
Abstract
Microbial contamination in foods could lead to illnesses and substantial losses in both food industry and public health sectors. Rapid detection of microbial hazards (i.e., pathogens, hygiene indicator microorganisms) can accelerate surveillance and diagnostic processes reducing transmission and minimizing undesirable consequences. This study developed a multiplex PCR (m-PCR) for the detection of six common foodborne pathogens and hygiene indicators using specific primers for uidA of Escherichia coli, stx2 of Escherichia coli O157:H7, invA of Salmonella spp., int of Shigella spp., ntrA of Klebsiella pneumoniae, and ail of Yersinia enterocolitica and Yersinia pseudotuberculosis. Sensitivity of the m-PCR was 100 fg or ∼20 bacterial cells. Each primer set amplified only the targeted strain, and specificity was demonstrated by lack of nonspecific bands with DNA from 12 other bacterial strains. Following ISO 16140-2:2016, the relative limit of detection of the m-PCR was comparable to that of the gold-standard method; however, the processing time was five times faster. The m-PCR was applied to detect the six pathogens in 100 natural samples (50 pork meat and 50 local fermented food samples) and compared to results of the gold-standard method. Positive cultures for Klebsiella, Salmonella, and E. coli were 66%, 82%, and 88%, respectively, of meat samples and 78%, 26%, and 56%, respectively, of fermented food samples. Escherichia coli O157:H7, Shigella, and Yersinia were not detected in any of the samples by both standard and m-PCR methods. The developed m-PCR assay showed comparable results with the traditional culture technique proving its rapid and reliable detection of six foodborne pathogens and hygiene indicators in food.
Introduction
Effective surveillance for foodborne diseases has raised a major global public health concern over increasing rates of infection, underestimation of the problem, and the spread of drug resistant strains. In 2021, 538,608 cases of foodborne disease were reported in Thailand, with an associated morbidity rate of 813.96/100,000 population (Bureau of Epidemiology, 2021). During 2017, the Foodborne Diseases Active Surveillance Network (Foodnet) reported that the top five causes of infection in the United States included Salmonella spp., Shigella spp., Shiga toxin–producing Escherichia coli, and Yersinia spp. (Marder et al., 2018).
The abovementioned problematic species belong to Enterobacterales order. Salmonella spp. is the most common foodborne pathogen found in contaminated food. In Thailand the involved foods are notably eggs, dairy products, and meat (Ananchaipattana et al., 2012). E. coli, used as a fecal indicator index for food or drink, includes both commensal and pathogenic bacteria depending on strain (Jang et al., 2017). Pathogenic E. coli found in food can be divided into five groups: Shiga toxin–producing E. coli or enterohemorrhagic E. coli (STEC/EHEC), enteropathogenic E. coli, enteroinvasive E. coli, enteroaggregative E. coli, and enterotoxigenic E. coli.
Outbreaks of Escherichia coli O157:H7, one serotype of STEC/EHEC, caused by contaminated vegetables or beef were commonly reported in Europe, America, and Canada during 2006–2020 (CDC, 2020). A high detection rate (8%) of STEC was found in raw beef sold in markets in southern Thailand in 2016, a surprising increase from year 2015 (Sirikaew et al., 2016). In the past decade, non-O157 STEC serotypes have emerged and become more widespread around the world. These serotypes (namely O26, O103, O111, and O145) have also shown to cause hemolytic uremic syndrome in invasive cases (Bosilevac and Koohmaraie, 2011; Eichhorn et al., 2015; Hoyle et al., 2021).
Shigella spp. is a major cause of acute diarrhea in children and a serious problem in developing countries (Bodhidatta et al., 2010). Yersinia enterocolitica infections can cause gastrointestinal disease (yersiniosis) acquired by eating contaminated raw meat and poultry products (Rahman et al., 2011). Klebsiella spp., although uncommonly identified as a foodborne pathogen, in foods are good indicators for food hygiene, and multidrug-resistant Klebsiella pneumoniae found in raw and ready-to-eat foods have been shown to be a potential pool of antibiotic resistance genes (Hartantyo et al., 2020). Rising prevalence of K. pneumoniae in foods worldwide, that is, Singapore, Denmark, France, and Greece, is alarming in that the bacterium might not be only an indicator of poor hygiene given the strict food safety measures in these industrialized countries (Rodrigues et al., 2022; Theocharidi et al., 2022). Detection and elimination of these pathogens and indicators in food supply chain can significantly improve food safety as a whole.
Conventional culture is the gold-standard method to identify foodborne pathogenic and indicator bacteria. However, the process is multistep and time consuming due to long incubation times, laborious, and requires several biochemical tests. To better control transmission and lessen undesirable effects, a rapid and sensitive diagnostic method is needed to simultaneously detect and identify the most common foodborne pathogens and indicators. Monitoring for the bloody diarrhea-causing bacterium, EHEC serotype O157:H7, and for the underestimated gastrointestinal K. pneumoniae should be done in parallel to avoid undesirable loss.
A molecular method, multiplex PCR (m-PCR), provides an efficient alternative because it allows rapid and low-cost detection with high specificity and sensitivity and could be made readily available. m-PCR is among the approved techniques exploited to detect a large group of foodborne bacteria and shows comparable results with conventional culture methods while having a significantly shorter processing time (Huang et al., 2018). Several attempts to develop m-PCR assays that can detect and differentiate common foodborne pathogens have been made (Babu et al., 2013; Lee et al., 2014; Lei et al., 2008; Nguyen et al., 2016; Zhang et al., 2009) but, to our knowledge, no m-PCR focused on the six commonly found foodborne pathogens and indicators.
The aim of this study was, therefore, to develop the m-PCR which simultaneously targets E. coli, Escherichia coli O157:H7, Salmonella spp., Shigella spp., K. pneumoniae, and Y. enterocolitica. The six-plex PCR target genes previously reported to be specific to corresponding species and subspecies are uidA of E. coli (Anbazhagan et al., 2011), stx2 of Escherichia coli O157:H7 (Li et al., 2017; Van Giau et al., 2016), invA of Salmonella spp. (Chiu and Ou, 1996), int of Shigella spp. (Ranjbar et al., 2016), ntrA of K. pneumoniae (Anbazhagan et al., 2011), and ail of Y. enterocolitica and Yersinia pseudotuberculosis (Wiemer et al., 2011). The m-PCR was then validated in-house and subsequently used in natural sample detection in comparison to the culture technique.
Materials and Methods
Bacterial strains and culture conditions
The bacterial strains used in this study are listed in Table 1. For the m-PCR development, sensitivity testing, and validation, the following strains were used: Escherichia coli DMST 4212 (ATCC 25922), Escherichia coli O157:H7 MT, Klebsiella pneumoniae ATCC 27736, Salmonella Typhimurium DMST 562 (ATCC 13311), and Shigella dysenteriae DMST 15111. Bacterial strains were propagated and grown on tryptic soy agar and incubated at 37°C. After overnight incubation, single colonies were inoculated into tryptic soy broth and incubated at 37°C with shaking at 200 rpm for 18 h.
Bacterial Strains Used in This Study
These strains were used for the m-PCR development, sensitivity testing, and validation.
ATCC, American Type Culture Collection; DMST, Department of Medical Sciences Thailand; ETEC, enterotoxigenic E. coli; LT, Lilleengen type; m-PCR, multiplex PCR; MT, Collection of Department of Medical Technology, Thammasat University.
DNA template preparation
For all strains except Y. enterocolitica, genomic DNA (gDNA) was extracted from 1 mL of an overnight culture using an E.Z.N.A Bacterial DNA Isolation Kit (Omega Bio-Tek, Norcross, GA, USA) following the manufacturer's protocol. After purification, the qualities (A260/230 and A260/280) and concentration (ng/μL) of gDNA were determined using a spectrophotometer (DeNovix, Wilmington, DE, USA). For Y. enterocolitica, the 700 bp synthetic oligonucleotide sequence of the ail gene from Yersinia enterocolitica ATCC27729 was used as the DNA template.
m-PCR primer design
Two pairs of primers for detection of Salmonella spp. (Chiu and Ou, 1996) and Y. enterocolitica (Wiemer et al., 2011) were obtained from literature reviews; four pairs of primers for detection of E. coli, Escherichia coli O157:H7, Shigella spp., and K. pneumoniae were designed in the present study using BioEdit 7.2 program. The sequences are shown in Table 2. Size and position of each PCR product were evaluated by single PCR amplification.
Primers Used in This Study
The m-PCRs consisting of six primer sets were performed in a total volume of 25 μL containing 0.5 μM of each forward and reverse primer, 1 ng of each DNA template, and 1 × PCR Master Mix Kit (BiotechRabbit, Berlin, Germany). The m-PCR amplification was optimized by varying the annealing temperature from 50°C to 60°C in the Applied Biosystems SimpliAmp™ Thermal Cycler PCR (Life Technologies, South San Francisco, CA, USA). The amplification steps were followed by: 95°C for 2 min, 30 cycles each of 94°C for 30 s, 50–60°C for 45 s, and 72°C for 45 s, with a final extension at 72°C for 5 min. The PCR products were separated using 1.5% agarose gel electrophoresis, stained with 1 × GelRed (Merck, Darmstadt, Germany), and visualized under a UV transilluminator (Bio-Rad, Hercules, CA, USA).
Sensitivity and specificity tests of m-PCR
m-PCR sensitivity was determined by a serial dilution method. The DNA template of six species was 10-fold serially diluted from 100 pg to 1 fg (of each species) and applied into the m-PCR. m-PCR specificity was determined by testing the amplification on the genomes from 12 species of both Gram-negative and -positive bacteria (Table 1). The m-PCR condition for both sensitivity and specificity tests was 95°C for 2 min, 30 cycles each of 94°C for 30 s, 58°C for 45 s, and 72°C for 45 s, with a final extension at 72°C for 5 min.
In-house validation of m-PCR
Following ISO 16140-2:2016, the level of detection at 50% (LOD50) of the developed m-PCR (LOD50 m-PCR) was determined and compared with the standard method (LOD50 Ref) using six samples of canned processed meat. Each analysis was replicated thrice along with the reference method for the six pathogens as previously described.
The process was performed as follows: (1) taking 25 g of each sample into sterile plastic bags and adding 225 mL pre-enrichment broth (brain-heart infusion for E. coli, Shigella broth for S. dysenteriae, buffered peptone water for Escherichia coli O157:H7, Salmonella Typhimurium, and K. pneumoniae, and peptone sorbitol bile salt for Y. enterocolitica); (2) spiking an inoculum of bacteria at 100 μL (low concentration, ∼0.7 colony-forming unit [CFU]/mL), 500 μL (medium concentration, ∼4 CFU/mL), and 1 mL (high concentration, ∼13 CFU/mL) of working stock culture (at ∼10 CFU/mL) into the sample, the bacterial concentration of spike inoculum was calculated using aerobic plate count technique; (3) incubating at appropriate conditions following the standard methods [BAM 2017 for E. coli (Feng et al., 2020), ISO21567:2004(E) for S. dysenteriae, AOAC Official Method, 2017 for Escherichia coli O157:H7, ISO6579-1:2017(E) for Salmonella Typhimurium, BAM 2017 for K. pneumoniae (Feng et al., 2020), and ISO10273:2017 for Y. enterocolitica].
The pre-enrichment samples were tested with the m-PCR and compared with standard methods; LOD50 was calculated using the PODLOD calculation program. POD is probability of detection and LOD is limit of detection. The relative limit of detection (RLOD) was calculated using this equation:
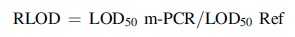
Prevalence of six foodborne pathogens as determined by standard methods and m-PCR
One hundred samples (50 pork meats and 50 local fermented food [i.e., fermented pork, fish, or soybean typically consumed raw]) were collected from five different regions (north, northeast, east, south, and central) of Thailand. Detection of six bacterial strains was carried out following standard methods. Food samples (25 g each) were transferred into sterile plastic bags with 225 mL pre-enrichment broth, mixed with a stomacher, and incubated under appropriate conditions following standard methods. Enrichment broth was used for both direct culture and m-PCR techniques.
Total gDNA was extracted from pre-enrichment broths obtained during standard culture techniques by a boiling method (Ngamwongsatit et al., 2008). The following PCR cycling condition was used: a denaturation step at 95°C for 2 min, 30 cycles of 30 s at 94°C, 45 s at 58°C, 45 s at 72°C, and finally 5 min at 72°C. Spiked samples were used as positive controls. The sensitivity, specificity, accuracy, positive predictive value, and negative predictive value of the developed m-PCR were calculated from the results [including true positives (a), false negatives (b), false positives (c), and true negatives (d)] with the MedCalC program (
Results and Discussion
m-PCR primers and assay development using purified templates
An efficient and sensitive m-PCR was developed to detect and identify six commonly found foodborne pathogens. Two primer pairs were obtained from literature reviews: Sal244F and Sal244R, specific primers to invasion protein A of Salmonella spp. (Chiu and Ou, 1996); Yer185F and Yer185R, specific primers to attachment-inversion locus protein of Y. enterocolitica (Wiemer et al., 2011). Ecol600F and Ecol600R, specific primers to beta-glucuronidase gene of E. coli, were designed in our study. Three primer pairs amplifying the integrase gene (int) of Shigella spp., Shiga toxin 2 (stx2) of Escherichia coli O157:H7, and nitrogen regulation gene (ntrA) of K. pneumoniae were also designed in this study.
As shown in Figure 1, all six primer pairs were combined in m-PCRs, and a total of six different sized PCR products were amplified in each reaction with the optimal annealing temperature at 58°C. The amplicon sizes of uidA, int, stx2, invA, ail, and ntrA are 600, 438, 321, 244, 185, and 130 bp, respectively, which can be easily discriminated by gel electrophoresis. Only expected positive bands were observed after m-PCRs were performed on one template DNA. Using a mixed DNA template from the six pathogens, six bands were observed in the gel representing a successful m-PCR.

Agarose gel electrophoresis analysis of m-PCR amplification products. Lane M, 100 bp DNA marker; Lane P, mixed DNA templates of the six pathogens. DNA samples from specific pathogens or hygiene indicators are indicated. m-PCR, multiplex PCR.
Although m-PCR assays that can differentiate common foodborne pathogens have already been developed (Babu et al., 2013; Lee et al., 2014; Lei et al., 2008; Nguyen et al., 2016; Zhang et al., 2009), an m-PCR designed for these six foodborne pathogens or hygiene indicator bacteria has not been done. Other quantitative m-PCRs (Liu et al., 2019; Van Lint et al., 2015) or more sophisticated techniques such as gene chip (Feng et al., 2016), m-PCR coupled with analyzer (Zhou et al., 2013) or with high-performance liquid chromatography (Xu et al., 2012) are reported. However, these techniques must be run in expensive instruments which are unaffordable for many laboratories and field studies.
Sensitivity and specificity of the developed m-PCR
The m-PCR was tested with 10-fold serially diluted, mixed DNA template to determine the sensitivity. The detection sensitivity of the m-PCR was 100 fg or ∼20 bacterial cells for all targeted strains (Fig. 2). To evaluate the primer specificity, the six pairs of primers were tested by PCR on DNA templates isolated from 12 strains of other bacteria (Citrobacter freundii, Enterobacter cloacae, Pseudomonas aeruginosa, Proteus vulgaris, Vibrio cholerae, Vibrio parahaemolyticus, Bacillus cereus, Bacillus subtilis, Enterococcus faecalis, Listeria monocytogenes, Staphylococcus aureus, and Staphylococcus epidermidis). The results showed that each primer pair amplified only the targeted strain and did not cross-react with DNA from any of the other 12 bacterial strains nor produce nonspecific bands and thus demonstrated its specificity (data not shown). The specificity of the diagnostic method provided confidence that the assay would avoid false positive results and unexpected signals.

Sensitivity of the m-PCR assay. Lane M, 100 bp DNA marker; Lane P, mixed DNA template of the six pathogens; Lane N, negative control or no DNA template.
In-house validation of m-PCR
The LOD50 of the developed m-PCR was evaluated with six samples of canned processed meat inoculated with three concentrations (low, medium, and high) of each targeted bacterium, and the presence of the six foodborne bacteria was assessed by standard culture methods. Negative controls used for validation were samples without bacterial contamination. The pre-enrichment broth from each sample was tested with the developed m-PCR. The m-PCR results were analyzed and are shown in Table 3. Using the PODLOD program, the LOD50 values of both the standard culture method and the m-PCR were calculated. The RLOD of the m-PCR was found to be comparable to that of the gold-standard method; however, the processing time was five times shorter. Thus, the m-PCR appeared suitable for a quick in-house analysis for contamination detection and prompt response.
Validation Results of Multiplex PCR
RLOD = LOD50 culture/LOD50 m-PCR.
CFU, colony-forming unit; EHEC, enterohemorrhagic E. coli; LOD50, level of detection at 50%; RLOD, relative limit of detection.
Prevalence of six foodborne target organisms as determined by standard methods and m-PCR
To assess the detection capability of the m-PCR in natural food samples, 100 meats and fermented foods from five different parts of Thailand were sampled and used to determine the prevalence of the six foodborne pathogens by both m-PCR and standard culture methods (Table 4).
Frequency of Six Pathogens in Food Samples as Determined by Standard Methods and Multiplex PCR
Meat refers to fresh pork, and fermented food refers to local fermented pork, fish, or soybean typically consumed raw.
Using BAM 2017, E. coli was detected in 72 (72%) samples, including 44 (88%) from meat and 28 (56%) from fermented foods. K. pneumoniae and Salmonella spp. were found in 72 (72%) samples (33 meat and 39 fermented food samples) and 54 (54%) samples (41 meat and 13 fermented food samples), respectively. Presumptive colonies were randomly chosen and confirmed for their biochemical characteristics. Total DNA was extracted from pre-enrichment broths obtained during standard culture and used as templates for m-PCR detection.
Prevalence of E. coli, K. pneumoniae, and Salmonella spp. in 100 meat and fermented food samples, as determined by the developed m-PCR, was 81%, 70%, and 44%, respectively. From 100 samples, there was no Escherichia coli O157:H7, Shigella spp., nor Y. enterocolitica detected by either culture technique or m-PCR. The m-PCR assay showed comparable results with those of the traditional culture technique proving its capacity to rapidly and reliably detect these six target organisms in foods. In addition, the developed m-PCR was able to detect multiple target organisms per food samples, for example, 30 (of 50) meat samples and 8 (of 50) fermented foods were positive for all E. coli, K. pneumoniae, and Salmonella. Sixty-four food samples (of 100) were positive for more than one target organism as detected by m-PCR.
Discrepancies in positive samples between culture and m-PCR techniques could be due to multiple aspects: (1) the lower gDNA release during boiling extraction, (2) bacterial cell wall physiology (particularly high capsule production in K. pneumoniae), and (3) nature of food sample (i.e., less positive samples were identified for Salmonella and K. pneumoniae by m-PCR than by culture).
Analysis by the MedCalC program (Table 5) showed that, in combined sample types, the developed m-PCR had high (>85%) sensitivity, accuracy, as well as high positive and negative predictive values for detection of E. coli. The uid primers still made up the best detection set in comparison to invA and ntrA. For Salmonella spp. detection, the pitfalls are relative sensitivity and negative predictive value, suggesting that this invA primer set is excellent in high contamination detection. Unfortunately, the ntrA primer set for K. pneumoniae showed only positive predictive value in acceptable range. The accuracy and negative predictive value by m-PCR for detection of K. pneumoniae were the lowest compared with that of E. coli and Salmonella spp. By taking a closer look at the calculated parameters, we have seen the effects of food types, that is, fermented food samples lowered the sensitivity in E. coli, Salmonella spp., and Klebsiella spp. (i.e., down to 96.13%, 46.15%, and 52.63%, respectively). This finding confirms the discrepancies in positive samples between culture and m-PCR techniques in that nature of food sample plays a role in detection.
The Relative Sensitivity, Specificity, Accuracy, Positive Predictive Value, and Negative Predictive Value of Developed Multiplex PCR Analyzed by MedCalC Program
NPV, negative predictive value; PPV, positive predictive value.
In conclusion, our results showed that 72% of specimens were contaminated by E. coli, more so in meats. High prevalence of E. coli is commonly found in food samples collected in Thailand (Minami et al., 2010), pointing out the importance of good hygiene and safe handling of raw foods. Salmonella spp. were detected at 54%, notably in meats from both retail and wholesale markets, while contamination of fermented foods was comparatively low. Ananchaipattana et al. (2012) revealed that ∼80% of meat samples were contaminated with Salmonella spp. while only 9% of fermented food samples were contaminated.
Prevalence of K. pneumoniae in food is underestimated in Thailand. This bacterium can contaminate various foods and contribute to food spoilage and human disease. In a recent study, K. pneumoniae strains were isolated from 9.9% of raw and cooked food samples, with fresh raw chicken showing the highest frequency (13.8%) (Guo et al., 2016). K. pneumoniae was found to contaminate 21% of raw and ready-to-eat foods (Hartantyo et al., 2020), while in the present study it was up to 72%. Contamination of K. pneumoniae in food might not cause any harm, but nonetheless it could act as a reservoir of antimicrobial-resistant K. pneumoniae (Guo et al., 2016). We developed an m-PCR assay which would provide a useful tool for monitoring and ensuring safety along the food chain with high sensitivity and specificity.
Footnotes
Authors' Contributions
N.N.: Conceptualization, funding acquisition, investigation, methodology, validation, writing–review and editing. S.C.: Conceptualization, funding acquisition, investigation, methodology, validation, writing–review and editing. R.A.: Conceptualization, funding acquisition, investigation, validation, writing–original draft, writing–review and editing.
Ethics Approval
This article does not contain data from any studies with human participants or animals which were performed by any of the authors.
Disclosure Statement
The authors declare that they have no conflicts of interest related to the studies reported here.
Funding Information
This work was supported by National Science and Technology Development Agency (Grant No. P-16-51223).