs. individual) (Hanon et al, 2015).
A small percentage of isolates (≤6%) from cattle and/or pigs in this study demonstrated resistance to critically important antimicrobials such as amoxicillin-clavulanic acid, cefepime, cefoxitin, and colistin. Our finding concurred with previous studies in Korea (Do et al, 2020; Lim et al, 2014) and some European countries (de Jong et al, 2012; Enne et al, 2008). Thus, E. coli isolates resistant to antibiotics that are considered critically important for humans can spread to humans via direct contact or through the food chain (Ho et al, 2011).
In the present study, MDR was considerably more prevalent among pig isolates (73%) than cattle isolates (31%). Similarly, previous reports in Korea and other Asian countries (Do et al, 2020; Ho et al, 2011; Xu et al, 2015; Yamamoto et al, 2013) identified very high MDR rates in pig isolates (56–96%) compared with cattle isolates (18–25%). Concordant with the above reports and a study in Europe (EFSA, 2015), profiles of MDR usually include streptomycin, sulfisoxazole, tetracycline, and/or ampicillin. This could be associated with the colonization of tetA, sul1, aac(3)Ia, and aac(3)IV genes on a single mobile genetic element (Boerlin et al, 2005).
Overall, E. coli isolated from pigs exhibited significantly (p < 0.05) high resistance rates to most of the tested antimicrobials compared with those isolated from cattle. Our results may reflect the differences in the amounts of antimicrobials used in pig and cattle husbandry. Although data on the history of antimicrobial usage in Korean livestock farms are lacking, the total annual sales of antimicrobials for cattle and pig farms during the study period were about 90 and 500 tons, respectively. Indeed, the annual average sale of penicillin, phenicol, sulfonamides, and tetracyclines for the pig industry during the study period was much higher than the amount sold for the cattle industry (APQA, 2020).
However, caution must be exercised when comparing and contrasting antimicrobial resistance rates and MDR profiles among studies because of the differences in the health status of animals, locally approved antimicrobials, farm management systems, history of antimicrobial use, and methodologies used for resistance detection and resistance breakpoints.
About 13% (22/173) of pig isolates, predominantly from finisher pigs, carried the qnrS gene, while two isolates from adult cattle were positive for qnrS or aac (6′)-lb-cr genes. The qnrS gene has been identified in several enterobacterial isolates from food-producing animals in many countries, including Korea (Pungpian et al, 2021; Song et al, 2020a; Szmolka et al, 2011; Yue et al, 2011). Although the PMQR genes confer low-level resistance to fluoroquinolones, they are sufficient to facilitate the selection of mutants with higher levels of resistance (Cambray et al, 2010).
Dissemination of ESBL-producing Enterobacteriaceae has become a major global health challenge. The ESBL genes identified in this study belonged to bla CTX-M-14, bla CTX-M-55, and bla CTX-M-65. Despite geographical variability in the distribution of CTX-M ESBLs, the bla CTX-M-14, bla CTX-M-55, and bla CTX-M-65 were frequently detected in ESBL-producing E. coli isolated from human, food, and companion animals in Korea (Hong et al, 2019; Park et al, 2019; Song et al, 2020a; Tamang et al, 2013b) and other Asian countries (Bevan et al, 2017; Zhang et al, 2014). The bla CTX-M genes carrying E. coli can spread between animals and humans through the food chain.
Some reports have found identical sequence types and plasmid incompatibility groups in E. coli from food-producing animals, animal products, and humans (Day et al, 2019; Irrgang et al., 2017). Notably, the emergence of bla CTX-M-55, a variant of bla CTX M-15, in E. coli isolated from food-producing animals in Korea should be given due consideration because this CTX-M type is associated with broad-spectrum β-lactamase activity (Faheem et al, 2013).
In this study, all ESBL-producing isolates were resistant to multiple antimicrobials. In addition, the transfer of bla CTX-M genes was observed in 78% (7/8) of the bla CTX-M-carrying isolates, which was higher than the findings of Tamang et al (2013b) (63%) in isolates from cattle and pigs in Korea, and Liu et al (2018) (63%) in isolates from pigs in China. Notably, the qnrs1 gene was also cotransferred along with the bla CTX-M-65 genes on the same plasmid in two isolates from finisher pigs. The bla CTX-M genes are usually encoded on transmissible plasmids, which are highly mobile and carry other resistance genes, especially the PMQR genes and genes encoding 16S rRNA methyltransferases, allowing possible coselection and dissemination of multidrug-resistant E. coli (Ramos et al, 2020).
Commensal bacteria are considered a potential reservoir of mobile genetic elements (O'Brien, 2002). In this study, we identified three plasmid replicon types: IncI1α, IncFII, and IncHI2. Epidemiological studies have demonstrated a close linkage between bla CTX-M genes and the IncF, IncI, IncN, IncHI2, IncL/M, and IncK plasmid groups (El Salabi et al, 2013; Singh et al, 2018). In addition, several reports in Korea and other countries have revealed the association of bla CTX-M-14, bla CTX-M-55, and bla CTX-M-65 genes with diverse plasmid types including IncFII, IncI1, IncFIB, IncN, and IncA/C replicon types (Lupo et al, 2018; McGann et al, 2016; Zheng et al, 2012). Plasmids facilitate the spread of ESBL-associated genes by horizontal gene transfer to other commensal or pathogenic bacteria (Geser et al, 2011).
Consistent with previous studies (Park et al, 2019; Song et al, 2020a; Zheng et al, 2012), the ESBL-producing isolates belonged to different phylogenetic groups (A, B1, B2, and D). Most commensal strains belong to phylogroups A and B1, whereas virulent strains of E. coli belong primarily to phylogroups B2 and D (Xu et al, 2015). Cortés et al (2010) and Cao et al (2011) have identified E. coli isolates belonging to phylogroups B2 and D in humans with urinary tract and other extraintestinal infections. Therefore, the detection of virulent strains of E. coli in healthy pigs constitutes a public health concern.
The bla CTX-M genes are often associated with insertion sequences (ISs), which further contribute to their dissemination via diverse genetic platforms (Cantón et al, 2012). The ISs in this study were noted upstream (ISEcp1 and IS26) and downstream (IS903) of the bla CTX-M-carrying isolates. Reports showed that ISs located upstream and downstream of the bla CTX-M genes might have facilitated the widespread transmission of bla CTX-M-14, bla CTX-M-55, and bla CTX-M-65 genes in E. coli from food animals (Liao et al, 2015; Lupo et al, 2018; Na et al, 2019; Tamang et al, 2013b). Among the identified IS elements, ISEcp1 is frequently associated with the mobilization of the bla CTX-M genes (Zhao and Hu, 2013).
PFGE analysis revealed that bla CTX-M-positive isolates were heterogeneous, except for the two specific clonal strains. This suggests that the bla CTX-M genes are spreading through bacterial populations mainly through horizontal transmission or that bla CTX-M-positive bacteria are entering pork industries from diverse sources.
Conclusions
In summary, our study showed that the cattle and pigs were potential carriers of ESBL-producing E. coli strains. The bla CTX-M-carrying pathogenic strains were resistant to multiple antibiotics, including those ranked as critically important for humans. Antimicrobial-resistant strains of E. coli can be transferred from food-producing animals to humans through the food chain if humans consumed or come in contact with these animals. Therefore, the prudent use of antimicrobials and extensive monitoring of food-producing animals are needed to limit the emergence and dissemination of ESBL-producing E. coli.
Footnotes
Authors' Contributions
S.-K.L., D.C.M., and S.-S.Y. designed the experiments; H.-J.S., H.Y.K., J.-H.C., S.-H.N., S.-J.K., and N.B. carried out the experiments; A.F.M. and H.-J.S. completed the original draft of article and the statistical analysis; A.F.M. and N.B. revised the article; S.-S.Y., D.C.M., and S.-K.L. were involved in funding acquisition and supervision. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Animal and Plant Quarantine Agency, Ministry of Agriculture, Food, and Rural Affairs, Republic of Korea [Grant No. N-1543081-2017-24-01]; and the Research of Korea Centers for Disease Control and Prevention [Grant No. 2017NER54070]. The sponsor did not play any role in study design; in the collection, analysis, and interpretation of data; in the writing of the report; or in the decision to submit the article for publication.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.