
Abstract
The Gram-positive bacterium Listeria monocytogenes causes a significantly high percentage of fatalities among human foodborne illnesses. Surface proteins, specifically expressed from a wide range of L. monocytogenes serotypes under selective enrichment culture conditions, can serve as targets for the isolation of this pathogen using antibody-based methods to facilitate molecular detection. In this study, monoclonal antibodies (MAbs), previously raised against the L. monocytogenes LPXTG surface proteins LMOf2365_0639 and LMOf2365_0148, were investigated for their ability to isolate L. monocytogenes from bacterial samples with immunomagnetic separation (IMS). Only 1 out of 35 MAbs against LMOf2365_0639, M3644, was capable of capturing L. monocytogenes. Among all the 24 MAbs examined against LMOf2365_0148, 4 MAbs, M3686, M3697, M3699, and M3700, were capable of capturing L. monocytogenes cells specifically from abbreviated primary selective enrichment cultures in either Palcam or LEB/UVM1 media or from mixed samples containing target and nontarget bacteria. MAb M3686 showed a unique specificity with the capability to capture strains of seven L. monocytogenes serotypes (1/2a, 1/2b, 1/2c, 3a, 4a, 4b, and 4d). These promising MAbs were subsequently characterized by quantitative measurements of antigen-binding affinity using surface plasmon resonance analysis and epitope mapping using overlapping recombinant polypeptides. The usefulness of these MAbs to LMOf2365_0148 in bacterial capture was consistent with their high affinities with K D constants in the nanomolar range and can be explored further for the development of an automated IMS method suitable for routine isolation of L. monocytogenes from food and environmental samples.
Introduction
The Gram-positive bacterium, Listeria monocytogenes, causes a rare but serious foodborne illness, known as listeriosis, which often results in fatality rates of 30% in susceptible individuals (Mead et al., 1999). L. monocytogenes is widely distributed in the environment (Farber and Peterkin, 1991; McLauchlin et al., 2004) and can easily contaminate food processing environments and foods (Beumer and Hazeleger, 2003). Moreover, some ready-to-eat foods, which support the growth of L. monocytogenes, are frequently implicated vehicles of listeriosis infection (Swaminathan and Gerner-Smidt, 2007) as contaminating L. monocytogenes cells in food products can multiply at refrigeration temperatures. Due to the high fatality rate of listeriosis in susceptible individuals and the fact that L. monocytogenes is present in various natural environments, it is important to make a continued effort to prevent L. monocytogenes contamination in foods.
Nucleic acid-based molecular detection of L. monocytogenes in environmental and food samples plays a crucial role in reducing the risk of transmitting this pathogen to humans. Preparation of high-quality samples for downstream molecular detection methods becomes a bottleneck in pathogen detection processes. Although molecular detection methods such as polymerase chain reaction (PCR) and reverse transcriptase–PCR have expedited L. monocytogenes detection, these methods still rely on the purity of DNA or RNA templates. PCR inhibitors from food matrices (Liu, 2008) and selective enrichment cultures (Fluit et al., 1993) as well as genetic materials from nontarget bacteria (Hsih and Tsen, 2001) in samples can confound test results and are thus challenges in achieving accurate detection. Moreover, background microflora is a continued issue hampering L. monocytogenes detection even using selective enrichment culture (Garrec et al., 2003).
Selective enrichment cultures enrich for all Listeria species and certain species such as Listeria innocua can outcompete the growth of L. monocytogenes (Besse et al., 2010; Oravcova et al., 2008). Isolation of L. monocytogenes from environmental and food samples or selective enrichment culture can eliminate the presence of interfering materials in the PCR assays and offers a solution to the accurate detection of the target pathogen by molecular methods.
Antibody-based isolation of foodborne pathogens from complex food matrices or enrichment cultures is often based on immunomagnetic separation (IMS) using monoclonal antibodies (MAbs) coated on magnetic beads. Advantages of using the IMS method include concentration of targeted viable bacteria, removal of inhibitory factors, and reduction of background microflora that may confound molecular test results. Reports have documented the development of MAbs against surface proteins for L. monocytogenes isolation (Mendonca et al., 2012; Paoli et al., 2007). Strains of L. monocytogenes are serologically differentiated into at least 14 serotypes and phylogenetically grouped into 4 distinct lineages (I, II, III, and IV) (Orsi et al., 2011; Yin et al., 2019). Nonetheless, an L. monocytogenes-specific MAb capable of isolating strains of various serotypes is lacking (Mendonca et al., 2012; Paoli et al., 2007).
This work was undertaken to identify and characterize MAbs capable of isolating L. monocytogenes from culture samples. This involved screening a panel of MAbs previously raised against recombinant proteins rLMOf2365_0639 (Zhang et al., 2016) and rLMOf2365_0148 (Zhang et al., 2022), derived from L. monocytogenes serotype 4b strain LI0521, for L. monocytogenes capture ability. The functions of Lmof2365_0148 and Lmof2365_0639 have not been elucidated. The two proteins are a type of surface proteins with a conserved LPXTG motif that allows covalent anchoring of the proteins to the cell wall peptidoglycan. Sortase A is involved in the cell surface presentation of Lmo0130 and Lmo0610 (Mariscotti et al., 2012), the homologues of Lmof2365_0148 and Lmof2365_0639, respectively.
Selected MAbs were assessed by IMS for their ability to capture seven serotypes of L. monocytogenes commonly isolated from environmental and clinical samples, five other Listeria species, and two other foodborne pathogens (Table 1). The capture ability of the MAbs was also tested with L. monocytogenes grown in selective enrichment cultures according to Health Canada's Compendium of Analytical Methods, MFHPB-07 (Warburton et al., 2011) and MFHPB-30 (Pagotto et al., 2011). Furthermore, selected MAbs were characterized by epitope mapping and affinity analysis (i.e., determination of the equilibrium dissociation constants [K D] by surface plasmon resonance [SPR] analysis), which are relevant to the MAb capture ability.
Bacterial Strains Used for Immunomagnetic Separation
n/a, not applicable.
Materials and Methods
Bacterial culture
Bacterial cells (Table 1) were cultured for this study as outlined in the Supplementary Materials and Methods section (Supplementary File S1).
Screening of anti-LMOf2365_0148 and anti-LMOf2365_0639 MAbs for bacterial capture ability
Anti-LMOf2365_0639 and anti-LMOf2365_0148 MAbs were assessed for capturing live L. monocytogenes by coating bacteria with MAbs followed by IMS with magnetic beads conjugated with sheep anti-mouse IgG (Supplementary File S1). A positive IMS result was considered to be a capture of five or more colony-forming units (CFUs). All experiments involving the use of animals were approved by the local Animal Care Committee under the guidelines of the Canadian Council on Animal Care.
Determination of capture efficiency for MAbs
L. monocytogenes strain LI0521 (serotype 4b), premixed with MAb hybridoma tissue culture fluid (TCF), was used to assess the capture efficiency of M3686, M3697, M3699, M3700, and M3644 with the IMS procedure, as described in Supplementary File S1.
Capture of various L. monocytogenes serotypes, other Listeria species, and non-Listeria bacteria with MAbs
One milliliter of bacterial cells at 104 cells/mL was mixed with 200 μL of MAb (M3686, M3697, M3699, and M3700) TCF diluted 1:100 in phosphate-buffered saline (PBS). As a negative control, M1169 specific for Campylobacter jejuni was used at 1:100 dilution. Capture was performed by IMS with a BeadRetriever™ as described in Supplementary File S1. Two technical replications were performed for each trial. Two trials were performed on two different days to generate an average number of cells captured by MAbs. Strains of L. monocytogenes and other bacterial species used in the experiments are listed in Table 1.
A positive IMS result was considered to be a capture of ≥5 CFUs. This threshold was determined based on IMS of Salmonella enterica serovar Typhimurium and Escherichia coli O157:H7 using the negative control MAb M1169 reactive against C. jejuni (Table 4). This experiment revealed a minimal nonspecific capture of up to 5 CFUs, when accounting the variability within one standard deviation.
Isolation of L. monocytogenes from a bacterial mixture by IMS and confirmation of captured bacterial cells by colony blot immunoassay
The ability of MAbs to isolate L. monocytogenes in the presence of L. innocua was evaluated with a bacterial mixture with ratios of L. monocytogenes to L. innocua at 1:10 and 1:100 by using IMS and confirmed by using a colony blot immunoassay (Hoszowski et al., 1996). The experimental procedure is described in detail in Supplementary File S1.
SPR analysis
The binding affinity of selected anti-LMOf2365_0148 and anti-LMOf2365_0638 MAbs toward corresponding antigens was determined by SPR analysis using purified rLMOf2365_0148, rLMOf2365_0639 and Fab fragments. Purification of rLMOf2365_0148 and rLMOf2365_0639 from the E. coli expression host was carried out as previously described (Zhang et al., 2016). Preparation of Fab fragments and SPR analysis of Fab-antigen interactions are described in Supplementary File S1.
Epitope mapping
A total of seven overlapping, similarly sized polypeptides of surface protein LMOf2365_0148 were designed, generated in a recombinant form by PCR cloning of the corresponding coding sequences (Supplementary Table S1), and used for epitope mapping (Supplementary Fig. S4). In L. monocytogenes, the LMOf2365_0148 protein presumably undergoes enzymatic processes to cleave off the N-terminal signal peptide during protein secretion (predicted by Signal P version 5.0) and to cleave the C-terminal LPXTG motif for surface anchoring. The overlapping polypeptides span the mature form of LMOf2365_0148 with polypeptide 7 designed to narrow the location of the epitope for a particular MAb M3692. Production of the polypeptides and their application to epitope mapping by Western blotting are presented in Supplementary File S1.
Results
Selection of the MAbs most suitable for L. monocytogenes isolation
Screening of MAbs previously raised against both rLMOf2365_0639 (Zhang et al., 2016) and rLMOf236_0148 (Zhang et al., 2022) proteins demonstrated that 1 out of 35 MAbs (M3644) against rLMOf2365_0639 and 16 out of 24 MAbs against rLMOf2365_0148 had the ability to capture L. monocytogenes by IMS (results not shown). Surprisingly, MAb M3692, one of the five MAbs to rLMOf2365_0148 (M3686, M3692, M3697, M3699, and M3700) previously demonstrated to react with a range of L. monocytogenes serotypes belonging to lineages I, II, and III and other Listeria species grown under nonselective and selective enrichment culture conditions (Zhang et al., 2022), failed to capture live L. monocytogenes cells. Four of the 16 MAbs to rLMOf2365_0148 (M3686, M3697, M3699, and M3700) that captured the most cells and MAb M3644 against rLMOf2365_0639 were selected for assessing the limits of capture and capture efficiencies.
The assessment was performed with indirect IMS in which bacterial cells are incubated with antibodies before addition of beads coated with anti-mouse secondary antibodies. MAbs in the TCF were used. To avoid cell loss due to washing, the free antibody was not removed from the cell suspension once mixed with the cells. Theoretically, both insufficient and excessive amounts of MAb can reduce capture capacity. An excess of free MAb can compete with MAbs in complex with the pathogen for binding to secondary anti-mouse antibodies coated on beads. Hence, a series of different TCF dilutions (1:200, 1:100, 1:50, 1:10, 1:5, and 1:2) were tested to find the dilution that resulted in the most cells being captured. Dilutions of 1:10 for M3644 and of 1:100 for M3686, M3697, M3699, and M3700 were found to be optimal for IMS.
Table 2 shows the results of capturing live cells of L. monocytogenes strain LI0521 (serotype 4b) by IMS. MAb M3644 against rLMOf2365_0639 had a capture limit of 104 cells/mL with a capture efficiency of 0.2%. MAbs M3686, M3697, M3699, and M3700 against rLMOf2365_0148 had a capture limit of 103 cells/mL. Capture efficiencies for M3686, M3697, M3699, and M3700 were 3.8%, 2.7%, 1.7%, and 2.3%, respectively, at 103 cells/mL (Table 2). With a concentration of 104 cells/mL, similar capture efficiencies of 3.7%, 2.6%, 2.0%, and 2.8% were observed for M3686, M3697, M3699, and M3700, respectively (Table 2). M3686, M3697, M3699, and M3700 had the highest capture efficiencies and the lowest capture limits and were thus selected for further evaluation.
Efficiency of Capturing Listeria monocytogenes at Various Input Cell Numbers by Immunomagnetic Separation with Five Selected Monoclonal Antibodies
Capture was performed using L. monocytogenes strain LI0521 (serotype 4b) at three different cell numbers in 1 mL of PBS containing one of the MAbs: M3644 against rLMOf2365_0639 and M3686, M3697, M3699, and M3700 against rLMOf2365_0148. The irrelevant MAb M1169 against Campylobacter jejuni was used as a negative control. Three independent experiments with two replicates each time were performed to obtain the average number of colonies captured ± standard deviation. A positive result in IMS is ≥5 CFUs captured.
CFU, colony-forming unit; IMS, immunomagnetic separation; MAb, monoclonal antibody; PBS, phosphate-buffered saline.
Application of IMS with selected MAbs to capturing various serotypes of three L. monocytogenes lineages
Serotypes of L. monocytogenes lineages I (1/2b, 4b, and 4d), II (1/2a, 1/2c, and 3a), and III (4a) are often involved in listeriosis. Development of the IMS capability to capture serotypes from the three lineages (I, II, and III) is particularly important in the identification of food and environmental sources contaminated with L. monocytogenes. MAbs M3686, M3697, M3699, and M3700, selected earlier with respect to the capture efficiencies and capture limit for serotype 4b (lineage I), were evaluated in IMS for capturing the cells of serotypes 1/2b and 4d (lineage I), serotypes 1/2a, 1/2c, and 3a (lineage II), and serotypes 4a (lineage III) (Table 3), with one strain for each serotype tested. MAbs 3686 and M3700 captured cells of all the L. monocytogenes serotypes tested. M3697 captured L. monocytogenes serotype 4a and 1/2b cells. M3699 only captured L. monocytogenes serotype 4a cells.
Capture of Various Listeria monocytogenes Serotypes by Selected Monoclonal Antibodies
Six additional serotypes (1/2a, 1/2b, 1/2c, 3a, 4a, and 4d) besides serotype 4b of L. monocytogenes were used to assess the capture ability of M3686, M3697, M3699, and M3700. Capture was performed using 104 cells in 1 mL of PBS. The irrelevant MAb M1169 against Campylobacter jejuni was used as a negative control. Two independent experiments with two replicates each time were performed to obtain the average number of colonies captured ± standard deviation. See Table 1 for information about the isolates used. A positive result in IMS is ≥5 CFUs captured. The data for serotype 4b are shown in Table 2.
CFU, colony-forming unit; IMS, immunomagnetic separation; MAb, monoclonal antibody; PBS, phosphate-buffered saline.
The capture specificity of selected MAbs
The selective enrichment culture, routinely used to increase the cell number of L. monocytogenes in test samples before the isolation and detection, can enrich all members of the Listeria genus. It is thus crucial to have MAbs specific for L. monocytogenes for use in IMS to isolate the target pathogen from selective enrichment culture and facilitate downstream detection. IMS of bacteria from 104 cells in 1 mL of PBS with four selected MAbs M3686, M3697, M3699, and M3700 showed that these MAbs were unable to capture cells of Listeria seeligeri, L. innocua, Listeria ivanovii, Listeria grayi, Listeria welshimeri, Salmonella Typhimurium, and E. coli O157:H7 with the exception that M3697 and M3700 were able to capture L. welshimeri cells (Table 4).
Evaluation of Monoclonal Antibodies M3686, M3697, M3699, and M3700 for Capture Specificity Using Non-Listeria monocytogenes Bacterial Species
Capture was performed using 104 cell in 1 mL of PBS. The irrelevant MAb M1169 against Campylobacter jejuni was used as a negative control. Two independent experiments with two replicates each time were performed to obtain the average number of colonies captured ± standard deviation. A positive result in IMS is ≥5 CFUs captured.
CFU, colony-forming unit; IMS, immunomagnetic separation; MAb, monoclonal antibody; PBS, phosphate-buffered saline.
Targeted isolation of L. monocytogenes from a mixed bacterial population
The ability of IMS with MAbs M3686, M3697, M3699, and M3700 to separate the target pathogen from nontarget cells was assessed by using L. monocytogenes mixed with excessive nontarget L. innocua cells at ratios of 1:10 and 1:100. Figure 1 shows an example of using colony blot immunoassays, with colony detection by an L. monocytogenes-specific polyclonal antibody (PAbs to rLMOf2365_0639), to determine the proportion of L. monocytogenes cells recovered by IMS from a mixture of L. monocytogenes and L. innocua cells. Comparison of the colony counts on brain–heart infusion broth (BHI) agar plate and colony blot revealed that nearly all the captured cells were L. monocytogenes cells.

A colony blot immunoassay for detecting Listeria monocytogenes isolated by IMS using MAb M3697 from a mixture of L. monocytogenes and Listeria innocua. One milliliter of L. monocytogenes strain LI0521 (5 × 103 cells) mixed with L. innocua (5 × 104 cells) at a ratio of 1:10 was incubated with M3697 for IMS. Captured cells were plated on BHI agar
The specificity of colony blot immunoassay for L. monocytogenes was demonstrated by plating ∼100 CFUs of L. innocua cells and then detecting similarly by colony blot immunoassay. Almost no L. innocua colonies were detected (Fig. 1). From a mixed suspension of L. monocytogenes and L. innocua at a ratio of 1:10, 75.0%, 73.3%, 54.4%, and 82.8% of the cells captured, respectively, by MAbs M3686, M3697, M3699, and M3700 were L. monocytogenes; at a ratio of 1:100 (the cell number of L. monocytogenes to L. innocua), 52.3%, 72.3%, 63.6%, and 75.1% of the cells captured were L. monocytogenes.
A small proportion of colonies at the edge of plates were not lifted onto the membrane for colony blot immunoassay due to the smaller size of the membrane. Therefore, the percentage of L. monocytogenes in the total cells captured may be underestimated. Nonetheless, the proportions of captured L. monocytogenes cells were well above 10% and 1% of L. monocytogenes present in the initial suspension of mixed bacteria.
Capture of L. monocytogenes cultured in selective enrichment media
L. monocytogenes that may exist at a trace quantity in food and environmental samples must be enriched by proliferation in selective enrichment cultures to detectable levels before detection. Health Canada's standard selective enrichment culture methods for Listeria isolation and detection, described in MFHPB-07 (Warburton et al., 2011) and MFHPB-30 (Pagotto et al., 2011), involve primary and secondary enrichment steps. Both MFHPB-07 and MFHPB-30 methods are used to isolate L. monocytogenes and other Listeria species from food and environmental samples. Palcam and UVM1 (LEB) are the primary culture enrichment media of MFHPB-07 and MFHPB-30 methods, respectively. Both Palcam and UVM1 (LEB) are designed to inhibit the growth of Gram-negative bacteria and Gram-positive cocci. Palcam contains selective agents polymyxin-B, ceftazidime, acriflavine, and lithium chloride, whereas UVM1 (LEB) has nalidixic acid and acriflavine.
To expedite bacterial isolation and facilitate downstream pathogen detection, it is possible to capture L. monocytogenes from primary selective enrichment culture using IMS. L. monocytogenes strain LI0521 (serotype 4b), grown in primary selective enrichment culture of Palcam in MFHPB-07 or LEB/UVM1 in MFHPB-30, was used to test the IMS procedure. Preliminary experiments were conducted to optimize the IMS procedure for capturing L. monocytogenes from selective enrichment culture. Due to the variability associated with preparing 1 CFU (in 1 mL) of L. monocytogenes for inoculation into 224 mL of Palcam or LEB/UVM1, ∼5 CFUs were used for inoculation. At an inoculum of 5 CFUs in 224 mL of Palcam medium, a concentration of 105–106 CFU/mL was reached after incubation at 35°C for 20–21 h.
Similarly, at an inoculum of 5 CFUs in 224 mL of UVM1/LEB medium, a cell concentration of 105 CFU/mL was reached after incubation at 30°C for 30 h. A dilution of the enrichment culture in PBS is necessary to minimize or eliminate possible inhibitory effects of salts in selective enrichment culture on IMS. Experiments with 2-, 5-, 10-, and 100-fold culture dilutions demonstrated that a 5-fold dilution resulted in capturing the most cells by IMS; therefore, this dilution was adopted for subsequent experiments. Table 5 shows the number of L. monocytogenes cells captured by IMS with M3686, M3697, M3699, and M3700 from fivefold diluted enrichment cultures. The minimum culture enrichment time for the IMS method described in this study is 20–21 h in Palcam medium and 30 h in UVM1/LEB medium.
Capture of Listeria monocytogenes After Culturing in Selective Enrichment Media by Immunomagnetic Separation with Monoclonal Antibodies M3686, M3697, M3699, and M3700
Approximately 5 CFUs of L. monocytogenes (strain LI0521 serotype 4b) were inoculated in 224 mL of either Palcam (primary selective enrichment broth for MFHPB-07) or LEB/UVM1 (primary selective enrichment broth for MFHPB-30) and incubated for 20–21 or 30 h, respectively. One milliliter of fivefold diluted enrichment culture in PBS was used in capture experiments. The average number of colonies captured ± standard deviation was obtained from two IMS experiments from two cultures inoculated together. The irrelevant MAb M1169 against Campylobacter jejuni was used as a negative control. A positive result in IMS is ≥5 CFUs captured.
CFU, colony-forming unit; IMS, immunomagnetic separation; MAb, monoclonal antibody; PBS, phosphate-buffered saline.
Under these experimental conditions, each of the four selected MAbs captured a greater number of L. monocytogenes cells from the Palcam enrichment culture than that from the UVM1/LEB culture (Table 5).
Determination of MAb affinity
The affinity of an MAb for interacting with the antigen's epitope plays an important role in the capture of the target pathogen using IMS. It is quantitated by measuring the equilibrium dissociation constants (K D) of an antibody–antigen interaction. The MAbs selected for K D determination by SPR were M3686, M3697, M3699, M3700, M3644, and M3692. Of the selected MAbs, M3686, M3697, M3699, and M3700 were shown above to have the ability to capture L. monocytogenes, whereas M3692 did not show the ability to capture L. monocytogenes. M3644 was the only MAb against LMOf2365_0639 capable of capturing L. monocytogenes.
The IgG of each MAb was affinity purified from TCF and used to prepare Fab fragments for SPR analysis. M3686, M3692, M3697, M3699, and M3700 IgGs were purified by affinity chromatography using a column of LMOf2365_0148-conjugated agarose; M3644 IgG was purified using a column of protein L-conjugated agarose as protein L binds ĸ light chain of M3644, but has no affinity for bovine IgG in the TCF. Supplementary Figure S1 shows sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis of purified M3697 IgG as an example. Following fragmentation of purified IgG by digestion, Fab fragments recovered in the flow-through from a commercial protein A spin column, and undigested IgG or Fc fragments eluted from the protein A column were analyzed for the purification of Fab fragments by SDS-PAGE and Western blotting analysis.
Being shown as an example in Supplementary Figure S2, Western blots using specific antibodies detected no Fc fragments, but only Fab fragments in the Fab preparation derived from MAb M3697 IgG. Immediately before SPR analysis, only peak fractions containing the Fab fragments from size exclusion chromatography of the Fab preparation were collected for the SPR analysis (Supplementary Fig. S3).
SPR analysis of binding of Fab fragments to the target antigen was performed by flowing the Fab fragments in a range of concentrations over the sensor chip immobilized antigen. Figure 2 shows the kinetics of interaction between the Fab fragments and the antigen in real time, as represented by SPR sensorgrams (plots of resonance units vs. time). All the binding data sets show good fitting to the 1:1 binding model (Fig. 2). All six MAbs had high affinities as K D was in the nanomolar range (Table 6). Interestingly, MAb M3692 has the highest affinity (lowest K D at 0.9 nM) but was incapable of capturing L. monocytogenes.
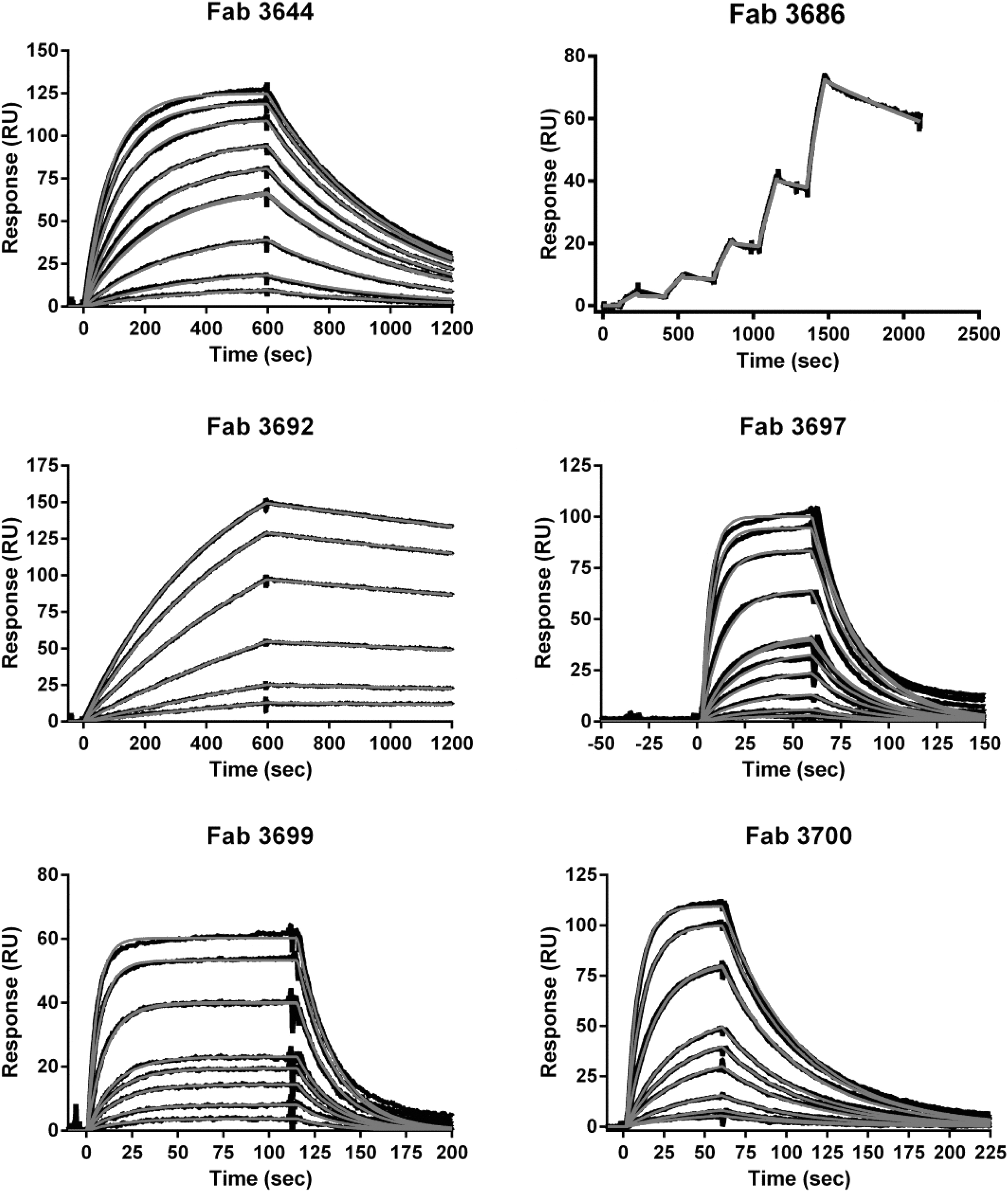
Kinetic analysis of binding of Fab fragments derived from M3644, M3686, M3692, M3697, M3699, and M3700 to the corresponding antigen with SPR. SPR sensorgrams show Fab fragments binding to immobilized antigens at: 0.5, 1, 2.5, 5, 5, 7.5, 10, 15, 20, and 25 nM for M3644, 1.25, 2.5, 5, 10, and 20 nM for M3686, 0.5, 1, 2.5, 5, 7.5, 10, and 10 nM for M3692, 0.5, 1, 2.5, 5, 7.5, 10, 10, 25, 50, 75, and 100 nM for M3697, 1, 2.5, 5, 7.5, 10, 25, 25, 50, and 75 nM for M3699, and 0.5, 1, 2.5, 5, 7.5, 10, 25, 25, 50, and 75 nM for M3700. For kinetic analysis of M3644, rLMOf2365_0639 was used; for kinetic analysis of M3686, M3692, M3697, M3699, and M3700, rLMOf2365_0148 was used. Red lines represent fitted curve and black line represents raw measurements. Binding kinetics of Fab fragments derived from M3686 was analyzed using single cycle kinetics, which involves five subsequent injections of increasing concentrations followed by a dissociation of 10 min. RU, resonance unit; SPR, surface plasmon resonance.
Association (k a) and Dissociation (k d) Rate Constants and Equilibrium Dissociation (K D) Constants for M3644, M3686, M3692, M3697, M3699, and M3700
MAb, monoclonal antibody.
Epitope mapping of five MAbs against the LMOf2365_0148 protein
Seven overlapping recombinant polypeptides that covered the length of mature LMOf2365_0148 protein were used to map the epitopes of five MAbs raised against rLMOf2365_0148, M3686, M3692, M3697, M3699, and M3700. Epitope mapping may predict the specificity of MAbs for L. monocytogenes strains and provide an explanation for the observation that certain MAbs of high affinity for its antigen such as M3692 fail to capture L. monocytogenes cells. Of the five MAbs examined, only M3692 was found to recognize an epitope in a region overlapped by peptides 6 and 7 and close to the C-terminal region of the LMOf2365_0148 protein (Supplementary Fig. S4). Western blots with the other four MAbs (M3686, M3697, M3699, and M3700) detected the denatured full-length LMOf2365_0148 protein but not any polypeptides derived from LMOf2365_0148, which were shown to be expressed in E. coli by the detection with PAbs (Supplementary Fig. S4).
Discussion
The present work investigated MAbs against surface proteins rLMOf2365_0639 (Zhang et al., 2016) and rLMOf2365_0148 (Zhang et al., 2022), described previously, for their ability to capture live L. monocytogenes cells. The affinity of selected MAbs was measured quantitatively for their respective antigens by SPR analysis so to provide a possible explanation for the capture ability observed. The epitopes of these MAbs were mapped to gain an insight into the binding specificity for L. monocytogenes strains.
Of the 35 MAbs against rLMOf2365_0639 examined, only M3644 exhibited the ability to capture L. monocytogenes although a previous study has shown that many have the potential for detecting L. monocytogenes by enzyme-linked immunosorbent assay (ELISA) (Zhang et al., 2016). Sixteen out of the 24 MAbs previously developed against rLMOf2365_0148 (Zhang et al., 2022) captured live L. monocytogenes with M3686, M3697, M3699, and M3700 and exhibited the best capture efficiencies. At the optimal working dilution of TCF, M3644 against rLMOf2365_0639 captured fewer cells, compared with the four MAbs (M3686, M3697, M3699, and M3700) against rLMOf2365_0148.
The number of cells captured by M3644 was consistently one order of magnitude less than the four MAbs against rLMOf2365_0148 regardless of the dilution of the TCF used (Table 2). The reduced capture ability of M3644 appeared to be unrelated to the affinity of M3644 because the dissociation constant of M3644 was comparable with those of M3686, M3697, M3699, and M3700 (Table 6). In addition to the importance of antibody affinity in antigen binding, the avidity of these MAbs likely contributes considerably to their capture ability.
Previous immunofluorescence microscopy results showed that the epitopes recognized by MAbs against rLMOf2365_0639 were surface exposed on live cells (Zhang et al., 2016). The reduced ability or inability of MAbs to capture L. monocytogenes cells expressing LMOf2365_0639 could be due to the characteristics of this protein on the bacterial cell surface rather than affinity of the MAbs. One possible explanation is that LMOf2365_0639 is expressed at a low level compared with LMOf2365_0148. In addition, the distribution of the LMOf2365_0639 on the cell surface may contribute to the reduced or no capture ability of the corresponding MAbs. Possible diffused localization of the LMOf2365_0639 protein on the cell surface may result in a decreased avidity and thereby a decreased capture of target bacteria.
Epitope mapping of M3686, M3692, M3697, M3699, and M3700 to LMOf2365_0148 by Western blot analysis of overlapping polypeptide fragments yielded results only for M3692 (Supplementary Fig. S4). Interestingly, the epitope of M3692 is located at the region of LMOf2365_0148 close to the LPXTG motif at the C-terminus via which the protein is presumably anchored to the cell wall peptidoglycan. Although M3692 had the highest affinity with a K D of 8.87 × 10−10 M among all the MAbs analyzed by SPR, it was not able to capture live L. monocytogenes cells by IMS. The location of the M3692 epitope close to the protein anchor motif may interfere with binding of the MAb to its epitope and thus affects capture of live cells.
The ability of M3692 to detect formalin-killed cells as revealed by indirect ELISA and live cells as revealed by immunofluorescence microscopy (Zhang et al., 2022) suggested that its epitope, to some degree, was surface exposed. Peptides of LMOf2365_0148 identified from the proteomics study of surface exposed proteins (Zhang, 2015) revealed a much higher coverage in the N-terminal region of the protein than its C-terminal region (Supplementary Fig. S5). It is conceivable that the C-terminal region of LMOf2365_0148 was less surface exposed and thus less accessible to antibodies. Other researchers have noted the importance of epitope surface exposure in pathogen detection. Lack of epitope surface exposure has been reported to limit the binding of MAbs to Bacillus anthracis spores (Nuttall et al., 2011). Given challenges such as weak association of antibody-coated beads to cells (Hudson et al., 2001), any potential limitations may abrogate the capture ability of an MAb.
M3686, M3697, M3699, and M3700 did not react to overlapping large polypeptides (∼16 kDa) of LMOf2365_0148 used for epitope mapping. However, these MAbs detected the denatured full-length LMOf2365_0148 protein on Western blots, suggesting that they recognized a linear epitope. Lack of the reactivity of these MAbs to other overlapping polypeptides did not appear to be consistent with these antibodies recognizing linear epitopes.
The capture experiments revealed the greatest capture efficiency for M3686 in comparison with M3697, M3699, and M3700 (Table 2). This finding is consistent with the measured dissociation constants of the antibody–antigen interaction. The dissociation constant for M3686 (1.41 × 10−9 M) is one order of magnitude lower than those for M3697, M3699, and M3700 (Table 6). Antibody affinity plays an important role in capture efficiency as many reports have demonstrated the importance of antibody affinity in immunoassay sensitivities (Liang et al., 2007; Peng et al., 2014; Steward and Lew, 1985). However, additional factors may also play a role in capture efficiency, as noted in this study in the case of MAb M3644 with a dissociation constant of 6.30 × 10−9 M showing a low capture efficiency.
Extensive IMS analysis of the four MAbs M3686, M3697, M3699, and M3700 using strains of several L. monocytogenes serotypes, other Listeria species, and other non-Listeria bacteria revealed the capture specificity of these MAbs. M3686 was specific for capturing L. monocytogenes as it was incapable of capturing selected strains of L. seeligeri, L. innocua, L. ivanovii, L. grayi, L. welshimeri, E. coli O157:H7, and Salmonella Typhimurium. Moreover, the four MAbs were able to selectively capture L. monocytogenes from a mixed suspension of L. monocytogenes and L. innocua with 10 or 100 times more L. innocua cells than L. monocytogenes cells.
The ability to enrich the target pathogen from a sample containing other bacteria is an important quality of an MAb in IMS, because it allows separation of the target from high background microflora that can produce false-negative results in PCR detection (Hsih and Tsen, 2001). The high specificity of M3686 is encouraging as the use of this MAb in IMS will facilitate the isolation of L. monocytogenes from an enrichment culture, which likely contains a high background of other Listeria species (Besse et al., 2010; Beumer and Hazeleger, 2003; Oravcova et al., 2008).
Previous studies have reported the use of MAbs in the IMS for Listeria species (Garrec et al., 2003; Hsih and Tsen, 2001; Hudson et al., 2001; Kaclikova et al., 2001; Rijpens and Herman, 2004; Skjerve et al., 1990; Uyttendaele et al., 2000). The MAbs that were previously shown to have high capture efficiencies for L. monocytogenes also recovered a considerable quantity of other Listeria species. Paoli et al. (2007) used IMS to isolate L. monocytogenes grown in BHI. These researchers found high capture efficiencies of 17.34–1.38% for the L. monocytogenes strains tested, but a significant capture of other Listeria species (500–10 CFU counts with an input of 105 cells for either L. monocytogenes or other Listeria species).
Mendonca et al. reported an MAb with capture efficiencies of 49.2% and 32.2% for L. monocytogenes and L. ivanovii, respectively (Mendonca et al., 2012). They observed a significant capture of L. innocua and Listeria marthii (480 and 400 CFUs, respectively) at an input of 2 × 104 cells under the same conditions. In the present investigation, the capture efficiencies for L. monocytogenes with M3686, M3697, M3699, and M3700 were 3.7%, 2.6%, 2.0%, and 2.8%, respectively, with an input of 104 cells. The capture efficiencies observed in Table 2 are lower than those observed in other studies (Mendonca et al., 2012; Paoli et al., 2007). For the purpose of obtaining isolated colonies for subsequent molecular detection, the capture efficiencies of MAbs generated here were sufficient as 200–370 CFUs were captured.
Importantly, M3686 is highly specific for capturing L. monocytogenes cells by IMS. No CFUs were recovered by M3686 using 104 cells for each of L. seeligeri, L. innocua, L. ivanovii, L. grayi, and L. welshimeri; <3 CFUs were isolated using 104 cells for each of E. coli O157:H7 and Salmonella Typhimurium. Both specificity and capture efficiency are equally important when developing MAbs for L. monocytogenes isolation. Capture efficiencies are not only dependent on the quality such as affinity of the antibodies used but also on the level of the surface antigens expressed as well as many aspects of the experimental procedure.
The experimental aspects include initial cell concentration, the size and concentration of the immunomagnetic beads, as well as the number of wash steps after pathogen capture (Hudson et al., 2001; Mendonca et al., 2012). Therefore, experimental details should be considered when attempting to make a meaningful comparison of the results between studies.
M3686 exhibited the ability to capture cells of seven L. monocytogenes serotypes 1/2a, 1/2b, 1/2c, 3a, 4b, 4a, and 4d. However, this MAb captured 10 times L. monocytogenes serotype 4b cells compared with cells of other serotypes. Given that M3686 was raised against the LMOf2365_0148 derived from the serotype 4b, it may be speculated that the epitope recognized by M3686 has some sequence variations in other serotypes. The difference in capture efficiency between serotypes has been previously reported (Paoli et al., 2007). The MAbs developed here appear to be suitable for capturing the majority of L. monocytogenes serotypes at a cell concentration of 104 CFU/mL.
In addition to evaluating the ability of M3686, M3697, M3699, and M3700 to capture L. monocytogenes grown in nonselective BHI medium, this study also tested the ability of the selected MAbs to capture L. monocytogenes grown in primary selective enrichment cultures according to the MFHPB-30 and MFHPB-07 methods. All four MAbs were capable of capturing L. monocytogenes after abbreviated 20–21 and 30 h in primary selective enrichment culture according to MFHPB-30 and MFHPB-07, respectively (Table 5). While some antibodies captured L. monocytogenes cultured in BHI but did not react to L. monocytogenes grown in selective enrichment cultures (Paoli and Brewster, 2007), the ability of the MAbs tested here to capture L. monocytogenes grown in enrichment culture makes them applicable to practical diagnostic testing.
The IMS method reported in the present study was successfully adapted to the BeadRetriever platform to streamline multiple IMS testing steps and greatly increase the test efficiency. Moreover, metabolomic-based methods have been investigated for the identification of L. monocytogenes in food samples (Feng et al., 2022; Li et al., 2021). Bacterial isolation methods such as the IMS procedure reported herein can facilitate a downstream metabolomic analysis.
In conclusion, the application of the selected MAbs against the previously discovered surface protein biomarkers (Zhang et al., 2022; Zhang et al., 2016) to capturing L. monocytogenes was successful. The results showed that MAbs M3686, M3697, M3699, and M3700 were capable of capturing L. monocytogenes by IMS after a shortened time of primary selective culture enrichment according to both MFHB-07 and MFHP-30 methods. Particularly, one of the selected MAbs, M3686, specifically captured seven serotypes of L. monocytogenes from three lineages. While these MAbs were capable of capturing significant numbers of L. monocytogenes serotype 4b cells, they were also able to capture cells of other serotypes in reduced numbers.
Further research is needed to assess the capture of L. monocytogenes of various serotypes using the IMS method developed in this study in naturally or artificially contaminated food samples after 20–30 h of selective culture enrichment. Nonetheless, the results of the present study are an encouraging step toward the development of an automated IMS method applicable for routine isolation of L. monocytogenes from food and environmental samples.
Footnotes
Acknowledgments
The authors would like to thank Drs. C. Fehlner-Gardiner and O. Andrievskaia for their insightful and constructive comments on the article. They acknowledge the assistance from the Monoclonal Antibody Unit (C. Elmgren, K. Arnold, and J. Widdison) at the Canadian Food Inspection Agency (CFIA) in the production of monoclonal antibodies.
Authors' Contributions
C.X.Y.Z.: Investigation, methodology, validation, visualization, writing—original draft, review, and editing. H.D.: Investigation and validation. H.v.F.: Investigation, validation, and visualization. B.W.B.: Resources, methodology, and writing—review and editing. H.H.: Resources, methodology, and writing—review and editing. M.L.: Conceptualization, funding acquisition, methodology, project administration, resources, supervision, and writing—review and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the funding of the CFIA Research Partnership Strategies program [Project ID: OLF (1011) 01].
Supplementary Material
Supplementary File S1
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.