
Abstract
Staphylococcus aureus is a common foodborne pathogen and spoilage bacterium in meat products. To develop a natural preservative for meat products, this study revealed the antibacterial activity and mechanism of Rosa roxburghii Tratt pomace crude extract (RRPCE) against S. aureus, and applied RRPCE to the preservation of cooked beef. The diameter of inhibition zone, minimum inhibitory concentration (MIC), and minimum bactericide concentration of RRPCE against S. aureus were 15.85 ± 0.35 to 16.21 ± 0.29 mm, 1.5 mg/mL, and 3 mg/mL, respectively. The growth curve of S. aureus was completely stalled by treatment with RRPCE at 2 MIC. RRPCE results in the decrease of intracellular adenosine 5′-triphosphate (ATP) content, depolarization of cell membrane, leakage of cell fluid including nucleic acid and protein, and destruction of cell membrane integrity and cell morphology. During storage, RRPCE significantly reduced S. aureus viable counts, pH, and total volatile basic nitrogen of cooked beef compared with untreated samples (p < 0.05). In addition, RRPCE could significantly increase the redness (a*) value, decrease lightness (L*) and yellowness (b*) values, and slow down the color change of cooked beef (p < 0.05). These findings suggest that RRPCE can effectively inhibit S. aureus, and has the potential as a natural preservative for the preservation of cooked beef.
Introduction
S
In recent years, the prevalence of S. aureus in cooked meat products has been reported constantly, which indicates that this pathogen has brought great trouble to the development of meat industry and the health of consumers (Liu et al., 2020). Currently, adding chemical preservatives is the most commonly used method to inhibit the growth of S. aureus in cooked meat products and prolong the product shelf life (Fei et al., 2018). However, with the improvement of consumers' health awareness, they are not willing to accept chemical preservatives, because long-term consumption of chemical preservatives may cause potential harm such as carcinogenesis and teratogenesis (Fei et al., 2019; Guo et al., 2020a). Therefore, a green, safe, effective, and side effect-free natural preservative should be developed and used in the preservation of cooked meat products.
Rosa roxburghii Tratt pomace is a byproduct of R. roxburghii processing. At present, the main processing mode of R. roxburghii is juicing, resulting in about 40–50% residue of fruit pomace (Peng et al., 2017). Most of the treatment methods of R. roxburghii pomace are directly discarded, however, the discarded R. roxburghii pomace is easy to become moldy in the process of stacking, which leads to the waste of resources and environmental pollution (Meng et al., 2017). It can be seen that how to make full use of R. roxburghii pomace crude extract (RRPCE) resources has become an urgent problem to be solved.
Previous studies have found that R. roxburghii pomace is rich in a variety of active substances with strong antibacterial activity, including polyphenols, flavonoids, polysaccharide, organic acids, terpenoids, and cranberry anthocyanin (CA), suggesting the potential of R. roxburghii pomace as a natural preservative (Huang et al., 2022; Zhou and Fan, 2021). Therefore, we consider to further study the antibacterial pattern of RRPCE against S. aureus and use it as a natural preservative in cooked beef, thus reducing the use of chemical preservatives and avoiding the waste of R. roxburghii pomace.
With this background, the objectives of this study include the following: (1) to evaluate the antibacterial activity of RRPCE against S. aureus by measuring the diameter of inhibition zone (DIZ), minimum inhibitory concentration (MIC), minimum bactericide concentration (MBC), and growth curve; (2) to reveal the possible antibacterial pattern by analyzing the effects of RRPCE on intracellular adenosine 5′-triphosphate (ATP) content, membrane potential, leakage of nucleic acid and protein, cell membrane integrity, and cell morphology of S. aureus; and (3) to clarify the potential of RRPCE as a natural preservative by measuring the changes in a number of S. aureus viable bacteria, pH, color, and total volatile basic nitrogen (TVB-N) of cooked beef after treatment with RRPCE during storage.
Materials and Methods
Strains and culture conditions
Details of the eight S. aureus strains used in this study are as follows: S. aureus ATCC 13565 and S. aureus ACTT 25923 were purchased from the American Type Culture Collection (ATCC, Manassas, VA); The other six S. aureus strains were all isolated from cooked beef samples by the microbiology laboratory of Institute of Integrated Agricultural Development Research, Guizhou Academy of Agricultural Sciences. All the eight strains were used in the evaluation tests of antibacterial activity of RRPCE, and only S. aureus ATCC 13565 was used in the experiments of antibacterial mode of RRPCE and the application of RRPCE as a natural preservative.
Before the experiment, stock culture was streaked on Baird–Parker agar (BPA) containing egg yolk tellurite emulsion followed by an incubation at 37°C for 24 h. Then, the culture was streaked on BPA and incubated at 37°C for 24 h to obtain isolated colonies. Then, a typical gray-black colony was inoculated into the Luria-Bertani (LB) broth and incubated at 37°C for 20 h for later experiments.
Preparation of RRPCE
RRPCE powder was prepared according to the method reported by Chang et al. (2023). Briefly, after R. roxburghii is juiced, the pomace was fully dissolved in sterile distilled water at a ratio of 1:10 and then fully extracted at 65 °C for 2 h. After the filtrate was removed, the remaining residue was extracted two more times using the same method, and then, all the filtrate was transferred to a rotary evaporator (Zhejiang Nader Scientific Instrument Co. Ltd., Hangzhou, China) and treated at 40 °C for 22 h. The above RRPCE viscous liquid was freeze-dried at −80°C for 22 h to obtain RRPCE powder. The images of R. roxburghii, R. roxburghii pomace, and RRPCE powder are shown in Supplementary Figure S1, and the chemical composition of RRPCE is shown in Supplementary Table S1.
Determination of DIZ
According to the study of Chang et al. (2021), the DIZ of RRPCE against S. aureus was measured by the Oxford cup method. A hundred microliters of S. aureus suspension (approximately 107 colony-forming unit [CFU]/mL) was evenly spread onto the BPA plates supplemented with egg yolk tellurite emulsion, and three sterile 8-mm-diameter Oxford cups were placed on each plate followed by a gentle press. Then 200 μL of RRPCE aqueous solution (10 mg/mL) was added into each Oxford cup followed by a cultivation at 37°C for 24 h. Finally, the DIZ values were measured and recorded.
Determination of MIC and MBC
The MIC and MBC values of RRPCE against S. aureus were determined as described in the study by Fei et al. (2018). RRPCE was added to sterile warm BPA medium and the final concentrations were adjusted to 0 (Control), 0.375, 0.75, 1.5, 3, and 6 mg/mL, respectively, and then the systems were added to sterile 24-well plates. After the medium solidified, 2 μL of S. aureus suspension (106 CFU/mL) was added onto the center of each well and incubated at 37°C for 24 h. The minimum concentration at which the strain did not grow at all was the MIC of RRPCE against S. aureus. S. aureus was treated with RRPCE (≥1 MIC) for 30 min and then enumerated on BPA. After an incubation at 37°C for 24 h, the minimum concentration at which no colonies grew was the MBC of RRPCE against S. aureus.
Growth curve
The effect of RRPCE on the growth curve of S. aureus ATCC 13565 was plotted according to Kang et al. (2022), with slight modifications. The tested strain was added to LB medium to achieve a final concentration of 106 CFU/mL. The equal volume of LB medium containing RRPCE was mixed with each of the above samples to make the concentration of RRPCE reach 0.5 MIC, 1 MIC, 1.5 MIC, and 2 MIC, respectively.
Untreated S. aureus LB medium was used as the control group. Finally, the above samples were detected by an automatic growth curve analyzer (FP-1100-C; OY Growth Curves Co., Ltd.) at 37°C for 24 h to obtain the growth curves.
Measurement of intracellular ATP concentration
According to the method of Fei et al. (2018), 2 mL of S. aureus ATCC 13565 suspension (108 CFU/mL) was prepared, and RRPCE was added into suspension to make the final concentration reach 1 MIC and 2 MIC, respectively. Untreated S. aureus suspension was used as the control group. After treatment at 37°C for 30 min, ATP was extracted from the cells using an ATP assay kit (Beyotime Bioengineering Institute, Shanghai, China), and then its relative luminescent units were determined using a multifunctional microplate reader (Jinan Qiansi Biotechnology Co. Ltd., Shandong, China).
Membrane potential analysis
According to the study described by Guo et al. (2019), RRPCE was added in the S. aureus ATCC 13565 bacterial suspension (107 CFU/mL) to adjust its concentration to 1 MIC and 2 MIC, respectively. Untreated S. aureus suspension was used as the control group. Then, 125 μL of the above samples and 0.5 μL of bis-(1, 3-dibutylbarbituriis)—(1,3-dibutylbarbituric acid) trimethine oxonol fluorescent probe were added to a black opaque 96-well plate. After cultivation at 37°C for 30 min, the fluorescence intensity of the samples was measured at an excitation wavelength of 492 nm and an emission wavelength of 515 nm using a multifunctional microplate reader (Jinan Qiansi Biotechnology Co. Ltd.).
Cell membrane integrity
The effect of RRPCE on cell membrane integrity of S. aureus ATCC 13565 was determined as described in a report by Xing et al. (2022a). RRPCE was added into the S. aureus suspension (108 CFU/mL), and the final concentrations were adjusted to 1 MIC and 2 MIC, respectively. Untreated S. aureus suspension was used as the control group. After incubation at 37°C for 3 h, S. aureus cell pellets were collected, washed, and resuspended in phosphate buffered saline (PBS) buffer (0.01 M, pH = 7.4). The samples were stained with propidium iodide (PI) dye and incubated in the dark for 30 min. Finally, 10 μL of the system was observed and photographed with an inverted fluorescence microscope [Thermo Fisher Technology (China) Co., Ltd., Shanghai, China] at an excitation wavelength of 536 nm and an emission wavelength of 617 nm.
Determination of efflux of nucleic acid and protein
According to previous studies (He et al., 2014; Ma et al., 2016), the efflux of nucleic acid and protein from S. aureus ATCC 13565 treated with different concentrations of RRPCE was determined. In brief, to determine the effect of RRPCE on nucleic acid leakage from the tested bacteria, 107 CFU/mL of bacterial solution was exposed to RRPCE at 0 MIC (Control), 0.5 MIC, 1 MIC, 1.5 MIC, and 2 MIC, respectively, and incubated at 37 °C for 12 h. Every 2 h, samples with different treatments were centrifuged at 15,000 × g for 10 min to obtain the supernatant. Then the optical density of the supernatant was measured at 260 nm using a multifunctional microplate reader (Jinan Qiansi Biotechnology Co. Ltd.).
Sterile normal saline with RRPCE of varying concentrations (0 MIC, 0.5 MIC, 1 MIC, 1.5 MIC, and 2 MIC) was used as control. Meanwhile, every 3 h, the samples treated with 0 MIC, 1 MIC, and 2MIC were centrifuged to obtain the supernatant, which was analyzed by the Coomassie brilliant blue method to determine the amount of protein released from tested cells.
Scanning electron microscopy and transmission electron microscopy analysis
According to previous reports (Fei et al., 2020; Xing et al., 2022a), the effects of RRPCE on the morphology of S. aureus ATCC 13565 cells were observed by scanning electron microscopy (SEM) and transmission electron microscopy (TEM). Briefly, S. aureus suspensions (107 CFU/mL) were treated with RRPCE at 0 MIC (Control), 1 MIC, and 2 MIC for 4 h and then centrifuged to obtain pellet. The pellet was fixed with 0.1 M sodium phosphate buffer containing 2.5% glutaraldehyde for 2 h, and then washed with PBS buffer three times. The samples were dehydrated in 30%, 50%, 70%, 90%, and 100% ethanol solutions for 15 min. Finally, after vacuum freeze-drying and gold spraying, the samples were observed under an SEM (TM3030; Hitachi, Tokyo, Japan). For TEM analysis, after glutaraldehyde fixation, S. aureus cells were fixed with 1% osmium tetroxide for 2 h.
The samples were washed with PBS three times and dehydrated as in the SEM procedure. Finally, the above samples were embedded by Epon Lx-112 and double-stained with uranyl acetate and lead citrate followed by an observation under TEM (H-7650; Hitachi).
Preparation of cooked beef
Using the method in the study of Gong et al. (2021), fresh beef was cut into small pieces weighing about 1.5 g followed by sterilization at 121°C for 15 min. The obtained sterile cooked beef samples with a mass of ∼1 g were soaked in RRPCE solutions at concentrations of 0 MIC (Control), 1 MIC, and 2 MIC for 10 s, respectively, and subsequently placed in Petri dishes for 30 min. Finally, the samples were sealed in sterile bags and stored at 4°C for later experiments.
Enumeration of S. aureus on cooked beef
On the basis of the method reported by Guo et al. (2020b), the cooked beef samples with different treatments were inoculated with S. aureus ATCC 13565 at a concentration of 103 CFU/mL and stored at 4°C. Samples were taken out on days 0, 3, 6, and 9, and thoroughly mixed with 9 mL of sterile normal saline. After proper gradient dilution, 100 μL of mixture was spread evenly on BPA (with egg yoke tellurite emulsion) plates, and then, colonies were counted after an incubation at 37°C for 24 h.
Measurement of pH
The pH value of cooked beef samples with different treatments was measured according to a previous report (Xing et al., 2022b). On days 0, 3, 6, and 9, each sample (1 g) was added into 9 mL of distilled water, and the pH value was measured using a pH meter (Sartorius, Gottingen, Germany) after homogenization treatment for 1 min.
Color analysis
On days 0, 3, 6, and 9, the lightness (L*), redness (a*), and yellowness (b*) of cooked beef samples with different treatments were determined using a CR-300 Chroma Meter (Minolta Co., Osaka, Japan) as previously reported (Gong et al., 2021). Finally, the total color difference (ΔE*) was calculated according to the following formula:
Determination of TVB-N
According to the method reported by Zhang et al. (2011), the changes in TVB-N values of cooked beef treated with different concentrations of RRPCE during storage were determined. On days 0, 3, 6, and 9, samples (1 g) were added to 10 mL of distilled water followed by beating, and filtrated after a 30-min stand. Then, 5 mL of filtrate and 5 mL of 1% magnesium oxide solution were added into the reaction chamber of a semimicro Kirschner apparatus (Nanjing Tenghui Experimental Technology Co., Ltd., Nanjing, China). After 5 min of distillation, the distillate was absorbed by 100 mL of ethanol solution containing 10 mL of boric acid solution and mixed indicator. Finally, the mixture was titrated with 0.01 mol/L of hydrochloric acid solution, and the TVB-N value was calculated based on the consumption of hydrochloric acid.
Statistical analysis
All experiments were repeated three times, and all data were expressed as mean ± standard deviation. The analysis of variance in the SPSS 20.0 software (SPSS, Inc., Chicago, IL) was used to calculate differences between treatment groups. p < 0.05 indicated significant differences among different treatments.
Results and Discussion
Antibacterial activity of RRPCE against S. aureus
The DIZs, MICs, and MBCs of RRPCE against eight strains of S. aureus are shown in Table 1. DIZ values ranged from 15.85 ± 0.35 mm to 16.21 ± 0.29 mm, and no significant differences in DIZs among different strains were found (p > 0.05). The MICs and MBCs of RRPCE against eight S. aureus strains were 1.5 and 3 mg/mL, respectively. Currently, the antibacterial activities of a number of natural extracts against S. aureus have been evaluated, for example, the MICs of CA, vine tea extract, Pyrrosia petiolosa ethyl acetate extract, Eucommia ulmoides male flower extract (EUMFE), and Amaranthus tricolor crude extract (ATCE) against S. aureus were 5, 6.3, 7.8, 40, and 80 mg/mL, respectively (Gong et al., 2021; Guo et al., 2020b; Liang et al., 2020; Song et al., 2017; Xing et al., 2022a). Compared with the above extracts, RRPCE, as a crude extract, showed better antibacterial activity against S. aureus.
Diameter of Inhibition Zone, Minimum Inhibitory Concentration, and Minimum Bactericide Concentration of Rosa roxburghii Tratt Pomace Crude Extract Against All Eight Staphylococcus aureus Strains
Values were expressed as mean ± SD of triplicate experiments. Different lowercase letters in the same column indicate significant differences between groups (p < 0.05).
ATCC, American Type Culture Collection; DIZ, diameter of inhibition zone; MBC, minimum bactericide concentration; MIC, minimum inhibitory concentration; SD, standard deviation.
Supplementary Table S1 shows that RRPCE contains polyphenols (24.7%), flavonoids (16.7%), organic acids (8.27%), anthocyanins (1.32%), and terpenoids (1.07%), which have been shown to have antibacterial activities (Gong et al., 2021; Tan et al., 2022; Yamaguchi, 2022; Zhang et al., 2020). In addition, Alvarez-Ordonez et al. (2014) found that organic acids reduced the intracellular pH of tested strains and thus accelerated irreversible damage to proteins and DNA. Li et al. (2016) reported that tea polyphenols combined with organic acids can inhibit the growth of foodborne pathogens more effectively, which may be one of the reasons for the RRPCE's strong antibacterial activity.
The effects of treatments with different concentrations of RRPCE on the growth curve of S. aureus ATCC 13565 are shown in Figure 1. Compared with the control group, the growth curves of S. aureus treated by RRPCE were obviously inhibited, and the inhibition effect was more obvious with the increase of RRPCE concentration. When the concentration of RRPCE was equal to or greater than 1 MIC, the logarithmic phase of the growth curve of S. aureus disappeared. When the concentration of RRPCE reached 2 MIC, S. aureus almost did not grow in the whole growth phase.

Growth curves of Staphylococcus aureus ATCC 13565 treated with different concentrations of RRPCE. ATCC, American Type Culture Collection; RRPCE, Rosa roxburghii Tratt pomace crude extract.
Similar results were found in the process of ATCE and EUMFE against S. aureus (Guo et al., 2020b; Xing et al., 2022a), Chrysanthemum bud crude extract (CBCE) against Cronobacter sakazakii and Salmonella Typhimurium (Chang et al., 2021; Xing et al., 2022b), and olive oil polyphenol extract (OOPE) against Listeria monocytogenes, Bacillus cereus, and S. aureus (Fei et al., 2019; Guo et al., 2020a; Guo et al., 2019).
Change in intracellular ATP levels
ATP can provide essential energy for normal physiological activities and proliferation of microorganisms, and so, the decrease of intracellular ATP content will directly lead to the arrest of microbial growth (Shi et al., 2016). Figure 2A shows the changes in intracellular ATP content of S. aureus ATCC 13565 after treatments with RRPCE. Compared with untreated strains, the intracellular ATP contents of S. aureus treated by RRPCE were significantly reduced (p < 0.05), and decreased with the increase of RRPCE concentration. Similarly, previous studies have shown that the intracellular ATP content of tested bacteria decreased sharply after exposure to natural extracts (Fei et al., 2019; Guo et al., 2020a; Xing et al., 2022b), which is consistent with our results. In addition, Fei et al. (2018) suggested that the direct causes of a significant reduction in the ATP content of tested cells were increased cell membrane permeability, impaired cell morphology, and excessive ATP hydrolysis.

Effect of RRPCE on
Change in membrane potential
Once normal cells are stimulated by external environment or substances, their membrane potential will change, which is considered to be an important indicator to judge whether cells are destroyed (Fei et al., 2020). Figure 2B shows the change in membrane potential of S. aureus ATCC 13565 after treatments with RRPCE. The fluorescence intensities of S. aureus treated with RRPCE at 1 MIC and 2 MIC were significantly higher than that of control group (p < 0.05), indicating that RRPCE treatment could cause the depolarization of S. aureus cell membrane. Membrane potential depolarization is associated with a decrease in the negative charge in cell, and has also been found in S. aureus cells exposed to OOPE, CA, and ATCE (Gong et al., 2021; Guo et al., 2020a; Guo et al., 2020b).
In contrast, the hyperpolarization of membrane potential results from a change in intracellular pH and potassium efflux, and occurs in S. aureus strains after treatments with 2R, 3R-dihydromyricetin, EUMFE, and chlorogenic acid (Li et al., 2014; Wu et al., 2017; Xing et al., 2022a).
Increased cell membrane permeability
As a DNA staining reagent, PI can penetrate the membrane of damaged cells and then show red fluorescence, and therefore, PI staining can be used to evaluate the degree of cell membrane damage (Yang et al., 2020). As shown in Figure 3, S. aureus ATCC 13565 cells treated with RRPCE at 1 MIC and 2 MIC showed more red fluorescence than untreated cells, indicating that after treatments with RRPCE, the cell membrane integrity of tested bacteria was destroyed. Similar results were found in S. aureus cells treated with EUMFE and limonene (Han et al., 2021; Xing et al., 2022a). Furthermore, the amounts of nucleic acid and protein flowing out of S. aureus ATCC 13565 were measured, as shown in Figure 4.

Fluorescence microscope images of Staphylococcus aureus ATCC 13565.
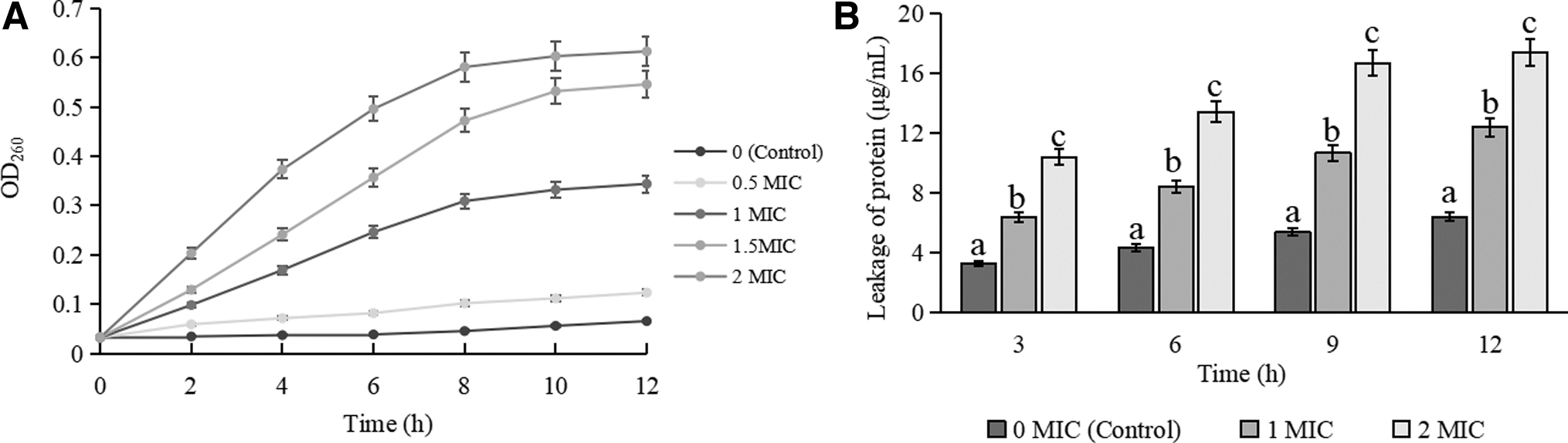
Figure 4A shows that nucleic acid leakage increased significantly with the increase of RRPCE concentration (p < 0.05). Figure 4B shows that the amount of protein efflux from S. aureus treated with RRPCE was also significantly increased compared with untreated cells (p < 0.05). These results suggested that the cell membrane permeability of tested cells increased after RRPCE treatments, which is consistent with the results of Chang et al. (2021) and Xing et al. (2022b), who found that the nucleic acids and proteins were severely leaked from C. sakazakii and S. Typhimurium exposed to CBCE. The above contents indicate that RRPCE, similar to other natural extracts, can destroy the cell membrane integrity of tested cells, and then improve the permeability of cell membrane, eventually leading to the outflow of a large number of nucleic acids and proteins from cell.
Changes in cell morphology
Figure 5 displays the SEM and TEM images of S. aureus ATCC 13565 cells after treatment with RRPCE at 0 MIC, 1 MIC, and 2 MIC for 4 h, respectively. SEM images indicated that compared with untreated cells (Fig. 5A), the surface of cells treated with 1 MIC of RRPCE became rough, and there appeared bubbles (Fig. 5B), while the surface of cells treated with 2 MIC of RRPCE showed cracks and fragmentation (Fig. 5C). TEM images showed that compared with the control cells (Fig. 5D), the cell fluid of S. aureus treated with RRPCE leaked, and the amount of leakage increased with the increase of RRPCE concentration. Moreover, when the action concentration of RRPCE was 2 MIC, the tested cells showed obvious deformation (Fig. 5F).
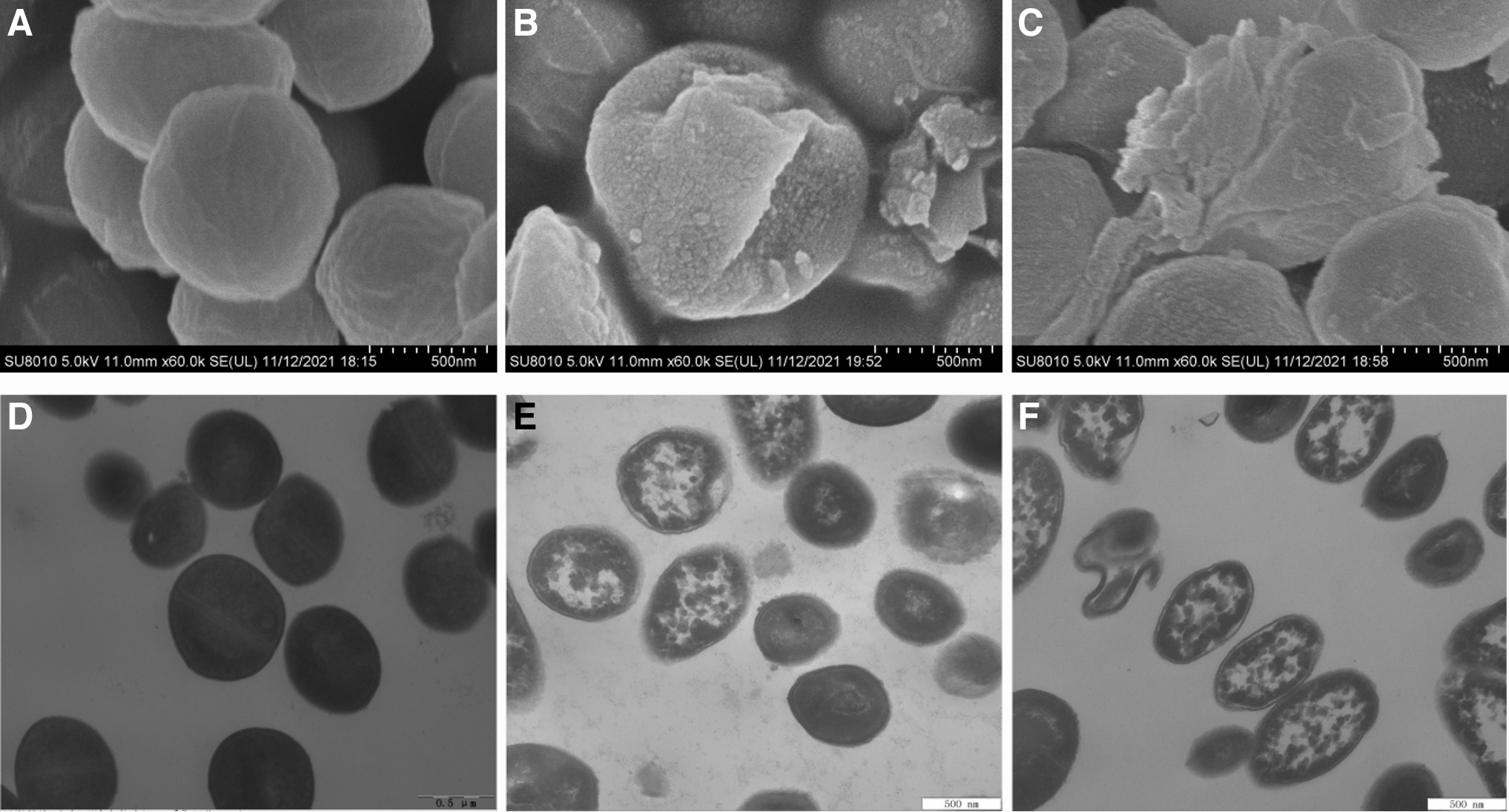
Effect of RRPCE on cell morphology of Staphylococcus aureus ATCC 13565. SEM images:
A similar phenomenon occurred in S. aureus exposed to OOPE, CA, ATCE, and EUMFE, suggesting that the inhibitory pattern of these natural extracts was irreversible bactericidal rather than bacteriostatic effects on S. aureus. Previous studies have found that these irreversible damages are mainly because natural products changed the structure of cell walls and membranes, improved membrane permeability, and caused cell fluid leakage (Fei et al., 2018; Kang et al., 2022; Zhang et al., 2016). In contrast, the surface of C. sakazakii treated with blueberry juice and blueberry proanthocyanidins showed stomata and vesicles, but no cell fluid leakage was observed, leading to continued growth of the treated cells in medium (Joshi et al., 2014).
Antibacterial effect of RRPCE on S. aureus during storage of cooked beef
As shown in Figure 6A, S. aureus ATCC 13565 was significantly inhibited on RRPCE-treated cooked beef compared with the untreated sample during the 9-d storage period (p < 0.05). Especially when the concentration of RRPCE was 2 MIC (3 mg/mL), on the sixth day of storage, the S. aureus viable count of treated sample decreased by ∼2.5 log CFU/mL compared with that of control group. In the previous studies, on the sixth day of storage, Gong et al. (2021) found that CA treatment with 2 MIC (10 mg/mL) reduced 1 log CFU/mL of S. aureus on cooked beef; Xing et al. (2022a) detected a reduction of ∼1.5 log CFU/mL of S. aureus on cooked beef treated by 2 MIC (80 mg/mL) of EUMFE. These results further demonstrated the superiority of RRPCE treatment in inhibiting S. aureus during cooked beef storage.

Change in pH
Table 2 shows the effect of RRPCE on the pH of cooked beef samples during storage. The results showed that during the entire storage period from day 0 to 9, the pH of RRPCE-treated cooked beef was significantly lower than that of untreated sample (p < 0.05), and decreased with the increase of treatment concentration, which was consistent with the previous studies of Gong et al. (2021) and Xing et al. (2022a). This should be due to the fact that RRPCE contains a large amount of organic acids, and the pH of meat products will be significantly reduced after soaking in RRPCE solution.
Effect of Rosa roxburghii Tratt Pomace Crude Extract on the pH of Cooked Beef During Storage at 4°C for Days
Values were expressed as mean ± SD of triplicate experiments. When p < 0.05, the difference between the groups was significant: different lowercase letters in the same row indicate significant differences between groups; different uppercase letters in the same column indicate significant differences between groups.
Some studies have reported that low pH is one of the important factors that natural extract can exert an antibacterial activity (Fei et al., 2020; Guo et al., 2020b; Ozaki et al., 2020), and Li et al. (2016) found that the antibacterial activity of the natural extract added with organic acids was stronger than that without organic acids. In addition, the pH of samples in each group did not change significantly over time (p > 0.05), which is because meat in the heating process can accumulate a large number of soluble substances with pH buffer function, so that cooked meat products in a longer period of time maintain a relatively stable pH (Lawrie, 2006).
Effect of RRPCE on color of cooked beef
As shown in Table 3, compared with the control samples, the L* and b* values of cooked beef treated with RRPCE were significantly decreased (p < 0.05), while the a* value was significantly increased (p < 0.05). During the storage period, myoglobin in cooked beef is easily oxidized to ferric myoglobin, resulting in a decrease in a* value and an increase in b* value of cooked beef (Verma et al., 2019). However, the addition of RRPCE effectively preserved the original color of cooked beef, probably through its antioxidant activity and similar color to cooked beef. Similarly, Xing et al. (2022b) applied CBCE to the preservation of cooked chicken, and believed that if the color of natural preservative was close to the product color, it would be more conducive to maintain the original color of meat product and be accepted by consumers.
Effect of Rosa roxburghii Tratt Pomace Crude Extract on the Color of Cooked Beef During Storage at 4°C for Days
Values were expressed as mean ± SD of triplicate experiments. Different lowercase letters in the same row indicate significant differences between groups (p < 0.05); different uppercase letters in the same column indicate significant differences between groups (p < 0.05).
ΔE* , total color difference; a* , redness; b* , yellowness; L *, lightness.
In addition, a* and ΔE* values of cooked beef were significantly increased, while L* and b* values were significantly decreased with the increase of RRPCE concentration (p < 0.05). This result is similar to the studies of Gong et al. (2021) and Xing et al. (2022a) who used EUMFE and CA in the preservation of cooked beef.
Effect of RRPCE on TVB-N of cooked beef during storage
TVB-N refers to alkaline nitrogen-containing substances such as ammonia and amines produced by the decomposition of protein in animal food under the action of microorganisms and endogenous enzymes (Bekhit et al., 2021). A higher TVB-N value indicates more spoilage bacteria in meat products, and so it is commonly used to assess the freshness of meat and meat products (Li et al., 2017). Figure 6B shows that the TVB-N values of cooked beef in the RRPCE treatment group were always significantly lower than that in control group during storage period (p < 0.05), which suggests that RRPCE effectively inhibits the growth of spoilage microorganisms, including S. aureus.
On the sixth day of storage, the TVB-N value (about 13.8 mg/100 g) of cooked beef treated with 40 mg/mL of EUMFE was higher than that (8.95 mg/100 g) of cooked beef treated with 1.5 mg/mL of RRPCE (Xing et al., 2022a), which may be due to the stronger antibacterial activity of RRPCE. In addition, e-polylysine, tea polyphenols, antibacterial protein from Rahnella aquatilis L103, and Maillard reaction products of shrimp shell chitosan have been demonstrated to significantly reduce the TVB-N in food, and can be used in the preservation of beef or beef products (Li et al., 2021; Tao et al., 2021; Zhang et al., 2022).
Conclusion
As a crude extract, RRPCE showed excellent antibacterial effect against S. aureus. The decrease of intracellular ATP level, depolarization of cell membrane, destruction of cell membrane integrity, leakage of nucleic acid and protein, and impaired cell morphology led to the death of RRPCE-treated S. aureus. Meanwhile, RRPCE could effectively inhibit the growth of S. aureus on cooked beef during storage, maintain the color of cooked beef, and reduce the production of TVB-N, which indicated that RRPCE had great potential as a natural preservative.
Footnotes
Acknowledgment
The authors acknowledge the contribution provided by the Northeast Agricultural University for the experimental equipment, including inverted fluorescence microscope, SEM, and TEM.
Authors' Contributions
X.C.: Conceptualization, methodology, formal analysis, and writing—original draft. X.Z.: Methodology, formal analysis, and supervision. C.F.: Methodology and formal analysis. Q.S.: Methodology and formal analysis. Z.T.: Formal analysis and supervision. D.H.: Methodology and formal analysis. Y.L.: Formal analysis and validation. F.M.: Data curation and software. Y.C.: Supervision and funding acquisition. X.W.: Conceptualization, methodology, formal analysis, and supervision. P.F.: Conceptualization, methodology, supervision, funding acquisition, formal analysis, and writing—original draft. All the authors have read and agreed to the published version of the article.
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This research was supported by the Natural Science Foundation of Guizhou Province (Qiankehe foundation-ZK [2022] General 008), the Guiyang Science and Technology Achievements Transformation Cultivation Program (Zhuke Contract [2022]5-13), and the Natural Science Foundation of Henan Province (212300410137).
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.