
Abstract
Vibrio parahaemolyticus is a foodborne pathogenic bacterium commonly found in seafood. The emergence of drug-resistant strains poses a threat to human public health and economic development. Therefore, there are increasing needs to develop new technologies in controlling multidrug-resistant V. parahaemolyticus strains and to evaluate their practical efficiency in seafood or mariculture. In this study, we screened two genetically related V. parahaemolyticus phages, F23s2 and H256D1, which belonged to the siphoviridae family and podoviridae family, respectively. They showed 97.13% and 96.13% identity with Vibrio phage vB_Vpap_MGD1, respectively. Both phages were stable at pH 4–11 and displayed temperature tolerance (<70°C). Meanwhile they showed a broad host spectrum for multidrug-resistant V. parahaemolyticus, and Phage F23s2 lysed 16 of all 23 V. parahaemolyticus strains, while phage H256D1 lysed 10 strains. Phage F23s2 and H256D1 had a good inhibitory effect on V. parahaemolyticus in shrimp meat. Compared with the negative group, the bacterial amount of experimental group with phage F23s2 decreased by 1.60 log colony-forming unit (CFU)/mL at 12 h. For phage H256D1, the bacterial concentration of shrimp meat contaminated with V. parahaemolyticus H256 increased to 5.65 log CFU/mL at 72 h, while the concentration of the experimental group in presence of phage H256D1 was 3.58 log CFU/mL. All live clams infected with V. parahaemolyticus died after 96 h in the absence of phage, whereas clams with phage F23s2 and H256D1 still had a survival rate of 12% and 4%, respectively. Understanding the gene function and biology of phages facilitates its application for control of V. parahaemolyticus contamination worldwide.
Introduction
V
In recent years, the overuse of antibiotics in aquaculture has led to the development of antibiotic resistance in V. parahaemolyticus. It has been reported to be resistant to ampicillin (98.5%), cefuroxime (75.6%), cefepime (66.7%), piperacillin (59.1%), and ampicillin/sulbactam (50.0%) (Sun et al., 2022). The emergence of drug-resistant strains also poses a threat to human public health and economic development, and it costs $21 million per year to address the health problems caused by V. parahaemolyticus-contaminated seafood in the United States (Tan et al., 2020).
Phages are abundant on Earth and are harmless to other organisms other than their target hosts (Salmond and Fineran, 2015). As an alternative to antibiotics, phages have become increasingly popular for treating diseases in humans and animals, as well as for detecting and fighting bacterial contamination in food (Nagel et al., 2016). The U.S. Food and Drug Administration approved the use of Listeria-specific phage P-100 as food additives in certain ready-to-eat products such as instant scallop (Monk et al., 2010). To prevent listeriosis in dairy products, PhageGuard Listex (Intralytix) and ListShield, commercial phage products, were administered to artificially contaminated surface-ripened red-smear soft cheese, resulting in a significant reduction of Listeria (Guenther and Loessner, 2011; Perera et al., 2015).
In this study, two genetically related phages F23s2 and H256D1 were isolated. The biological characteristics of these two phages were then clarified, and the control effect of phages on V. parahaemolyticus in shrimp meat and living clam was determined.
Materials and Methods
Isolation and purification of phages
Chinese Yangcheng Lake hairy crabs (Yangcheng Lake, Jiangsu Province, China) were used as samples for the preliminary isolation of phages. The method of Alagappan et al. (2010) was adopted to isolate and purify the phages. Twenty-five grams of hairy crab meat and guts was weighed, chopped, and added to 75 mL of saline and then homogenized. The homogenate was filtered and taken as the phage stock solution. V. parahaemolyticus F23 (foodborne isolate) and H256 (a drug-resistant mutant of a foodborne isolate F7) (Jin et al., 2021) were used as host bacteria.
The host bacteria and phage were mixed with Luria-Bertani (LB) broth (Oxoid, Hants, United Kingdom) and incubated at 37°C with shaking (160 rpm) for 24 h. After incubation, the mixtures were centrifuged at 8500 rpm for 15 min and filtered with 0.22 μm filters (Millipore, Darmstadt, Germany). The phages in filtered samples were detected by using the double-layer plate method. Subsequently, purification of phage was performed; a single phage was considered to be obtained after the phage spots that appeared on the double layer plate were of uniform size.
Transmission electron microscopy of phage
Phage F23s2 and H256D1 were ultracentrifuged at 40,000 rpm for 1 h at 4°C. The precipitate was resuspended with 1 mL phosphate-buffered saline (PBS). Phage suspensions were stained with phosphotungstic acid and placed on copper webs and observed under transmission electron microscopy (TEM) (Hitachi, Tokyo, Japan).
Analysis of phages genome
Phage DNA was extracted from phage by phenol/chloroform/isoamyl alcohol method (Xing et al., 2017) and sequenced using the Illumina Hiseq platform (Illumina, San Diego, CA) in Shanghai Zeta (Shanghai, China). The GeneMarks was employed to determine its open reading frame. The online website RAST was used for initial genome annotation and the gene function was further annotated by NCBI's blast against Genbank and Swiss-Prot database. The virulence gene of the phage genome was analyzed using the Virulence Factors of Pathogenic Bacteria database. The phylogenetic tree based on phage large terminal subunit was constructed by MEGA7.0 (Hall, 2013), while the whole genome circle map was drawn using the software BRIG (Alikhan et al., 2011). Comparison and analysis between phage genomes were performed with Easyfig (Sullivan et al., 2011).
Multiplicity of infection for phages
The multiplicity of infection (MOI) was the ratio of phage to host bacteria. The phage solution (1.0 × 105 plaque-forming unit [PFU]/mL) and different concentrations of host bacteria were mixed, and seven mixtures with MOI of 0.001, 0.01, 0.1, 1, 10, 100, and 1000 were obtained and incubated at 37°C for 3.5 h with shaking at 160 rpm. After incubation, the mixtures were centrifuged at 12,000 rpm for 10 min at 4°C and filtered with 0.22 μm filters to obtain supernatant. The titer of the phage in the supernatant was measured by the double-layer plate method. The optimal MOI of phages was determined by highest titer. The experiment was repeated three times.
One-step growth curve of phages
In general, the latent period, lysis period, and burst size of phages were determined by the one-step growth curve (Li et al., 2020). The phage (1.0 × 104 PFU/mL) was mixed with host bacterial at optimal MOI and incubated for appropriate time. The mixture was centrifuged at 12,000 rpm for 30 s, and the precipitate was resuspended twice with LB medium. Then the suspension was immediately incubated at 37°C for 2 h. The samples were taken every 10 min during incubation and the phage titers were measured by double-layer plate method. The experiment was repeated three times.
Bacterial inhibition by phages
The host bacterium F23 and H256 were diluted to 1.0 × 105 colony-forming unit (CFU)/mL. Phage and host bacteria were mixed in 96-well plates (Corning, Shenzhen, China) to obtain seven mixtures with MOI of 0.001, 0.01, 0.1, 1, 10, 100, and 1000, respectively. The OD600 values of the mixtures were measured every 1 h for 24 h using an automated cell growth curvature meter (Bioscreen C MBR, Finland) to determine the inhibition effect of different concentrations of phage on the host bacteria. The host bacterium grown in LB medium was served as a negative control, whereas the LB medium was acting as blank control. The experiment was repeated three times.
Resistance to the environment
The phage was diluted to 1.0 × 105 PFU/mL. The diluted phage was placed in a constant temperature water bath at 20°C, 30°C, 40°C, 50°C, 60°C, 70°C, and 80°C, respectively. Phage titers were measured at 0, 30, and 60 min, respectively.
One hundred microliter of 1.0 × 106 PFU/mL phage was mixed with 900 μL of PBS buffer at different pH (2–13) in 1.5 mL centrifuge tubes. After placing the mixtures in a 37°C water bath for 2 h, the phage titers were determined by bilayer plate method.
Host spectrum of phages
Twenty-three strains of V. parahaemolyticus, which were laboratory preserved, were used to determine host range of phage F23s2 and H256D1, including 2 standard strains, 6 foodborne isolates, 12 clinical isolates, and 3 drug induced resistant mutants (Supplementary Table S1; Jin et al., 2021). Phage host profiles were determined by the presence or absence of phage spots using the double-layer plate method. The experiment was repeated three times.
Inhibitory effect of phage on V. parahaemolyticus in shrimp meat
According to the experiment method of Zhang et al. (2018), shrimp meat that has been heated in boiling water for 30 min was cut into small pieces of 2 × 2 cm and 50 μL phage and 50 μL host bacteria with ratio of 100:1 were inoculated into the shrimp meat, respectively. The final concentration of V. parahaemolyticus was 1.0 × 104 CFU/g. The shrimp meat was then kept at 15°C. Samples were collected at 0, 6, 12, 24, 48, and 72 h, respectively, and the concentration of bacteria was determined by plate counting. Only bacteria were inoculated in the negative control, and the same volume of PBS was used to replace phages and bacteria in the blank control.
Effect of phage on the survival of live clams infected with V. parahaemolyticus
Live adult clams were purchased from Aeon Supermarket, Wuhan, Hubei Province, China. Referring to the experiment of Lomeli-Ortega et al. (2014), live clams were sterilized with 0.3 ppm chlorine dioxide (ClO2) for 5 min, and cultured in artificially prepared seawater (0.9% NaCl) at 28°C for 24 h for acclimation, after which healthy clams were selected for continued aquaculture. Bacteria and bacteriophages at concentrations of 1.0 × 104 CFU/mL and 1.0 × 106 PFU/mL were then added to cultured seawater, and phage and bacterial concentrations were maintained for 24 h. The live clams cultured in artificially prepared seawater were considered the control group, the clams treated only with V. parahaemolyticus and the groups treated with V. parahaemolyticus and phages were used as experimental groups. And the survival rate of clam (the number of surviving clams/the original number of clams) was calculated.
Statistical analysis
In this study, the results in the experiments were compared and analyzed by students' t-test, and a p-value <0.05 was considered significant.
Results and Discussion
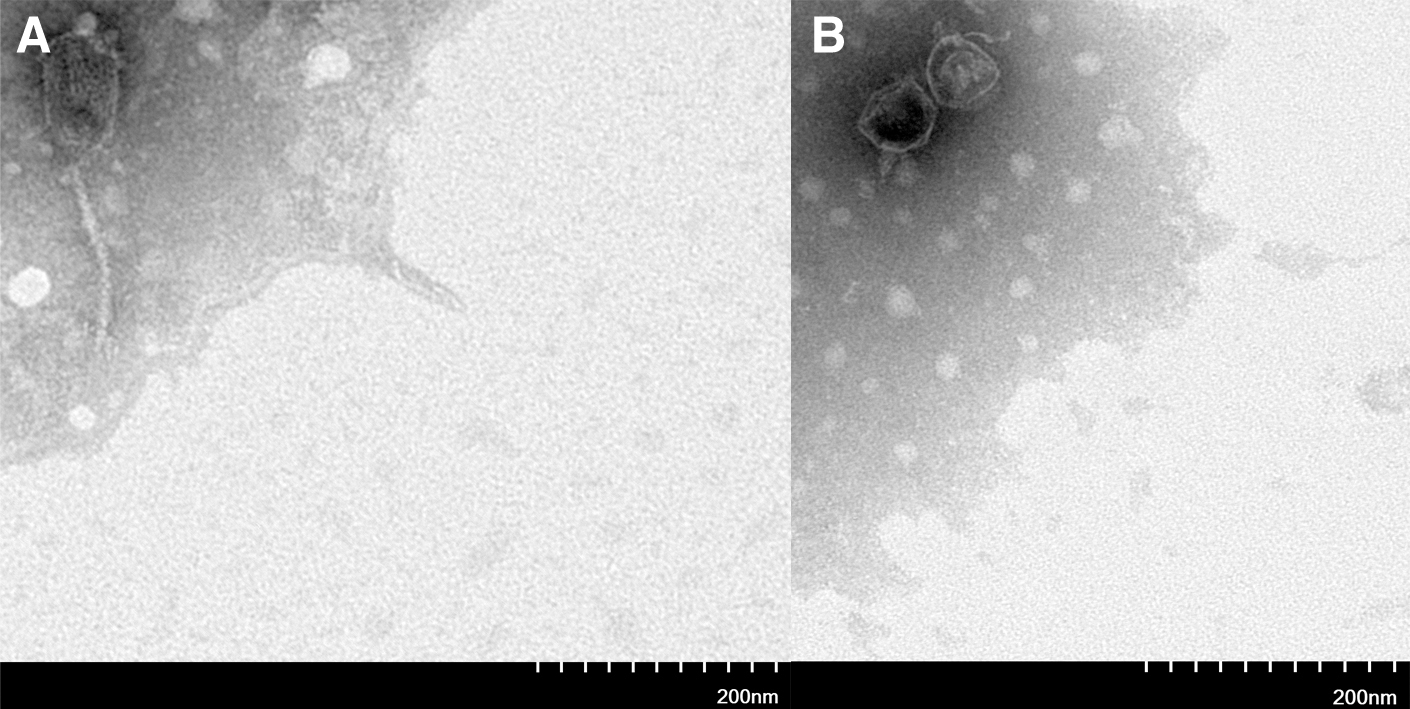
TEM of phage
Two phages (F23s2 and H256D1) were observed by TEM. Phage F23s2 had a head length of 111 ± 3 nm, width of 60 ± 2 nm, and a tail length of 157 ± 1 nm, which belonged to Siphoviridae family (Fig. 1A). The head length and width of phage H256D1 were 63 ± 1 and 52 ± 2 nm, respectively, and the tail length was 27 ± 1 nm (Fig. 1B), belonging to Podoviridae family.
Genome-wide analysis of phages
The whole genome of phage F23s2 (Accession: ON080941) had 43386 bases (49.40% G+C) and 49 open reading frames (ORFs), and phage H256D1 (Accession: ON080940) had 35587 bases (49.80% G+C) and 38 ORFs. The software SnapGene revealed that the genes of phage F23s2 were all located on the negative strand (Fig. 2A); in contrast, the genes of phage H256D1 were all located on the positive strand. In addition, no virulence gene was found in these two phages.
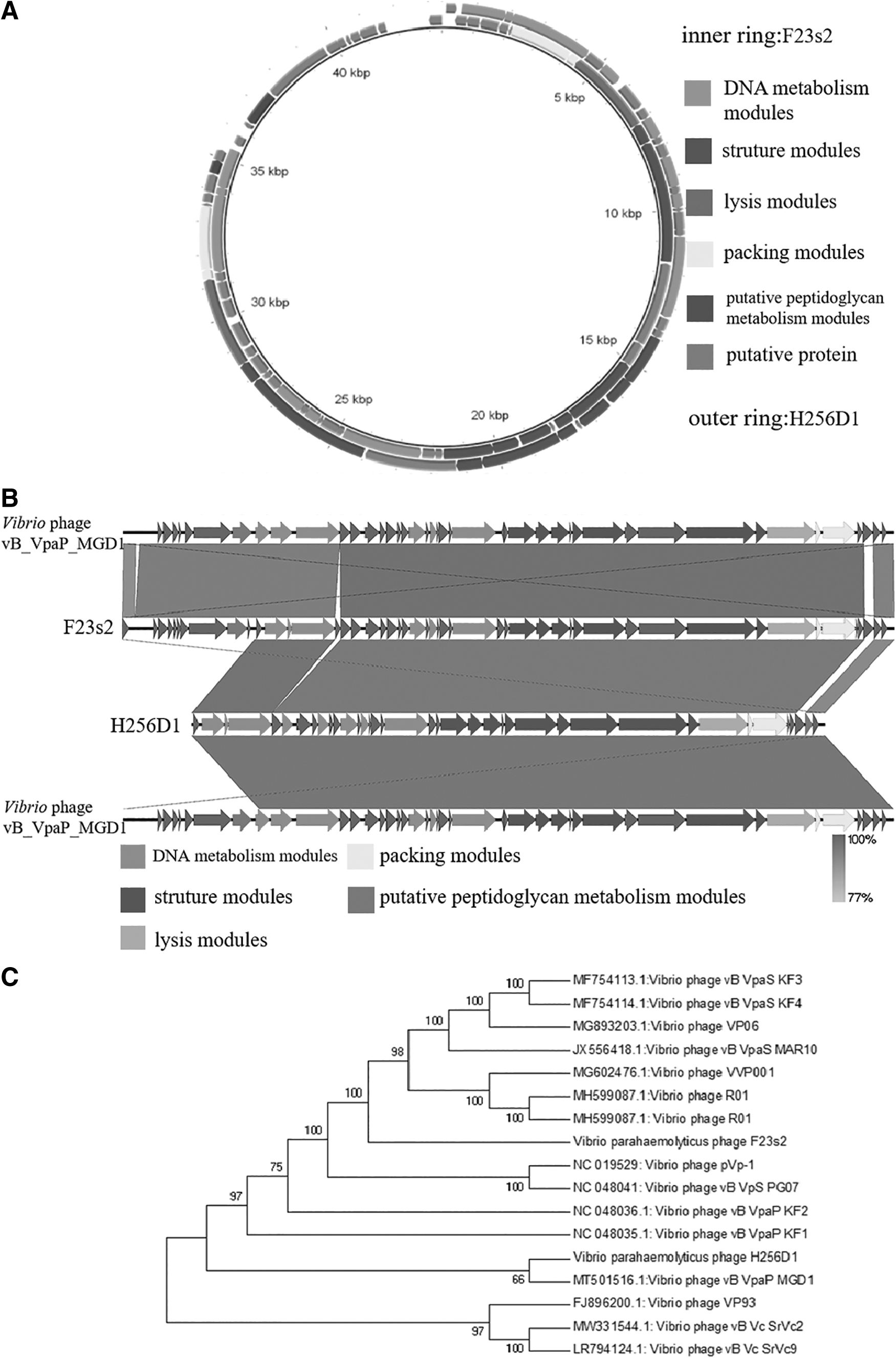
The homology between them at the nucleic acid level reached 96.33%, exceeding 95%, so that the two phages belonged to the same species according to the viral classification criteria of the International Committee for the Classification of Viruses (Adriaenssens and Brister, 2017). However, there was only 80% coverage between phage F23s2 and H256D1. Both phage F23s2 and H256D1 showed high homology with Vibrio phage vB_Vpap_MGD1 (Fig. 2B), The coverage between phage F23s2 and phage vB_Vpap_MGD1 was 98%, and the identity was 97.13%, while phage H256D1 showed 82% coverage and 96.13% identity with phage vB_Vpap_MGD1.
A phylogenetic tree based on the protein sequence of the large subunit of phage terminal enzyme was constructed, and the relationship between phages F23s2 and H256D1 and other 15 V. parahaemolyticus phages in NCBI Genbank database was analyzed. The results showed that phage vB_VpaP_MGD1 and H256D1 were most closely related and formed a cluster, suggesting that they may have originated from the same ancestor. The closest relatives of phage F23s2 were V. parahaemolyticus phages R01 and VVP001 (Fig. 2C).
Their genes can be classified into five categories based on their functions. The phage H256D1 genome included 9 annotated ORFs associated with phage structure modules, and F23s2 genome contained only 7 structure-related ORFs, which showed more than 95% identity with the homologous genes of H256D1 (Fig. 2B). However, ORF40-ORF49 of phage F23s2 did not exist in phage H256D1 genome, which might be involved in controlling the morphology of the phage tail, and be the reason why the phage F23s2 belonged to Siphoviridae family and the H256D1 belonged to Podoviridae family.
The two phage genomes shared homologous annotated genes in packaging and lysis modules, including two endolysin proteins named LysF23s2 and LysH256D1. The two proteins had the same sequence length and differed only at sites 37, 131, 149, 162, 307, 357, 370, and 681. Of these, the aspartic acid of LysF23s2 at site 37 was an acidic amino acid, while the asparagine of LysH256D1 was a hydrophilic amino acid. And the same was true for amino acid site 149, but the opposite was the case at site 162 where glutamine was hydrophilic in the former and glutamate was an acidic amino acid in the latter. Also, the amino acids of LysF23s2 at sites 131 and 681 were both lysine, which was basic amino acids, and those of LysH256D1 were alanine and leucine, respectively, which belonged to hydrophobic amino acids. Interestingly, at site 307, one was a hydrophobic alanine and the other was an acidic glutamate. In general, differences in amino acid residues in the protein sequence may lead to distinctions in the overall structure and hydrophobicity of the protein (Yan et al., 2017). The results showed that there was indeed some difference between their endolysin activities (unpublished data), and detailed analysis of two proteins could help us to understand the relationship between protein sequence and function (Wu et al., 2019).
Optimal MOI values for phages
The results showed that the optimal MOI of phage F23s2 and H256D1 was 0.001 and 0.01, respectively (Table 1). When the MOI was 0.001, the titer of phage H256D1 was 1.1 × 108 PFU/mL, which was lower compared with (9.4 × 108 PFU/mL) when the MOI was 0.01. It indicated that more host bacteria did not increase the titer of phage H256D1. This might be related to bacteria-phage co-evolution, that was, when the concentration of bacteria was high enough, the phage invasion rate might be reduced (Koskella and Brockhurst, 2014). Tan et al.'s (2015) research showed Vibrio anguillarum PF430-3 could form colonies and reduce the invasion rate of phage to assure its own survival when Vibrio phage KVP40 invaded.
Multiplicity of Infection of Phage F23s2 and H256D1
CFU, colony-forming unit; PFU, plaque-forming unit.
One-step growth experiment
The one-step growth curves for phage F23s2 and H256D1 displayed differences in the parameters of infection cycles between two phages (Fig. 3A, B). The phage F23s2 showed a latent period of 0–20 min and lysis period of 20–80 min with a burst size of 12 PFU/cell. This result was similar to that of the V. parahaemolyticus phage vB_VpaP_DE10 with a latent period of 0–25 min and an outbreak of 19 PFU/cell (Ye et al., 2022).
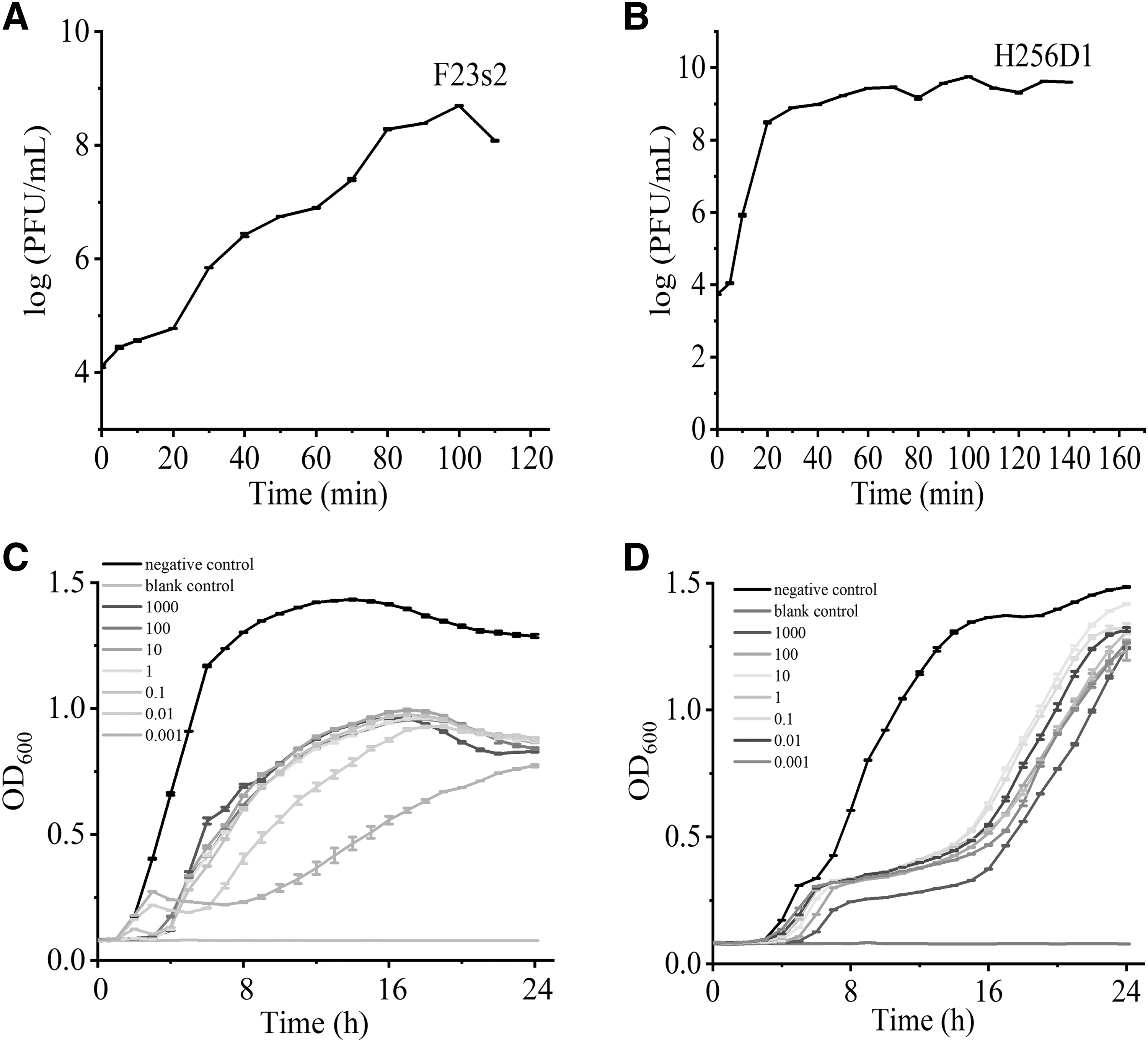
On the other hand, the latent period and lysis period of the phage H256D1 were 0–5 and 5–60 min, respectively. Average burst size of the phage H256D1 was 131 PFU/cell. The burst amount of phage H256D1 was nearly 10 times higher compared with F23s2, which was basically consistent with the results of the optimal MOI. In principle, phages with short latent period and a large outbreak were effective to suppress bacteria. Abedon et al. (2019) increased the outbreak of T-even phages by extending latent period.
Determination of bacteriophage lytic activity
The lytic activity of phages was determined by measuring the OD600 value of host bacteria cultures at MOI ranging from 0.001 to 1000. Phage F23s2 had the best inhibitory effect on V. parahaemolyticus F23 at an MOI of 0.001. Compared with the negative control, the OD600 value of V. parahaemolyticus decreased by 1.017 and 0.515 at 7 and 24 h, respectively (Fig. 3C). Phage F23s2 could inhibit bacterial proliferation for a long time, which was the basis of phage screening for applications (Endersen et al., 2014).
Phage H256D1 could effectively inhibit the growth of V. parahaemolyticus H256 from 0 to 15 h. Compared with the negative control, the OD600 value of V. parahaemolyticus decreased by 0.82–1.02 at 15 h at MOI of 0.001–1000, showing the best inhibitory effect at MOI of 1000 (Fig. 3D). In terms of bactericidal efficacy, phage H256D1 was more similar to those phages that have been successfully used. Sundarram et al. (2021) evaluated the efficacy of six concentrations (0.0001 × , 0.001 × , 0.01 × , 0.1 × , 1 × , and 10 × ) of Listeria monocytogenes phage against Listeria spp., and found that 1 × and 10 × phage treatments had the greatest efficacy. This indicated that higher concentrations of L. monocytogenes phages were more effective in controlling Listeria contamination in food and food processing (Sundarram et al., 2021).
Environmental resistance of phages
After thermal treatment, both phages were basically stable at 20–60°C (Fig. 4A, C). The phage titers remained 3.04 log PFU/mL and 4.00 log PFU/mL after 60 min of inactivation at 70°C, respectively. Meanwhile, phage F23s2 and H256D1 maintained high activity at pH 4–10 and 4–9, respectively, and phage titer of H256D1 retained 3.85 log PFU/mL at pH 11 (Fig. 4B, D).

V. parahaemolyticus phages vB_VpaS_OMN, vB_VpS_BA3, vB_VpS_CA8, and Vp1 were able to maintain its activity at pH 4–11 and under temperature less than 60°C (Stalin and Srinivasan, 2016; Yang et al., 2020). It indicated that the two phages in this study showed better heat resistance. Currently, high-temperature treatment is the most common sterilization method in food processing (Aaliya et al., 2021); phages with high heat tolerance have a wider application prospect.
Host rang of phage
There were some differences in host ranges between phage F23s2 and H256D1 (Supplementary Tables S1). Phage F23s2 lysed 16 of all 23 V. parahaemolyticus strains, while phage H256D1 lysed 10 strains. The host range of phage was mainly determined by phage tail fibronectin (Harada et al., 2018). Yehl et al. (2019) identified host range determining regions in the tail fibronectin of T3 phage and successfully expanded the host range of phage by modifying these regions using site-directed mutagenesis.
In this study, the tail fibronectin sequences of phage F23s2 and H256D1 were compared, and it was found that the length of the two proteins was the same, and only the amino acids at 32, 34, and 170 sites were different. The amino acids at these positions were methionine, aspartic acid, and lysine in F23s2, and threonine, glycine and glutamine in H256D1, respectively. Therefore, it was speculated that some specific amino acid differences in their tail fibronectin might contribute to their different host range.
The phage could inhibit V. parahaemolyticus in shrimp
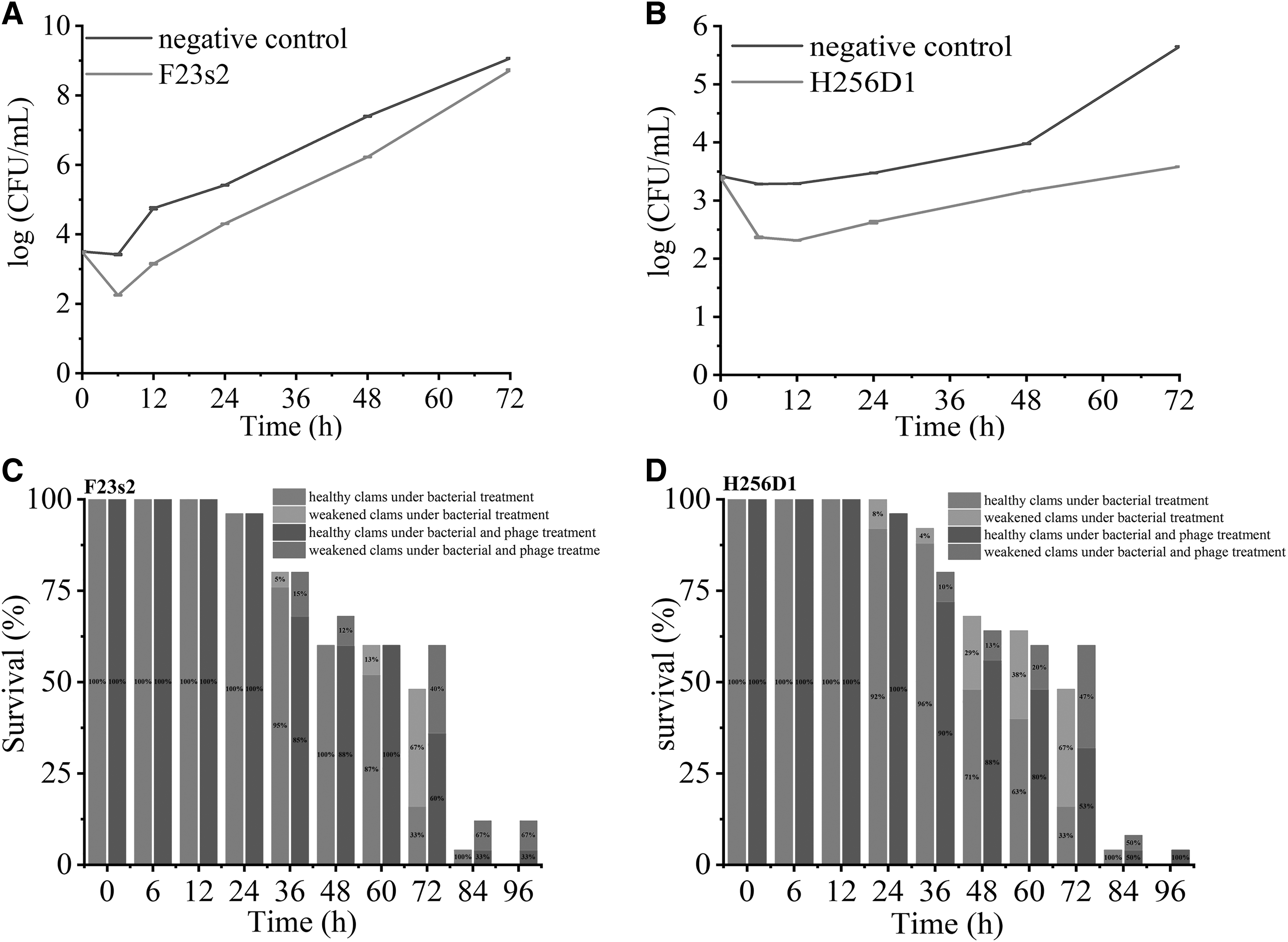
Referring to the experiment of Zhang et al. (2018), the experimental temperature was controlled at 15°C to simulate the preservation conditions of shrimp in the retail markets. In the negative control, the V. parahaemolyticus F23 concentration increased sharply within 72 h, from 3.51 log CFU/mL to 9.06 log CFU/mL (Fig. 5A). In the experimental group supplemented with phage F23s2, the concentrations of V. parahaemolyticus F23 were 2.25 log CFU/mL and 3.15 log CFU/mL at 6 and 12 h, respectively, while those in the negative control group were 3.42 log CFU/mL and 4.75 log CFU/mL, respectively. This was similar to the results of Zhang et al. (2018), in which, the concentration of V. parahaemolyticus decreased by 0.83 log CFU/mL at 6 h in the experimental group treated with phage OMN, and 2.9 log CFU/mL when the time was extended to 48 h.
Application of phage F23s2 and H256D1.
In the negative control, the concentration of V. parahaemolyticus H256 increased from 3.42 log CFU/mL to 5.65 log CFU/mL at 72 h. Meanwhile, the concentration of V. parahaemolyticus in the presence of phage H256D1 increased slightly from 3.39 log CFU/mL to 3.58 log CFU/mL (Fig. 5B). This result indicated that the contamination of shrimp meat with V. parahaemolyticus could be effectively reduced by the addition of phage H256D1.
Dubey et al. (2021) sprayed phage V5 on shrimp and observed a 78.1% reduction in the number of V. parahaemolyticus within 1 h of phage application. Teng et al. (2022) developed a phage cocktail and applied it to the surface of ready-to-eat crayfish meat, resulting in a 2.36 log CFU/mL reduction in the V. parahaemolyticus load. These reports showed that phages were efficient in controlling V. parahaemolyticus and could be used in practice.
Phages improved the survival of live clams infected with V. parahaemolyticus
At 72 h, the survival rate of clams treated only with V. parahaemolyticus was only 48%, and that in the experimental group supplemented with phage F23s2 and H256D1 was 60%. All live clams died after 96 h of V. parahaemolyticus inoculation, and the survival rates of clams treated with phage F23s2 and H256D1 were 12% and 4%, respectively (Fig. 5C, D). It indicated that phages could inhibit V. parahaemolyticus infection in living clams and this was consistent with the results of Lomeli-Ortega et al. (2014). After treatment with V. parahaemolyticus phages V3S and Vpms1, the survival of juvenile shrimp increased by 10–20% compared to the negative control (Lomeli-Ortega and Martinez-Diaz, 2014). Our results showed that phage therapy can effectively reduce V. parahaemolyticus infection in live clams and control clam mortality. This suggested that phage can be used as more than just a preventive. (Martinez-Diaz and Hipolito-Morales, 2013).
Conclusion
This study exhibited that both phage F23s2 and H256D1 possessed good antibacterial effects on V. parahaemolyticus, and can also be used to decontaminate V. parahaemolyticus in live clams. Meanwhile, the studies on the two phages made a contribution to clarify the relationship between phage gene sequences and functions, and provided valuable information for the application of phages in the field of antibacterials.
Footnotes
Disclaimer
This article has not been published or presented elsewhere, and is not under consideration by any other journal.
Authors' Contributions
W.L. and L.H. isolated and purified the two phages used in this experiment, designed the experiment, and prepared the article; H.X. had completed experiments related to the biological properties of two phage strains; H.W. and W.H. solved the problem of data processing; and M.Z. and H.W. are responsible for reviewing and revising the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Wuhan Knowledge Innovation Special Basic Research Project (2022020801010390), the National Key R&D Program of China (2017YFC1600100), and the school-established research projects of Wuhan Polytechnic University (2021Y02).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.