
Abstract
Staphylococcus aureus is a major foodborne pathogen that leads to various diseases due to its biofilm and virulence factors. This study aimed to investigate the inhibitory effect of 2R,3R-dihydromyricetin (DMY), a natural flavonoid compound, on the biofilm formation and virulence of S. aureus, and to explore the mode of action using transcriptomic and proteomic analyses. Microscopic observation revealed that DMY could remarkably inhibit the biofilm formation by S. aureus, leading to a collapse on the biofilm architecture and a decrease in viability of biofilm cell. Moreover, the hemolytic activity of S. aureus was reduced to 32.7% after treatment with subinhibitory concentration of DMY (p < 0.01). Bioinformation analysis based on RNA-sequencing and proteomic profiling revealed that DMY induced 262 differentially expressed genes and 669 differentially expressed proteins (p < 0.05). Many downregulated genes and proteins related to surface proteins were involved in biofilm formation, including clumping factor A (ClfA), iron-regulated surface determinants (IsdA, IsdB, and IsdC), fibrinogen-binding proteins (FnbA, FnbB), and serine protease. Meanwhile, DMY regulated a wide range of genes and proteins enriched in bacterial pathogenesis, cell envelope, amino acid metabolism, purine and pyrimidine metabolism, and pyruvate metabolism. These findings suggest that DMY targets S. aureus through multifarious mechanisms, and especially prompt that interference of surface proteins in cell envelope would lead to attenuation of biofilm and virulence.
Introduction
Food safety has been widely concerned in the field of public health. Staphylococcus aureus is the main pathogenic bacterium causing food poisoning, which can attach to various surfaces of food matrixes by building biofilm structures, leading to a serious risk of food contaminations and even foodborne outbreaks (Guo et al., 2021). Meanwhile, S. aureus can cause a wide variety of diseases, including skin infections, pneumonia, and endocarditis, due to its virulence and antibiotic resistance (Rozemeijer et al., 2015). As a consequence, the bacterial biofilm and virulence factors produced by S. aureus, especially staphylococcal enterotoxins and hemolysins, play a significant role in contributing to its pathogenesis. Therefore, much efforts have been devoted to exploring not only biofilm inhibition but also virulence factor release inhibition strategies.
Traditional chemical and physical strategies could produce the emergence of resistance, high energy consumption, and adverse effects on human health and sensory properties of food products (Simoes et al., 2010). To avoid these negative effects, a lot of efforts have been focused on natural antibacterial and antibiofilm agents. It was reported that resveratrol, a natural phenolic compound, can achieve a bacteriostatic effect by inhibiting biofilm formation of avian pathogenic Escherichia coli (Ruan et al., 2021). Moreover, various plant extracts rich in phenols and flavonoids have been demonstrated to possess a potential antibiofilm activity against S. aureus (de Oliveira et al., 2020; Dukanovic et al., 2020).
According to our previous study, the flavonoid compound isolated from pine needles of Cedrus deodara (Roxb.) Loud. (family: Pinaceae), 2R,3R-Dihydromyricetin (DMY), exerted excellent antibacterial and antibiofilm properties against S. aureus (Wu et al., 2018). Specifically, DMY could remarkably inhibit S. aureus cells adhering to food contact surfaces, such as stainless steel, thus showing a potential in controlling the persistence resuscitation and cross contamination of S. aureus in food industry. Although the mode of action of DMY against S. aureus was explored at cellular level (Wu et al., 2017), mechanisms driving its antibacterial and antibiofilm properties at molecular level are not clear.
The rapid enhancement of “omics” technologies has provided massive amounts of information on bacterial physiology, facilitating the exploration of bacterial targets for new antibiofilm agents. Using transcriptome and proteome analysis, it is speculated that plantaricin prevented the biofilm formation of S. aureus by inhibiting the production of surface matrix-associated proteins and the functions of regulatory factors (Du et al., 2020). In addition, proteomics showed that glabridin regulated the expression of S. aureus surface proteins and inhibited the adhesion of S. aureus (Gangwar et al., 2020). In this study, to explore the molecular targets of DMY against S. aureus, the alterations in gene and protein expression of bacterial cell were investigated using the incorporation of transcriptomic and proteomic technologies. This is the first report of DMY as a potent biofilm-targeting compound against S. aureus, which can effectively inhibit bacterial virulence, providing evidence that DMY exerts antibacterial activity through multiple pathways.
Materials and Methods
Experimental materials and culture medium
DMY (high-performance liquid chromatograph purity ≥98%) was purchased from Shanghai Yuanye Biotechnology Co. Ltd. (Shanghai, China). S. aureus ATCC 29213 strain was acquired from the China Medical Culture Collection Center (Beijing, China). Tryptone soy broth (TSB), nutrient broth, and glucose were purchased from Beijing Aoboxing Bio-Technology Co. Ltd. (Beijing, China). Dimethyl sulfoxide (DMSO) was obtained from Aladdin Reagent Co. (Shanghai, China). A stock solution of DMY was prepared in DMSO and diluted with corresponding vehicle before use to keep the final concentration of DMSO in bacterial suspension below 0.05%. The minimum inhibitory concentration (MIC) of DMY against S. aureus ATCC 29213 was 0.125 mg/mL (Wu et al., 2018).
Confocal laser scanning microscopic analysis
Confocal laser scanning microscope (CLSM) coupled with LIVE/DEAD BacLight Bacterial Viability Kit (Thermo Fisher Scientific, MA) were used to explore the biofilm microstructure changes induced by DMY (Bai et al., 2019). The biofilm of S. aureus in a 24-well microplate was prepared according to the previous method with some modifications (Dosler and Mataraci, 2013). In brief, logarithmic phase S. aureus cells were prepared in TSB containing 1% glucose at a cell density of 1 × 106 CFU/mL.
Aliquots of 500 μL of TSB containing DMY and 500 μL of the S. aureus suspension were added to each well of a microplate containing a sterile round glass slide (Φ 14 mm), giving the final concentration of DMY at 1/8 × MIC, 1/4 × MIC, and 1/2 × MIC. The S. aureus culture containing 0.05% DMSO without DMY was used as control. After incubating at 37°C for 24 h without shaking, the 1-d-old biofilm was formed. Then, 0.01 M phosphate-buffered saline (PBS, pH 7.2) was used for washing the glass slides twice. The biofilm was stained by a mixed dye solution of SYTO 9 and propidium iodide at final concentrations of 5 and 30 μM, respectively. After staining at 25°C for 15 min in the dark, the biofilm was observed using a CLSM (SP8; Leica, Germany).
Hemolysis assay
The hemolytic activity of S. aureus was evaluated as previously reported with some modifications (Shi et al., 2015). The logarithmic phase S. aureus cells were diluted with nutrient broth to 5 × 106 CFU/mL. Then, DMY was added to the bacterial suspension to obtain a final concentration of 1/8 × MIC, 1/4 × MIC, and 1/2 × MIC. After incubation for 12 h at 37°C, the bacterial culture was centrifugated at 1000 g and 4°C for 5 min. Aliquots of 100 μL of the obtained supernatant was added to 875 μL of 0.01 M PBS (pH 7.2), followed by addition of 250 μL of 4% defibrinated rabbit blood (Henan Shengkang Biotechnology Co. Ltd., Zhengzhou, China).
The mixture was incubated at 37°C for 30 min. After centrifugation, the optical density at 543 nm of the supernatant was determined. The S. aureus culture supernatant containing PBS without DMY was set as the 100% hemolysis control, while the supernatant incubated with 0.05% DMSO was tested as a vehicle control. The percent hemolysis of the samples was calculated by comparing with the 100% hemolysis control.
Treatment for transcriptomic and proteomic assays
DMY was added to logarithmic phase S. aureus cells suspension (1 × 107 CFU/mL) to obtain a final concentration of 1/2 × MIC. After incubation for 6 h at 37°C, the pellets were collected and washed twice with PBS, and then stored in liquid nitrogen for RNA and protein extraction. S. aureus culture containing 0.05% DMSO without DMY was set as the control. Each group was in triplicate.
Transcriptomic analysis
The total RNA of S. aureus was extracted with a TRIzol reagent (Invitrogen, Carlsbad, CA). The RNA purity and integrity assessment was performed on NanoPhotometer® spectrophotometer (IMPLEN) and Bioanalyzer 2100 system (Agilent). The construction of cDNA library and sequencing were conducted by Zhongke New Life Biotechnology Co. Ltd. (Shanghai, China). In brief, the first strand of cDNA was synthesized using a reverse transcriptase and 6-base random primers, and the second strand was synthesized using DNA Polymerase I and RNase H with the first strand as a template. The synthesized cDNA was purified on AMPure XP system and enriched by polymerase chain reaction (PCR). The resultant libraries were sequenced on an Illumina HiSeq4000 platform to generate 2 × 150 bp paired-end reads. The raw RNA sequence data were deposited in the National Center for Biotechnology Information (NCBI; PRJNA889563 and PRJNA890324).
Clean reads were obtained by removing reads containing adapter, ploy-N, and low-quality reads. Then, clean reads were mapped against S. aureus subsp. aureus NCTC 8325 genome using Hisat2 (v2.0.5). DESeq2 R package (1.16.1) was used for differential expression analysis between the two groups. Genes with padj <0.05 and fold change >2 or <0.5 were assigned as differentially expressed. Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis were used based on Fisher's exact test. GO terms and KEGG terms with p < 0.05 were considered to be significantly enriched.
Proteomic analysis
Bacterial cell pellets were added with SDT buffer (4% (w/v) sodium dodecyl sulfate, 1 mM dithiothreitol (DTT), and 100 mM Tris/HCl, pH 7.6) for protein extraction. After quantified by a bicinchoninic acid assay protein assay kit (Bio-Rad), the proteins were digested according to the filter-aided sample preparation procedure (Lin et al., 2017). The digested peptides were labeled with tandem mass tag reagent, and then fractionated by high pH reversed-phase peptide fractionation kit (Thermo Scientific). Liquid chromatography tandem mass spectrometry analysis was performed on a Q Exactive mass spectrometer coupled to an EASY-nLC system (Thermo Fisher Scientific). The mass spectrometry raw data were identified and quantitatively analyzed with Proteome Discoverer 1.4. Peptides with false discovery rate ≤0.01 were regarded as the identified protein.
Differentially expressed proteins (DEPs) with fold change >1.2 or <0.83 and p < 0.05 were conducted to bioinformation analysis. GO and KEGG enrichment analyses for DEPs were performed on Blast2GO and KEGG Automatic Annotation Server, respectively. Functional categories and pathways of p < 0.05 were set as significant.
Real-time-quantitative PCR verification
The expression levels of four randomly selected genes (gene-SAOUHSC_01110 encoding fibrinogen-binding protein, gene-SAOUHSC_02430 encoding iron citrate ATP-binding cassette (ABC) transporter substrate-binding protein, gene-SAOUHSC_02011 encoding regulatory protein recX, and gene-SAOUHSC_03037 encoding ABC transporter permease) were determined by real-time (RT)-quantitative PCR (qPCR) using ChamQ SYBR Color qPCR Master Mix (2 × ; Nanjing Novezan Biotechnology Co., LTD). The gene-specific primers used in the experiment are listed in Supplementary Table S1, and the 16S rRNA was set as an internal standard. The polymerase activation was carried out at 95°C for 5 min, followed by melting 95°C for 5 s, annealing 55°C for 30 s, and extending 72°C for 40 s for 40 cycles on a fluorescence quantitative PCR instrument (ABI 7300; Applied Biosystems).
Statistical analysis
All experiments were conducted in triplicate. Statistical analyses were performed on SPSS software (version 20.0; SPSS, I., Chicago, IL). Data differences with p < 0.05 were considered statistically significant.
Results
DMY damaged S. aureus biofilm
S. aureus cultured without DMY formed dense and uneven biofilms and the bacterial cells tightly adhered to each other (Fig. 1A). As suggested by Triveni et al. (2018) based on OD570nm value with crystal violet staining in microtiter plate method, the biofilm formation by S. aureus in this assay (OD570nm > 0.3) was strong. However, with the increasing of DMY concentration, the dispersibility of biofilms formed by S. aureus showed an increasing tendency of dose (Fig. 1B–D). When treated with 1/2 × MIC of DMY, single cells and intercellular spaces were clearly identified.
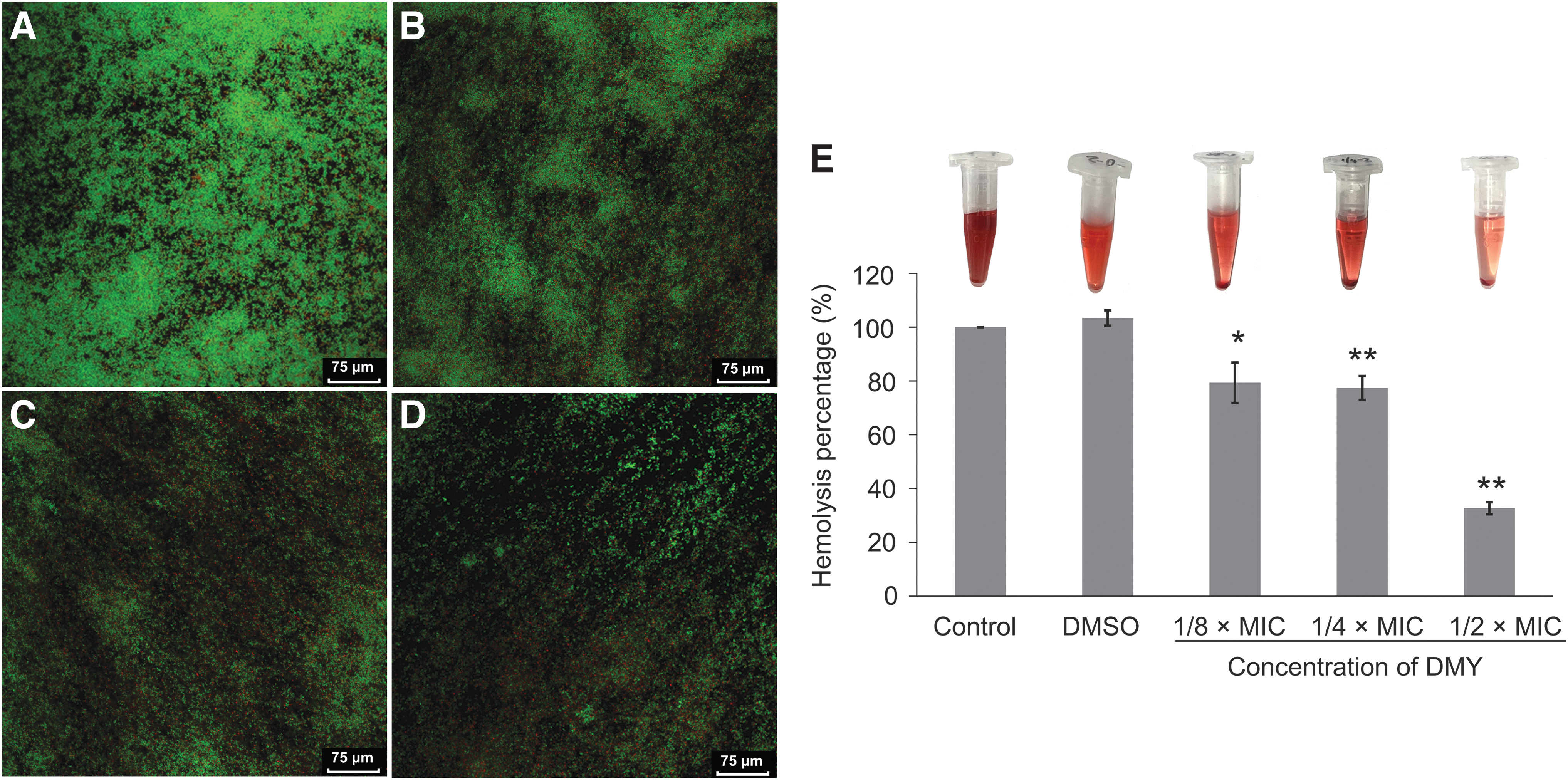
Confocal laser scanning microscopic images
In addition, the dense biofilm of control S. aureus was almost formed by green cells with strong fluorescence, suggesting that the untreated biofilm cells were viable and active. In comparison, as the concentration of DMY increased from 1/8 × MIC to 1/2 × MIC, the population of dead bacteria with red fluorescence increased, while that of live bacteria with green fluorescence decreased.
DMY attenuated hemolytic activity of S. aureus
As shown in Figure 1E, the degree of hemolysis of red cells of rabbit showed that DMY could obviously inhibit the hemolytic activity of S. aureus. The hemolytic percent in control group treated with PBS buffer served as 100%. Compared to the control, after cultured with 1/8 × MIC DMY, 1/4 × MIC, and 1/2 × MIC DMY, hemolysis percentages decreased to 79.3%, 77.4%, and 32.7%, respectively. Moreover, DMSO itself did not cause the hemolysis of rabbit erythrocytes.
Global analysis of the transcriptomic response
Compared with the control groups, there were 262 differentially expressed genes (DEGs) in DMY-treated group, containing 80 upregulated DEGs and 182 downregulated DEGs (Supplementary Fig. S1A). As shown in Table 1, gene-SAOUHSC_01110 and gene-SAOUHSC_01114 encoding fibrinogen-binding proteins and gene-SAOUHSC_02798 encoding surface protein G (SasG) were downregulated. Genes encoding iron-regulated surface determinants (staphylobilin-forming heme oxygenase IsdG, heme uptake protein IsdC, and heme transporter IsdA) were also downregulated. In addition, some genes involved in virulence, cell envelope, and amino acids metablism were significantly differentially expressed.
Differentially Expressed Genes Associated with Biofilm Formation, Virulence, Cell Envelope, and Amino Acid Metabolism in Staphylococcus aureus Following Treatment with 1/2 × Minimum Inhibitory Concentration of Dihydromyricetin
ATP, adenosine triphosphate.
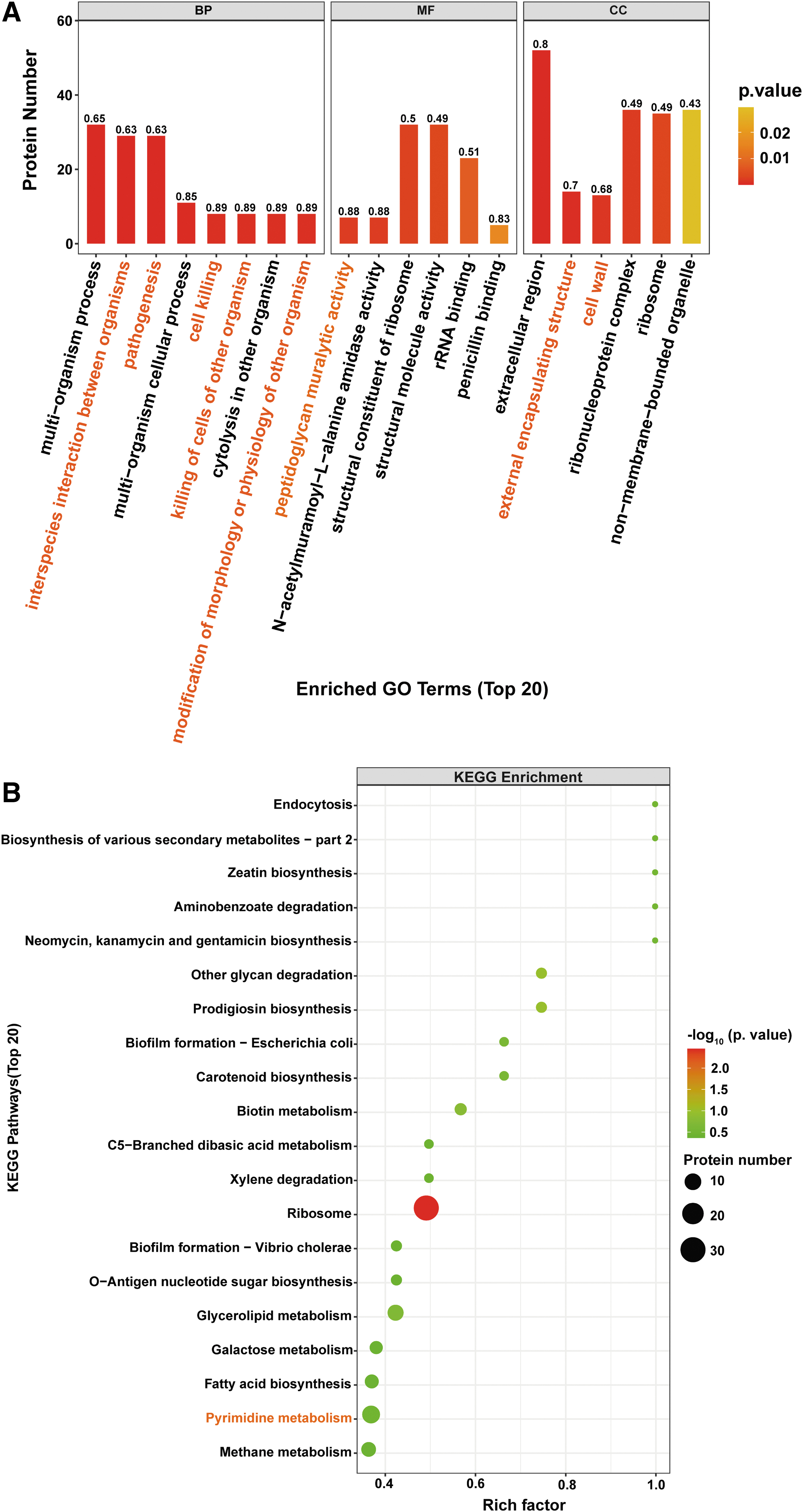
GO enrichment analysis was used to systematically analyze the DEGs functions (Fig. 2A). In biological processes, DEGs mainly annotated in pathogenesis (12), histidine biosynthetic process (7), and metabolic process (7). DEGs also classified into molecular functions and cellular components, including cell wall (5) and ribosome (12). Furthermore, DEGs were analyzed by KEGG enrichment analysis and annotated according to bioinformatics databases. As shown in Figure 2B, DEGs induced by DMY were mainly enriched in histidine metabolism (7) and ribosome (6).

The GO classification
The transcriptional profiles of DEGs from RNA-Seq were verified by RT-qPCR assay. As shown in Supplementary Figure S2, both RT-qPCR and transcriptome analyses showed the same expression trend for each of the four DEGs, indicating that the results of RNA-Seq were reliable.
Global analysis of the proteomic response
The volcano figures showed that the total differential proteins were 669 in the DMY-treated sample (Supplementary Fig. S1B), including 242 upregulated and 427 downregulated. As shown in Table 2, consistent with the transcriptomic results, the proteomic results showed the downregulation of surface-associated proteins (FnbA, FnbB) and clumping factor A (ClfA). Moreover, iron-regulated surface determinants (IsdA, IsdB, and IsdC), serine protease, and serine protease SplA were also downregulated. Notably, the results also verified that α-hemolysin was downregulated by DMY treatment. Immunoglobulin-binding protein Sbi, leukocidin LukS component, and γ-hemolysin component involved in bacterial pathogenesis were downregulated. Additionally, many proteins related to bacterial cell envelope were regulated by DMY, including D-alanine aminotransferase, lipid II isoglutaminyl synthase, FemA, FemB, ABC transporters, and permeases.
Differentially Expressed Proteins Associated with Biofilm Formation, Virulence, and Cell Envelope in Staphylococcus aureus Following Treatment with 1/2 × Minimum Inhibitory Concentration of Dihydromyricetin
ABC, ATP-binding cassette.
GO functional annotations showed that a large number of DEPs in biological processes were mainly enriched in pathogenesis (29), cell killing (8), and killing of cells of other organism (8) (Fig. 3A). Some other DEPs localized in ribosome (35) and cell wall (13). For the molecular functions, DEPs were mainly enriched in peptidoglycan muralytic activity (7) and rRNA binding (23). In addition, the KEGG pathway analysis of DEPs revealed that the DEPs were mainly mapped to the pathway of ribosome (30) (Fig. 3B), which is consistent with the RNA sequencing results.
The GO classification
Discussion
The transcriptome and proteome analyses exhibited a comprehensive view of the antibacterial mechanism of DMY against S. aureus (Fig. 4). Adhesion to surfaces is a major strategy by S. aureus at the early stage of biofilm formation, which involved in a variety of Staphylococcal surface proteins. ClfA, SasG, FnbA, and FnbB play a vital role both in cell attachment and accumulation for biofilm formation (Feuillie et al., 2017; Mazmanian et al., 2002). Kaempferol has been demonstrated to prevent S. aureus biofilm formation by decreasing the gene expression level of fnbA, fnbB, clfA, clfB, and sarA (Ming et al., 2017). Our transcriptome and proteome analyses indicated that DMY inhibited the expression levels of the genes and proteins (FnbA, FnbB, ClfA, SasG) related to biofilm attachment and accumulation stages.
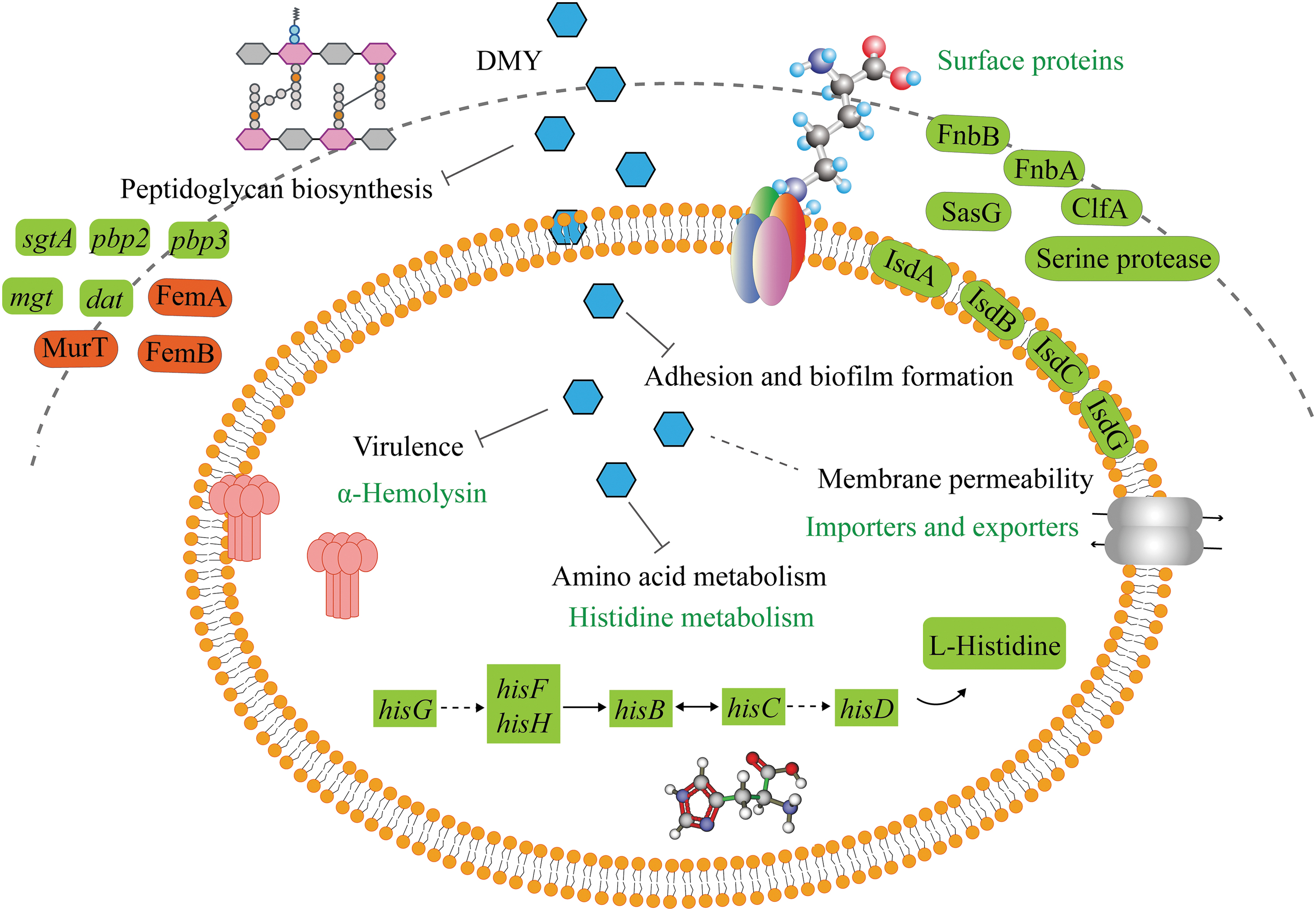
Schematic presentation of multitarget mechanisms for the antibacterial activity of DMY against Staphylococcus aureus. Red blocks in the diagram show the upregulated genes or proteins, while green blocks indicate the downregulated genes or proteins. DMY, dihydromyricetin.
In addition, iron-regulated surface determinant is necessary for the primary attachment and accumulation phase of biofilm formation (Missineo et al., 2014). As a consequence, regulation of iron-regulated surface determinants (IsdA, IsdB, IsdC, and IsdG) by DMY could promote the inhibition of S. aureus biofilm. Meanwhile, serine protease and serine protease SplA, which contribute to biofilm maturation (Fournier et al., 2001), were downregulated from proteomic results, implying that DMY might hinder the biofilm maturation of S. aureus by regulating serine protease expression (Fournier et al., 2001). It was noteworthy that no significant change in the expression of intercellular adhesion (ica) gene locus and its proteins were observed despite their importance in biofilm formation. This may be because S. aureus produced biofilm in an ica-independent manner in this study (Doulgeraki et al., 2017), or DMY principally targeted surface proteins rather than ica locus.
S. aureus causes chronic infections, mainly relies on its ability to produce various virulence factors that are closely related to host invasion and immune evasion (Park et al., 2019). Our results of hemolysis assay and proteome indicated that DMY could significantly reduce the hemolysin secreted by S. aureus. Gene sbi and its encoding protein immunoglobulin-binding protein Sbi were downregulated, which play a role in the inhibition of both the innate and adaptive immune responses. Moreover, γ-Hemolysin and leukocidin (Luk) toxins have lytic activity toward red blood cells (Date et al., 2014). Therefore, DMY suppressed the expression of toxin-ralated genes and proteins, so as to inhibit the pathogenesis of S. aureus.
Cell envelope of bacteria served as the first line of defense against antibacterial agents. In this study, DMY suppressed the expression level of D-alanine aminotransferase encoded by dat involved in the formation of bacterial cell wall, while upregulated the expression of the cell wall biosynthesis-associated protein lipid II isoglutaminyl synthase subunit MurT. In addition, DMY upregulated FemA and FemB which are required for peptidoglycan cross-linking, and downregulated multimodular transpeptidase-transglycosylase, penicillin-binding protein, and monofunctional glycosyltransferase (mgt) involved in peptidoglycan polymerase catalyzes glycan chain elongation. These results indicated that DMY disturbed the proper crosslinking of peptidoglycan, which resulted in peptidoglycan weakening, and eventually cell death.
Antimicrobial agents could inhibit biofilm formation and bacterial pathogenesis by changing the composition and permeability of membrane (Eder et al., 2017; Hobby et al., 2019). The cell membrane transportation function and permeability of S. aureus were considerably affected by DMY treatment. Proteins involved in membrane transport were extensively regulated, including proteins related to the process of heme transporters (IsdA, IsdB, IsdC, and IsdG), ABC transporters, and bacterial secretion system. In addition, various permeases such as glutamate permease, ABC transporter permease, and amino acid permease were differentially expressed (Table 2), suggesting the interference of cell membrane permeability in S. aureus under DMY treatment.
Meanwhile, cell envelope is important for metal ion homeostasis (Kern et al., 2010), biofilm formation (Holland et al., 2011), host tissue adhesion, and pathogenicity (Gerlach et al., 2018; Wanner et al., 2017). Thus, the extensive interference of DMY on the cell envelope of S. aureus might further hinder the biofilm formation and bacterial pathogenesis.
Transcriptome KEGG results suggested that the most significantly enriched KEGG pathway was the histidine pathway. Gene-SAOUHSC_03014 and gene-SAOUHSC_03015 encoding adenosine triphosphate phosphoribosyltransferase (hisG) and gene-SAOUHSC_03013 encoding histidinol dehydrogenase (hisD) were downregulated, which were participated in the first and last two steps of the histidine synthesis pathway, respectively (Chioccioli et al., 2020). In addition, gene-SAOUHSC_03012 associated with histidinol-phosphate aminotransferase (hisC) family protein and gene-SAOUHSC_03010 associated with imidazole glycerol phosphate synthase subunit HisH were downregulated. As a glycogen amino acid, histidine enters the gluconeogenesis pathway under unfavorable conditions (Feng et al., 2022).
Moreover, DMY not only perturbed biological functions of S. aureus but also affected the fundamental metabolic processes, such as alanine, aspartate and glutamate metabolism, purine and pyrimidine metabolism, pyruvate metabolism and so on, which are vital for cell synthesis. The disturbance of purine and pyrimidine metabolism might inhibit the energy metabolism and membrane transport systems (Cheng et al., 2021), and reduce the resistance of S. aureus to stressful environment (Du et al., 2020). These results indicated that DMY probably cause instability of intracellular biological process and bacterial growth metabolic systems, so as to lead to S. aureus damage or death.
In summary, DMY not only damaged the biofilm architecture of S. aureus, but also remarkably inhibited the hemolytic activity. The antibacterial activity of DMY seems to be predominantly achieved by inhibiting the expression level of the genes and proteins related to biofilm formation, including clumping factor A (ClfA), iron-regulated surface determinant (IsdA, IsdB, and IsdC), surface protein (SasG), fibrinogen-binding proteins (FnbA, FnbB), and serine protease. In addition, DMY disturbed multiple targets, including cell envelope, membrane transport, histidine metabolism, purine and pyrimidine metabolism, and pyruvate metabolism. These findings provided new insights into the antibacterial mechanism of DMY against S. aureus, giving a meaningful guidance for the study of the molecular targets of other plant-derived flavonoids on foodborne pathogens.
Footnotes
Authors' Contributions
W.R.: Conceptualization, methodology, formal analysis, writing—original draft, and writing—review and editing. Y.Y.: Methodology and formal analysis. F.L.: Formal analysis, and validation. K.Z.: Resources and methodology. J.B.: Validation and resources. Y.X.: Formal analysis. Q.B.: Validation and resources. Y.H.: Resources and supervision. Y.W.: Conceptualization, writing—review and editing, funding acquisition, and supervision. H.G.: Conceptualization, writing—review and editing, and funding acquisition.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 31901791 and No. 32072321).
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.