
Abstract
Pseudomonas aeruginosa biofilm formation has been considered to be an important determinant of its pathogenicity in most infections. The antibiofilm activity of trans-cinnamaldehyde (TC) against P. aeruginosa was investigated in this study. Results demonstrated that the minimum inhibitory concentration (MIC) of TC against P. aeruginosa was 0.8 mg/mL, and subinhibitory concentrations (SICs) was 0.2 mg/mL and below. Crystal violet staining showed that TC at 0.05–0.2 mg/mL reduced biofilm biomass in 48 h in a concentration-dependent mode. The formation area of TC-treated biofilms was significantly declined (p < 0.01) on the glass slides observed by light microscopy. Field-emission scanning electron microscopy further demonstrated that TC destroyed the biofilm morphology and structure. Confocal laser scanning microscopic observed the dispersion of biofilms and the reduction of exopolysaccharides after TC treatment stained with concanavalin A (Con-A)-fluorescein isothiocyanate conjugate and Hoechst 33258. Meanwhile, TC caused a significant decrease (p < 0.01) in the component of polysaccharides, proteins, and DNA in extracellular polymeric substance. The swimming and swarming motility and quorum sensing of P. aeruginosa was also found to be significantly inhibited (p < 0.01) by TC at SICs. Furthermore, SICs of TC repressed the several genes transcription associated with biofilm formation as determined by real-time quantitative polymerase chain reaction. Overall, our findings suggest that TC could be applied as natural and safe antibiofilm agent to inhibit the biofilm formation of P. aeruginosa.
Introduction
P
Bacterial biofilms are large cellular communities encased in an extracellular matrix containing cells in various physiological and morphological states and are the primary form in which most bacteria exist (Chou et al, 2022; Sadiq et al, 2021). P. aeruginosa is a common producer of biofilms, leading to persistent bacterial infections by providing protection against antibiotics, other antimicrobial agents, and the host immune defense system (Chang et al, 2022). The biofilm of P. aeruginosa is the leading cause of infection in patients with ventilator-associated pneumonia, with a mortality rate of 35.8% (Bonell et al, 2019). Biofilm formed by P. aeruginosa infection colonization hinders human wound healing (Stuermer et al, 2021). In addition, in the food industry, P. aeruginosa can bind to various surfaces of food and industrial equipment and form biofilms, posing a significant potential hazard (Balabanova et al, 2020; Chegini et al, 2020).
The biofilm formation process is relatively complicated, involving complex genetic networks and communication and cooperation between bacteria (Mevo et al, 2021). It often concerns many factors, for example, quorum-sensing (QS), proteases, the motility of bacterial cells, the attachment to biotic or abiotic surfaces by adhesive proteins, the secretion of extracellular polymeric substances (EPSs) composed of polysaccharides, DNA, proteins, and other global regulators (Zhang et al, 2020). These behaviors occur in different stages of biofilm formation. Therefore, by intervening in these activities, it is a strategy to influence biofilm formation.
Previous studies have shown that natural active substances can inhibit the bacterial virulence factor by reducing its expression, thus weakening the continuous increase of bacterial resistance (Silva et al, 2016). Sun et al (2017) demonstrated that the naturally occurring flavonoid hypericin inhibits biofilm formation in P. aeruginosa. Recent studies have suggested that soy isoflavones inhibit P. aeruginosa biofilm formation and QS (Yin et al, 2022). Trans-cinnamaldehyde (TC) is a natural compound separated from cinnamon essential oil; it is an unsaturated aldehyde, which is mainly used to add cinnamon fragrance to medical products, cosmetics, and perfumes (Shen et al, 2015).
It has been recognized as a Food and Drug Administration (FDA)-approved food safety ingredient due to its fast metabolism and less residue in the human body, and has been widely used in food flavoring, improving microbial safety and prolonging shelf life (Zhou et al, 2020). In recent years, studies have shown that TC has antimicrobial and antibiofilm activities against a wide range of bacterial pathogens, including Escherichia coli (Amalaradjou et al, 2010), Staphylococcus aureus (Kot et al, 2020), Staphylococcus epidermidis (Albano et al, 2019), and Streptococcus mutans (He et al, 2019).
It has a wide range of applications due to its broad spectrum of antibacterial, antiseptic, and antioxidant activities (Chen et al, 2017). Although the antimicrobial characteristics of TC have been extensively investigated, little research has been directed toward its antibiofilm activities against P. aeruginosa. Topa et al (2018) explored the antibacterial effect of TC on P. aeruginosa, but only focused on the effect of TC on the modulation of intracellular c-di-GMP levels and swarming motility, did not further explore the effect of TC on EPS production, QS, and gene expression associated with biofilms.
Thus, the study was aimed to investigate the effect of TC on the biofilm formation ability of P. aeruginosa, including the effects of TC on biofilm biomass, EPS production, bacterial motility, QS, and expressions of biofilm-associated genes, and the possible molecular mechanism of its antibiofilm activity was further elucidated.
Materials and Methods
Reagents
TC (HPLC ≥99%, CAS 14371-10-9) was purchased from Sigma-Aldrich Trading Co. (Shanghai, China) and was dissolved in dimethyl sulfoxide (DMSO; ST038-500 mL, Beyotime Biotechnology, Shanghai, China). The final concentration of DMSO in all sample solutions (treatment and control samples) was 1% (v/v), with no significant effect on the P. aeruginosa growth (Guo et al, 2016). Tryptic soy agar (TSA) and tryptic soy broth (TSB) were purchased from Land Bridge Technology Co. (Beijing, China). All other chemicals and reagents were analytically pure.
Bacterial strains and growth conditions
P. aeruginosa PAO1 strain was obtained from the American Type Culture Collection (Manassas, VA). The strain was stored at −80°C in TSB containing 25% (v/v) glycerol. P. aeruginosa was plated on TSA and incubated at 37°C for 24 h before each experiment. A single colony was then inoculated into TSB and shaken at 130 rpm for 12 h at 37°C. The resulting culture was centrifuged (8000 × g, 5 min, 4°C). The cell suspensions were washed twice with sterile phosphate-buffered saline (PBS; pH 7.4) and was diluted with PBS to produce a cell concentrate with an optical density at 600 nm (OD600nm) of 0.5 (∼4 × 108 colony forming units [CFU]/mL).
Determination of minimum inhibitory concentration and subinhibitory concentrations
The minimum inhibitory concentration (MIC) of TC against P. aeruginosa was determined according to the method recommended by the American Institute of Clinical and Laboratory Standards, with slight modifications (CLSI, 2021). The strain was diluted to 5 × 105 CFU/mL in TSB. One hundred microliters of bacterial solution with 100 μL of TC diluted with TSB were mixed in 96-well plates to achieve final TC concentrations of 0 (control), 0.1, 0.2, 0.4, 0.8, and 1.6 mg/mL. TSB supplemented with 1% DMSO was used as the blank control. The samples were incubated at 37°C for 24 h and determined the OD600nm values with a microplate reader (Model 680; BioRad Laboratories, Hercules, CA). The MIC was defined as the lowest antimicrobial concentration of TC corresponding to an OD600nm change of <0.05.
Then the subinhibitory concentrations (SICs) of TC against P. aeruginosa were determined using a fully automated Bioscreen C Plate Reader (Labsystems, Helsinki, Finland), which was conducted according to Fan et al (2022b) with slight modification. In brief, P. aeruginosa was diluted with TSB to 2 × 106 CFU/mL. One hundred twenty-five microliters of bacterial suspension was mixed with equal volume of TC with the concentrations of 0.0125–0.8 mg/mL, and then transferred to a 100-well plate. The plate was incubated in automatic Bioscreen plate reader (Labsystems) at 37°C for 24 h with OD600nm measurements of the cultures every 1 h. The obtained value was used to generate a growth curve. TC concentrations that did not inhibit the growth of P. aeruginosa were used as SICs.
Effect of TC on the total amount of P. aeruginosa biofilm
Specific biofilm formation inhibition assay
Crystal violet (CV) staining method was used to evaluate the antibiofilm formation ability with different concentrations of TC (0 [control], 0.05, 0.1, 0.2, and 0.4 mg/mL) as described by Kang et al (2018). A suspension of ∼108 CFU/mL P. aeruginosa and TC was mixed evenly and added into 96-well polystyrene plates with 200 μL per well, cultures were then incubated at 25°C and 37°C for 48 h, respectively, to form biofilms. At the end of the period, measurements of OD630nm were collected. Each well was then gently washed three times with sterile PBS to clean the biofilms. Biofilms were stained with 0.1% (w/v) CV for 20 min and cleaned with sterile distilled water. After the wells were dried, biofilms were stained with 33% (v/v) glacial acetic acid for 20 min and the absorbance was read at 570 nm using a microplate reader (Model 680; Bio-Rad Laboratories, Hercules, CA). The specific biofilm formation (SBF) capacity of the strain was assessed using the following formula:
Optical microscope-based observation
The inhibitory effect of TC on biofilm formation was assessed by visual observation of biofilm surface morphology using an optical microscope (XTZ-D; Shanghai Optical Instrument Factory, Shanghai, China) following the method described by Li et al (2021), slightly modified. Two milliliters of P. aeruginosa suspension (∼8 log CFU/mL) supplemented with TC (0 [control], 0.05, 0.1, 0.2, and 0.4 mg/mL) was seeded on sterile microscope cover glass (ϕ 1 cm) in 24-well polystyrene plates. Cultures were incubated at 25°C and 37°C for 48 h to allow biofilms to form on the surfaces of the glass coverslips. The slides containing biofilm were gently washed and stained with 0.4% (w/v) CV for 20 min. After staining, the glass coverslips were washed gently with distilled water, removed excess stain and dried. An optical microscope at 400 × magnification was used to observe the biofilm samples. Then percent biofilm coverage per unit area was measured by ImageJ software.
FESEM observation
The effect of TC on the morphology of P. aeruginosa biofilm was evaluated by field emission scanning electron microscopy (FESEM) conducted by a previous method with some modifications (Lee et al, 2020). P. aeruginosa biofilms were formed on the surface of sterile stainless steel coupons (304, 1 × 1 cm, finish 4) in a 24-well microtiter plate. Each well was filled with 2 mL bacterial suspension [∼108 CFU/mL]. A series of concentrations of TC (0 [control], 0.05, 0.1, 0.2, and 0.4 mg/mL) was added to the wells and incubated at 37°C for 48 h. Then the coupons were incubated at 4°C overnight in PBS containing 2.5% (v/v) glutaraldehyde to fix the biofilms, washed with PBS and continuously dehydrated with ethanol (30%, 50%, 70%, 80%, 90%, and 100%). Sputter-coated with gold under a vacuum, prepared biofilms were examined using FESEM (S-4800; Hitachi, Tokyo, Japan; 1500 × and 4000 × magnification).
Confocal laser scanning microscopy observation
The effects of TC on biofilm polysaccharides and structure were determined by confocal laser scanning microscopy (CLSM) as described previously by Peng et al (2018). P. aeruginosa biofilms on coupons were formed using the method described in Optical Microscope-Based Observation section, then biofilms were stained with concanavalin A (Con-A; Invitrogen/Molecular Probes, Eugene, OR)-fluorescein isothiocyanate (FITC) conjugate in the dark for 15 min at 4°C. Thereafter, they were immobilized with 2.5% (v/v) glutaraldehyde and stored at 4°C for 2 h and stained with Hoechst 33258 (Solarbio, Beijing, China) for 20 min at room temperature. Finally, the samples were washed with PBS and observed using a CLSM (CLSM; A1; Nikon, Tokyo, Japan) at 40 × magnification. Bacterial cells were stained blue by Hoechst 33258, biofilm polysaccharides were dyed green by Con-A. FITC and Hoechst 33258 were imaged with 488 and 405 nm excitation laser beams, respectively, and images were processed using NIS-Elements Viewer 4.20 software.
Effect of TC on EPS of P. aeruginosa
Components of polysaccharide, protein, and DNA in biofilm were measured by previously described method (Kang et al, 2018). P. aeruginosa biofilms were formed on stainless steel coupons in 24-well microtiter plates with the TC treatments (0 [control], 0.05, 0.1, 0.2, and 0.4 mg/mL) and incubated at 37°C for 48 h as described in the previous section. Then biofilms were cleaned with PBS to remove planktonic bacteria. Samples were placed in centrifuge tubes containing 0.2 g of glass beads and 1.5 mL of sterile PBS and vortexed for 3 min.
The biofilms were disrupted using ultrasound dispersal for 15 min and the mixtures centrifuged at 4000 rpm for 30 min and the supernatant was extracted to harvest EPS components. The content of extracellular polysaccharides was determined by the anthrone-H2SO4 reagent. The extracellular protein content was detected with a BCA protein assay kit (BB-3130; BestBio, Shanghai, China) following the manufacturer's instructions. And the extracellular DNA (eDNA) was measured by OD260nm spectrophotometry measurements.
Quantitative QS inhibition assay
The effect of TC on QS inhibition assay of P. aeruginosa was on a method described by Shi et al (2017). The QS inhibitory activity was measured by quantifying the violacein production with the indicator strain Chromobacterium violaceum ATCC 12472. The four highest concentrations of TC were selected as SICs for this study using the method described in Determination of MIC and SICs section, which did not inhibit the growth of C. violaceum after 24 h incubation. C. violaceum was diluted to OD600 nm = 0.2, cultured overnight, and added to TSB containing TC (0 [control], 0.00625, 0.0125, 0.025, and 0.05 mg/mL). Then the samples were incubated at 30°C for 24 h with 130 rpm shaking.
Three milliliters of culture was centrifuged (5000 × g, 5 min, 4°C) to precipitate the insoluble violacein and the liquid supernatant was discarded. Thereafter, 1 mL of DMSO was added to the centrifuge tube and the solution was vortexed to completely dissolve violacein. The violacein-containing supernatants were obtained after centrifugation (10,000 × g, 10 min, 4°C). Finally, the supernatant (200 μL) was added to 96-well microtitration plates and the optical absorbance was measured at 585 nm using a microplate reader (Model 680; Bio-Rad, Hercules, CA) (Choo et al, 2006).
Swimming and swarming motility assay
The motility assays were carried out using the method of Bai et al (2019) with minor modifications. For swimming assay, 5 μL of overnight P. aeruginosa culture (OD600 nm = 0.5) was spotted on the center of the swimming agar panel composed of 0.3% (w/v) agar and TSB containing TC (0 [control], 0.05, 0.1, 0.2, and 0.4 mg/mL). For analysis of bacterial swarming ability, TSB containing 0.5% (w/v) agar and 0.5% (w/v) glucose was used for swarming agar. Different concentrations of TC (0 [control], 0.05, 0.1, 0.2, and 0.4 mg/mL) were added to the medium and 5 μL of overnight culture was inoculated at the center of medium surface. Then the plates were incubated at 37°C for 48 h. The images of bacterial movement were collected by Gel Imaging System (Bio-Rad), and the area of the motility was measured by ImageJ software.
RNA extraction and real-time quantitative polymerase chain reaction
To explore the effects of TC on the expression of biofilm-related genes, P. aeruginosa PAO1 suspension was added to TSB with different concentrations (0 [control], 0.2, and 0.4 mg/mL) of TC and cultured at 37°C for 10 h. RNA isolation, cDNA synthesis, and real-time quantitative polymerase chain reaction (RT-qPCR) were performed using the method described by Liu et al (2022). According to the manufacturer's instructions, total RNA of the samples was extracted using RNAprep pure bacteria kit (Tiangen, Beijing, China). The RNA concentrations were subsequently quantified using a nucleic acid and protein spectrophotometer (Nano-200; Aosheng Instrument Co., Ltd., Hangzhou, China).
cDNA was then synthesized using the PrimeScript RT Master Mix (Takara, Kyoto, Japan). The primers used in the study are presented in Table 1. RT-PCR was performed in a 25-μL volumes using SYBR Premix Ex Taq II (Takara) under the following reaction conditions: initial denaturation at 95°C for 30 s, followed by 40 cycles of 95°C for 5 s, 60°C for 30 s, and a dissociation step of 95°C for 15 s and 60°C for 30 s. The relative gene expression levels of target genes were analyzed by the 2−ΔΔCt method and the rpoD gene was used as the internal standard to normalize the expression of genes (Yin et al, 2011).
Primer Sequences for Real-Time Quantitative Polymerase Chain Reaction
Statistical analysis
Statistical analysis was performed using SPSS software (version 20.0; IBM Corp, Armonk, NY). All experiments were conducted in triplicate. Data were expressed as mean values ± standard deviation. Significance was expressed as *p < 0.05 and **p < 0.01 versus the control; # p < 0.05 and ## p < 0.01 for comparison between TC treatments; and ns, no significant difference. Significant differences between experimental groups were determined by ANOVA with LSD test.
Results
Minimum inhibitory concentrations
TC showed antimicrobial effect against P. aeruginosa PAO1. The MIC value of TC toward P. aeruginosa PAO1 was 0.8 mg/mL.
Subinhibitory concentrations
Figure 1 showed the effect of TC on the growth of P. aeruginosa PAO1 at 0.0125 to 0.8 mg/mL. The obtained results showed that TC at 0.8 mg/mL could inhibit the growth of P. aeruginosa PAO1. And we found that the growth lag period of P. aeruginosa was prolonged by the treatment with 0.4 mg/mL of TC. Therefore, TC at 0.05, 0.1, and 0.2 mg/mL was selected as the SICs against P. aeruginosa in this study.

Growth curve analysis of TC against Pseudomonas aeruginosa PAO1 in TSB. Bars represent the standard deviation (n = 3). OD, optical density; TC, trans-cinnamaldehyde; TSB, tryptic soy broth.
Specific biofilm formation
The effect of different concentrations of TC on P. aeruginosa PAO1 biofilm formation, as quantified by CV, is shown in Figure 2. The quantitative results showed that the formation of biofilm of P. aeruginosa gradually decreased in a concentration-dependent manner under incubation conditions for 48 h at 25°C and 37°C. With the increase of concentrations of TC from 0.05 to 0.4 mg/mL, the SBF of P. aeruginosa was decreased to 1.29–0.84 and 1.99–1.42 after incubation at 25°C and 37°C, respectively (Fig. 2a, b). Staining of CV on 96-well microtiter plates showed that the color of CV gradually became lighter as the concentration of TC increased.
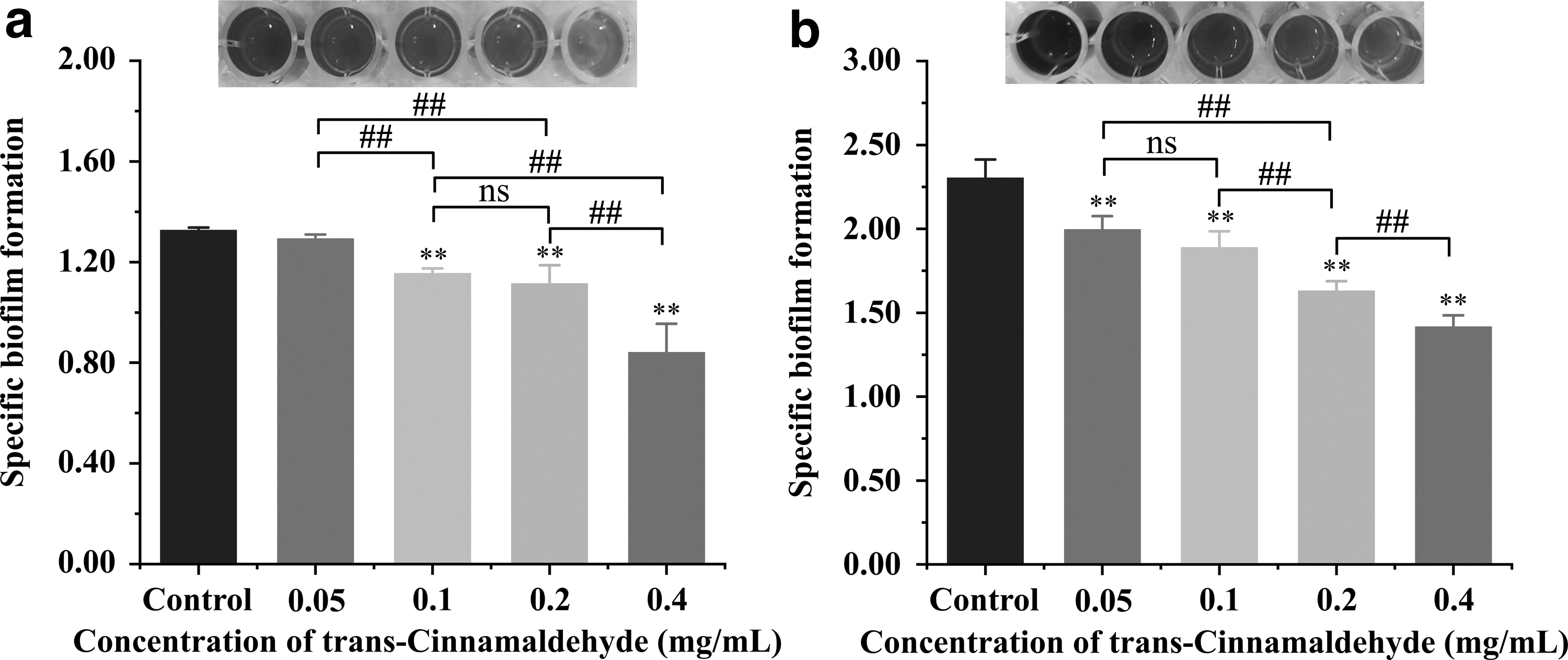
Inhibition of Pseudomonas aeruginosa PAO1 biofilm formation by TC at different concentrations with images were captured after staining with crystal violet at 25°C
Effects of TC on biofilm morphology
According to the images of light microscopy, the biofilm of the control group almost completely covered the surface of the coverslips, and was evenly distributed (Fig. 3a—A and F). In contrast, the surface coverage of P. aeruginosa on the glass slides was significantly declined (p < 0.01). The biofilm diffusion was increased and the adhesion decreased under the treatment of TC at 0.05 to 0.4 mg/mL (Fig. 3a—B–E and G–J). The biofilm coverage calculated from the light microscopy images was consistent with the specific trend of biofilm formation. The biofilm coverage of P. aeruginosa PAO1 decreased by 10.31%, 30.61%, 57.45%, and 91.64% after TC treatment at 0.05, 0.1, 0.2, and 0.4 mg/mL, respectively, under 37°C incubation, and by 12.77%, 49.47%, 85.49%, and 89.01% under 25°C conditions, respectively, compared with those in control (Fig. 3b, c).

Light microscope images
The effect of different concentrations of TC on the biofilm structures of P. aeruginosa was observed using FESEM under 1500 × and 4000 × magnification. Untreated P. aeruginosa were observed to form a dense, intact, and uniformly adherent biofilm on the surface of the steel sheets. Most cells adhered to each other and aggregated into a multilayered structure (Fig. 4A, F). With the increase of TC concentrations, the bacterial density was decreased and the structure gradually loosened. TC inhibited the development of P. aeruginosa biofilm structure. After 0.4 mg/mL of TC treatment, the typical structure of the biofilm was almost disappeared (Fig. 4B–E, G–J).
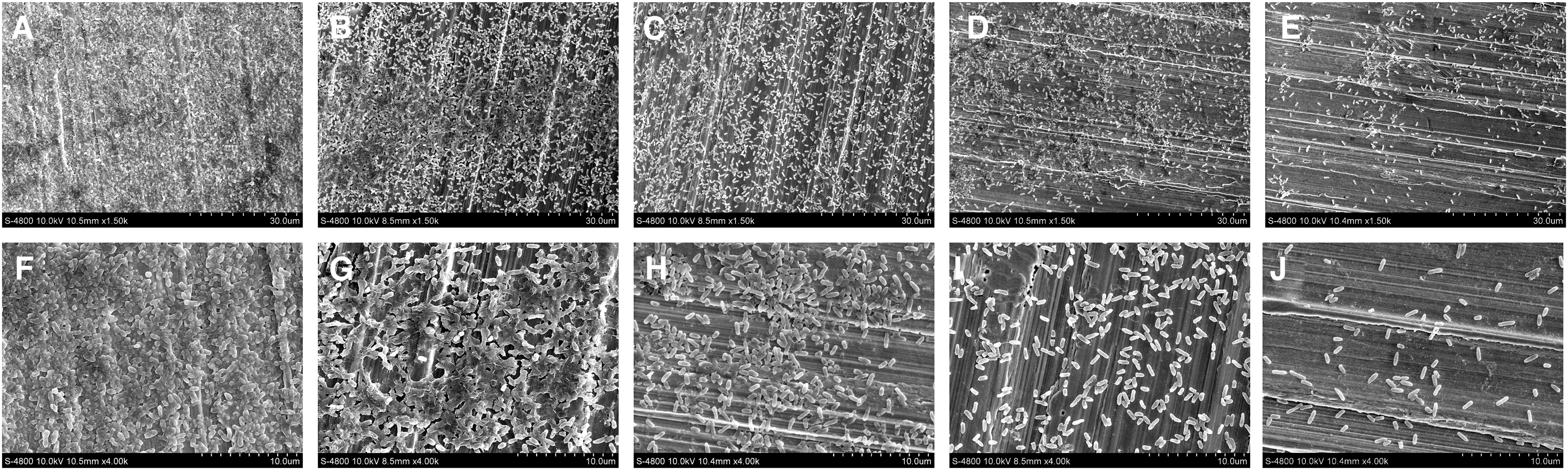
Effects of TC at 0
Effects of TC on polysaccharides, proteins, and DNA in biofilms
Changes in the content of the three main components of biofilms under the action of TC are shown in Figure 5. Compared with the control, TC treatment significantly reduced the contents of P. aeruginosa exopolysaccharides, extracellular proteins, and eDNA (p < 0.01). Under 0.05, 0.1, 0.2, and 0.4 mg/mL TC treatments, the extracellular polysaccharide content decreased to 0.084, 0.072, 0.038, and 0.030 mg/mL, respectively, decreasing by 20.00%, 31.43%, 63.81%, and 71.43% of the control level (Fig. 5a). Extracellular proteins content was significantly (p < 0.01) reduced by 17.46%, 22.82%, 59.82%, and 79.67% compared with that of the control after 0.05, 0.1, 0.2, and 0.4 mg/mL TC treatment, respectively (Fig. 5b). Furthermore, eDNA content has also been significantly affected, and the trend is similar to that of exopolysaccharides and proteins (Fig. 5c). In general, this inhibitory effect gradually increased as TC addition increased.

The contents of
CLSM analysis showed that untreated P. aeruginosa formed thick biofilms. Dense blue and green fluorescent regions could be observed. The bacteria were closely connected and the polysaccharides were evenly distributed throughout the biofilm (Fig. 6A1–A3). However, the blue and green fluorescence content decreased in the P. aeruginosa biofilm samples after TC treatment. This indicated that the bacteria were dispersed and the polysaccharide content was reduced (Fig. 6B1–B3, C1–C3, D1–D3, E1–E3).

Confocal laser scanning microscope images (40 × magnification) of Pseudomonas aeruginosa PAO1 biofilms with TC treatment at 0
Influence of TC on swimming and swarming motility
The motility of microorganisms plays a crucial role in the adhesion of surfaces and the subsequent formation of biofilms (Monte et al, 2014). It was found that TC significantly inhibited flagellate-mediated motility of P. aeruginosa PAO1 on agar plates in a dose-dependent way (p < 0.01), with the largest inhibition of bacterial migration observed in the 0.4 mg/mL treatment group (Fig. 7). As shown in Figure 7a and b, the swimming migration areas of cells treated with TC (0.05, 0.1, 0.2, and 0.4 mg/mL) were about 53.37%, 42.33%, 30.23%, and 21.89%, respectively, and the swarming areas were about 59.12%, 36.32%, 26.85, and 21.70%, respectively, of the control group.

The inhibition of TC on the motility of Pseudomonas aeruginosa PAO1.
Anti-QS activity of TC
To assess the effect of TC on P. aeruginosa QS indirectly, we quantified the production of violacein. In this study, TC at different concentrations (0. 00625–0.05 mg/mL) had no apparent antimicrobial activity on the growth of C. violaceum 12472 (data not shown). Figure 8 shows that TC significantly inhibited the violacein production of C. violaceum in an apparent dose-dependent manner (p < 0.01). Compared with the control group, the production of violacein decreased by 36.16%, 53.98%, 60.66%, and 74.10% following 0.00625, 0.0125, 0.025, and 0.05 mg/mL of TC, respectively.
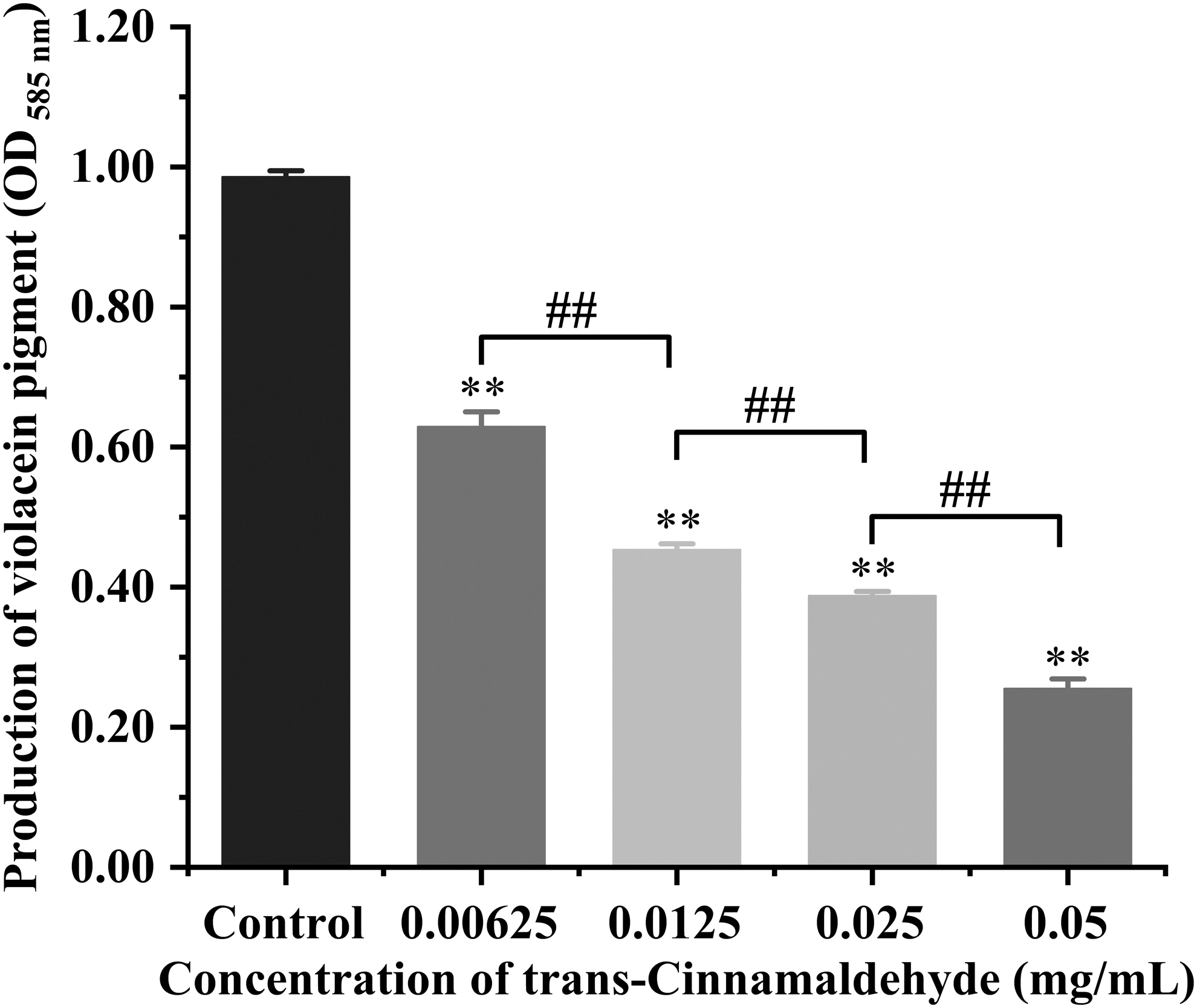
Inhibition of violacein production by Pseudomonas aeruginosa PAO1 at different concentrations of TC. Bars represent the means ± standard deviations (n = 3). **p < 0.01 when compared with control group; ## p < 0.01 for comparison between TC treatments.
Effect of TC on the gene expression levels related to biofilm formation of P. aeruginosa
QS, adherence, polysaccharide, and biofilm-related genes are involved in the processes of P. aeruginosa biofilms formation. RT-qPCR results (Fig. 9) demonstrated that TC at 0.2 mg/mL significantly (p < 0.01) reduced expression levels of lasR and rhlR (critical for QS), lecA (PA-I galactophilic lectin), oprM (outer membrane protein OprM), bdlA (biofilm dispersion protein), pslA (biofilm-formation protein PslA) genes, and pslB (biofilm-formation protein PslB) gene showed significant difference (p < 0.05) compared with the nontreated control (except lecB). Besides, 0.1 mg/mL concentrations of TC significantly (p < 0.01) downregulated rhlR gene expression and significantly (p < 0.05) suppressed lasR, oprM, and pslA genes transcription. However, TC had no significant effect on lecB (fucose-binding lectin PA-IIL) and pslB genes transcription (p > 0.05). RT-qPCR analysis showed that there was a different degree of transcription of P. aeruginosa biofilm-associated genes after varying concentrations of TC.

Effects of TC on the transcription of genes associated with the Pseudomonas aeruginosa PAO1 biofilms formation. Bars represent the standard deviation (n = 3). *p < 0.05, **p < 0.01 versus the control.
Discussion
Biofilm production produced by P. aeruginosa has been considered to be an important determinant of its pathogenicity in most infections (Shokri et al, 2018). P. aeruginosa causes chronic infections dominated by a biofilm lifestyle in some hospital settings, including medical equipment-related and pulmonary infections, and has the potential to harm the food industry through contaminated food (Taylor et al, 2014; Xu et al, 2019). Therefore, controlling its formation is an effective way to solve the related problems caused by biofilms. At present, antibiotics and disinfectants are widely used in clinical and industrial production to overcome biofilm contamination.
However, biofilms as a survival strategy prevalent among bacteria can promote bacteria to improve resistance to chemical disinfectants (Yu et al, 2020). Plant natural products can target the virulence of pathogens to control bacterial infections without disrupting bacterial growth or viability, thus having the potential to decrease the pressure for resistance (Silva et al, 2016). The biofilm formation of P. aeruginosa was measured at SICs (0.2, 0.1, and 0.05 mg/mL) of TC in this study. These concentrations were proved to have no significant inhibitory effects on the growth and survival of bacteria (Fig. 1), so as to ensure that subsequent antibiofilm formation experiments would not be caused by affecting bacterial growth.
In our study, the amount of biomass of P. aeruginosa was detected by CV staining (Fig. 2). TC inhibited the biofilm of P. aeruginosa at SICs at 25°C and 37°C in a concentration-dependent manner. Similarly, Nguyen et al (2021) reported that 24 μmol/L of alpha-mangostin-loaded nanoparticles decreased the biomass of S. aureus by 53–62%. Liu et al (2019) found that sulfated chitosan inhibited the amount of P. aeruginosa biofilm formation by >90% using the CV method. CV staining is a classical method for detecting biomass quantities.
At present, the application of natural active substances to the bacterial biofilm control on food-contact surfaces has been mainly investigated in terms of biofilm removal, biofilm formation inhibition, and biofilm diffusion prevention (Rossi et al, 2022). The CV staining method can be used for quantitative detection of bacterial biofilms. In this study, we found that TC could inhibit biofilm formation at SICs. This suggests that TC has the potential to be developed as a potent agent against P. aeruginosa biofilm formation in food industries.
The degree of dispersion and structure of P. aeruginosa were also shown to be reduced by TC (Figs. 3 and 4). After treatment with different concentrations of TC, the formation of P. aeruginosa biofilm was reduced and the adhesion capacity was decreased. Similarly, Bai et al (2019) observed that SICs of Shikimic acid significantly reduced the biofilm biomass of S. aureus and loosened the colonies. Zhang et al (2019) also visualized that SICs of hexanal reduced the amount of bacteria in the biofilm of Erwinia carotovora and Pseudomonas fluorescens.
LecA and LecB are soluble tetrameric lectins bound to galactose and fucose, respectively (Hu et al, 2017). They mediate the adhesive with host cells and the infection and biofilm formation of P. aeruginosa (Huang et al, 2018). BdlA is a chemotactic regulator essential for P. aeruginosa biofilm dispersion, and the lowering of bdlA expression level disrupts the biofilm dispersion response (Wang et al, 2019). The present RT-qPCR results demonstrated that SICs of TC downregulated the expression of lecA and bdlA without significant effects on lecB (Fig. 9). Therefore, TC may reduce the exertion of LecA and BdlA functions by downregulating the expression of lecA and bdlA, limiting the adhesion and dispersal ability of P. aeruginosa biofilm and thus inhibiting biofilm formation.
The EPS matrix consists of a mixture of polysaccharides, nucleic acids (e.g., eDNA), proteins, lipids, and other types of polymeric substrates. It is essential for the construction of three-dimensional biofilm architectures (Guo et al, 2021). In this study, the contents of polysaccharides, protein, and DNA in the biofilm declined (Fig. 5a–c). Nahar et al (2021) found that 800 μL/mL of flavourzyme could remove 94.3% and 90.0% of total protein and crude carbohydrate in the EPS of P. aeruginosa biofilm on rubber surfaces, respectively. Liu et al (2022) found that proteins, nucleic acids, and carbohydrates in EPS components of Vibrio parahaemolyticus were reduced after sodium butyrate treatment by the laser micro-Raman spectrometer.
Furthermore, CLSM was used to observe the dispersion of biofilms and the reduction of polysaccharides after TC treatment (Fig. 6). Similarly, Yang et al (2020) also found that the complete biofilm of P. aeruginosa was dispersed after 2 h LED irradiation at 4°C, 10°C, and 25°C, respectively, and the abundance of polysaccharides decreased significantly. Li et al (2022) observed by CLSM that cells and polysaccharides in S. aureus and P. aeruginosa biofilms were degraded into fragments after didecyldimethylammonium bromide combined with slightly acidic electrolyzed water treatment.
This study has shown that the mannose rich psl polysaccharides encoded by the psl cluster was required for formation of P. aeruginosa biofilms (Yang et al, 2011). The gene pslA and pslB are located in a 15-gene (pslA to pslO) psl operon required for exopolysaccharide synthesis and biofilm formation (Lee et al, 2008). Our RT-qPCR results demonstrated that the transcription of pslA and pslB were downregulated after TC treatment (Fig. 9). Therefore, these results indicated that TC inhibited the formation of P. aeruginosa biofilm by reducing the synthesis of polysaccharides and other components in EPS.
The QS signaling system produces several types of N-acyl-
Fan et al (2022a) reported that SICs of lactobionic acid inhibited the motility of Salmonella. The P. aeruginosa genome encodes two population-sensing transcription factors: lasR and rhlR (Parsek and Greenberg, 2000). lasR activates rhlR transcription, and the expression of rhlR-regulated target genes is downregulated by the absence of lasR (Wagner et al, 2003). We discovered that TC downmodulated the expression of lasR and rhlR (Fig. 9). Figure 8 shows that SICs of TC inhibited the QS signal resulting in a significant reduction in the production of violetin in C. violaceum. It is hypothesized that TC downregulates the expression of lasR, which in turn inhibits the exertion of rhlR, interfering with bacterial intercellular communication to inhibit QS and thus the motility of P. aeruginosa.
Conclusion
This study investigated the inhibitory effects of TC against P. aeruginosa biofilm formation. The findings demonstrated that TC could decrease the biofilm biomass of P. aeruginosa on stainless steel coupons and glass slides surfaces, collapsing the biofilm architecture. TC treatment also caused the lower EPS production, reduced polysaccharides, proteins, and bacteria content in biofilms. Moreover, TC also inhibited the swimming and swarming motility and QS of P. aeruginosa, downregulated the transcription of genes involved in biofilm-associated and QS. Our results provided an improved understanding of TC as a biofilm inhibitor against P. aeruginosa. This study provides a theoretical basis for the application of TC to control P. aeruginosa biofilm in food systems and other environments, and to mitigate foodborne disease caused by this pathogen.
Footnotes
Authors' Contributions
Conceptualization, investigation, formal analysis, project administration, writing—original draft, and writing—review and editing by L.S. Resources, investigation, data curation, and visualization by H.Y. Resources, data curation, and investigation by X.M. Data curation, resources, and methodology by R.S. Visualization, formal analysis, and resources by S.C. Methodology, supervision, and software by H.W. Visualization, formal analysis, and validation by X.B. and D.G. Methodology, resources, and supervision by X.L. Supervision and project administration by X.X. Project administration, supervision, and funding acquisition by C.S. All authors have read and agreed to the published version of the article.
Availability of Data and Materials
The data that support the findings of this study are available from the corresponding author on reasonable request.
Ethics Approval
This study does not present experiments with humans or animals.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (31801659), the class General Financial Grant from the Shaanxi Postdoctoral Science Foundation (2018BSHEDZZ150), and the Shaanxi Key Research and Development Program (2019NY-118).