
Abstract
Rhizopus nigricans is a widespread phytopathogen in fruits and vegetables that can cause considerable economic effects and resource waste. Flavonoids from Sedum aizoon L. (FSAL) have specific antifungal activities. This study selected FSAL as an antifungal to prolong the preservation of fruits and vegetables. The results showed that the mycelial morphology and ultrastructure were damaged by the FSAL treatment (1.0 minimum inhibitory concentration), led to the increase of reactive oxygen species and malondialdehyde, and affected the activity of key enzymes in the glycolytic pathway, such as lactic dehydrogenase, pyruvate kinase, and hexokinase of R. nigricans. Key genes in glycolysis were upregulated or downregulated. In addition, in the treatment and control groups, 221 differentially expressed genes were found, including 89 that were upregulated and 32 that were downregulated, according to the transcriptome results. The differential genes were mainly enriched in glycolysis, pyruvate metabolism, and citrate cycle pathways. The results revealed some insights into the antifungal mechanism of FSAL against R. nigricans and offered a theoretical foundation for its advancement as a novel plant-derived antifungal agent.
Introduction
R
Flavonoids, the most abundant polyphenols, are widely found in the roots, stems, leaves, flowers, and seeds of vegetables (Alsharairi, 2021). Flavonoids have a broad spectrum of activities, of which the antifungal properties are mostly investigated (Vica et al., 2022).
Meanwhile, previous studies found that flavonoids from Sedum aizoon L. (FSAL) with remarkable antifungal activity against Botrytis cinerea, which may cause leakage of macromolecules such as nucleic acids, led to accumulation of malondialdehyde (MDA) and relative oxygen species, and disrupted the ultrastructure of B. cinerea (Wang et al., 2022). RNA sequence analysis was used to investigate the underlying antifungal mechanism of FSAL on Penicillium italicum, and the results showed that FSAL inhibited genes related to the integral components of membrane, oxidation–reduction process, mitochondrion, ribosome, and amino acid metabolism (Luo et al., 2020). The antifungal mechanism of FSAL against R. nigricans has not been documented, though.
We examined the effect of FSAL on R. nigricans cell membranes and revealed the effect of FSAL on the R. nigricans glycolytic pathway by transcriptome. We used scanning electron microscopy (SEM) and transmission electron microscopy (TEM) observation to observe the effect of FSAL on the mycelial morphology and ultrastructure of R. nigricans, and the intracellular content reactive oxygen species (ROS) were detected in intracellular concentrations using the fluorescent probe 2′,7′-dihydrodichlorofluorescein diacetate (DCFH-DA). The activities of key enzymes in the glycolytic pathway were also measured to reveal the effect of FSAL on key enzymes of R. nigricans. The effect of FSAL on the metabolic pathway of R. nigricans was also explored by transcriptome.
Materials and Methods
Materials and chemicals
Peixian County provided the harvest for the Sedum aizoon L. (Jiangsu, China). The Hangzhou Microbial Reagent Co., Ltd. provided the potato dextrose agar (PDA) medium and the potato glucose broth (PDB) medium (Hangzhou, China). The company Shuoguang Electronics Technology Co., Ltd. provided purified water (Shanghai, China). All other reagents were of analytical grade.
Preparation of flavonoid extracts
Preparation of FSAL was according to a revised method of Xu et al. (2018), roasting the raw material in an oven at 60°C until dry, grinding it into a powder, and passing it through a sieve (60 mesh). Raw material was mixed with ethanol (70%) in a ratio of 1:10, homogenized by ultrasonication, and warmed in a 60°C bath for 2 h. Then followed by filtering and concentration using a vacuum rotary evaporator (RE-52, Yarong, Shanghai, China). Impurities were removed with anhydrous ethanol, purified with AB-8 resin, adsorbed with 30% ethanol, dissolved with 70% ethanol, and then again concentrated in a rotating vacuum evaporator. In this experiment, the content of flavonoid powder was 88% after purification by AB-8 macroporous resin. The concentrates were lyophilized and kept at −80°C.
Fungal strain and culture conditions
R. nigricans (CICC, 41346) was obtained from the China Industrial Microbial Strain Conservation and Management Center, and the strains were inoculated into PDA plate medium for activation under aseptic conditions before the experiment, and then incubated at 25 ± 1°C for 7 d, followed by secondary activation under the same conditions and set aside. The plates were washed with sterile water to scrape off the spores and to remove the hyphae; the spores were collected and filtered through four layers of sterile gauze, adjusting the spore count to 1 × 106 spores/mL with a hemocytometer (Shanghai Yongchuan Co., Ltd., Shanghai, China).
Detection of minimum inhibitory concentration
Minimum inhibitory concentration (MIC) was determined according to the method put forward by Low et al. (2011) with some modifications. FSAL with PDA medium to prepare a mixture at different concentrations (0.00, 0.23, 0.45, 0.90, 1.80, 3.60, 7.20, and 14.40 mg/mL) and poured into plates for cooling, and control plates were prepared without FSAL. Twenty microliters of spore suspension at a concentration of about 106 spores/mL was inoculated and coated well. The plates were incubated at 25 ± 1°C for 2 d, and the MIC, which was determined using five plates per group and three replications, was taken as the lowest FSAL concentration without mycelium growth.
Gene expression analysis by real-time quantitative polymerase chain reaction
R. nigricans spores were incubated in PDB medium for 48 h and mycelium was collected. The mycelium was subjected to FSAL (1 MIC) except the control samples, and treatment for 4 h with simultaneous shaking. Subsequently, the fungal cells were quenched with liquid nitrogen and stored at −80°C before analysis.
Total RNA was isolated from each of the aforementioned samples using the HiPure fungal RNA extraction kit (Magen Biotechnology Co., Ltd, Guangzhou, China) in accordance with the kit's instructions. RNA concentration was calculated using NanoDrop 2000 (OD260/280 values in the range of 1.8–2.2), and RNA integrity was evaluated using agarose gel electrophoresis. Using the Novozymes HiScript® II Q RT SuperMix for quantitative polymerase chain reaction (qPCR) (+gDNA wiper) kit, RNA of acceptable quality was reverse transcribed to create the complementary DNA (cDNA). The PCR was then performed using the ABI Prism 7500 rapid qPCR kit (Roche Switzerland Ltd., Basel, Switzerland) under the following conditions: initial denaturation for 10 s at 95°C, 95°C for 10 s, 60°C for 30 s for 40 cycles, and finally 95°C for 10 s. After qPCR was finished, the RNAs were simultaneously examined.
Total RNA was extracted from each of the aforementioned samples using the HiPure method. After the qPCR was finished, the relative expression levels of target genes were calculated using the 2−ΔΔCT method (Zhao et al., 2022) and 18S rRNA (Zhang et al., 2017) as an internal control gene. Primers were created using the Applied Biosystems Primer Express 5.0 (Primer Company, Canada). The genes of glycolysis (HK, PFK) and the sequence of 18S are shown in Table 1. Among them, HK and PFK are keys and irreversible enzymes in the glycolytic pathway and an important link in energy production. The glycolysis Kyoto Encyclopedia of Genes and Genomes (KEGG) PATHWAY and National Center for Biotechnology Information (NCBI) were used to gather the data mentioned above.
Primer Sequences of Genes Associated with Glycolysis of Rhizopus nigricans
cDNA library construction and sequencing
Shanghai Meiji Biomedical Technology Co. prepared the RNA sequencing and the cDNA library. Briefly, using magnetic beads and Oligo (dT), total RNA was used to extract the messenger RNA (mRNA), which was then cut into minuscule fragments at random. Six-base random hexamers were added to invert the mRNA template and create a cDNA chain through the activity of reverse transcriptase, which is then followed by two-stranded synthesis to create a stable double-stranded structure. AMPure XP beads were used for final fragment selection and PCR was performed to obtain the final sequencing library, which after clearing quality control is then sequenced on the Illumina platform.
Data analysis, differentially expressed gene screening, and bioinformatic analysis
Using the Illumina NovaSeq 6000 (Illumina, San Diego, CA, USA), the generated libraries were sequenced. Sequences with linkers, unidentified sequences, and low-quality sequences were all disqualified from data inclusion. Genome mapping analysis of clean reads was performed using Bowtie 2 software (Version 2.4.4; Langmead B, Washington, DC, USA). Each sample's gene expression was measured using Subread software. The normalized value of gene expression in all samples was determined using fragment number per kilobase million. On the basis of the absolute value of log 2 (fold change) >0, differentially expressed genes (DEGs) p < 0.05 were then tested for treatment versus control groups. The DEGs were subjected to Gene Ontology (GO) functional enrichment analysis and KEGG pathway enrichment analysis.
Validation of real-time quantitative polymerase chain reaction
Similar to the RNA-Seq analysis above, real-time quantitative polymerase chain reaction (RT-qPCR) analysis of R. nigricans was carried out. The relevant primers are presented in Table 2 for the following six DEGs: TRINITY_DN12636_c0_g, TRINITY_DN3792_c0_g1, TRINITY_DN8079_c0_g1, TRINITY_DN1650_c1_g1, TRINITY_ DN545_c0_g1, and TRINITY_DN12244_c0_g1 were chosen at random from the library for various metabolic pathways. TRINITY_DN12636_c0_g encodes phosphoglycerate kinase activity, TRINITY_DN3792_c0_g1 encodes cellular aldehyde metabolic process, TRINITY_DN8079_c0_g1 is involved in oxidoreductase activity, TRINITY_DN1650_c1_g1 encodes mitochondrial matrix, TRINITY_ DN545_c0_g1 is an important component of the tricarboxylic acid cycle and is involved in mitochondrial energy metabolism, function, and respiratory chain transfer, and TRINITY_DN12244_c0_g1 encodes transferase activity. Data were processed uniformly for each sample with three biological replicates and standardized by 18S rRNA expression. A t-test was used for statistical analysis.
Primers Used in This Study
Electron Microscope Observation of Fungal Morphological Changes
Scanning electron microscopy
A modified protocol based on a method described elsewhere (Wallace et al., 2017) was used to observe the morphological change of R. nigricans treated with FSAL. Briefly, mycelium was collected and FSAL at a concentration of 1 MIC was added, and the mycelium with the same volume of autoclaved phosphate buffered saline (PBS) was used as control. Samples were collected after 0, 4, and 8 h of treatment, respectively, followed by three PBS (pH 7.4) buffer washings. The samples were dehydrated with ethanol (30%, 50%, 70%, and 90%), replaced with tert-butanol, freeze-dried, glued, sprayed with gold, and observed and photographed in SEM (S-3400N; Hitachi, Japan) after being fixed in 3.0% glutaraldehyde (0.1 M PBS adjusted to pH 7.2) for 12 h at 4°C.
Transmission electron microscopy
The pretreatment for TEM and SEM was the same. With a few changes, the previously published approach was mentioned in TEM (Wang et al., 2022). Mycelium that had been treated was first fixed for an hour in 1% osmium acid (0.2 M PBS adjusted to pH 7.2) and then fixed for an additional hour in 2.5% glutaraldehyde at 4°C. The samples were rinsed three times for 15 min with 0.1 M PBS before being dehydrated using ethanol (30%, 50%, 70%, and 90%) for 15 min. Using a diamond knife, the samples were then cut into ultrathin slices of around 70 nm, put in epoxy medium, and dyed with uranyl acetate and lead citrate, and then observed with TEM (H-7650; Hitachi).
Detection of ROS
The intracellular content of ROS was detected in intracellular concentrations using the fluorescent probe DCFH-DA (Nanjing Jiancheng Bioengineering Institute, Nanjing, China), with slight modifications (Ren et al., 2019).
Determination of MDA content
Mycelium was collected, and then, the MDA content was measured using the kit (Nanjing Jiancheng Technology Co., Ltd., Nanjing, China).
Determination of enzyme activity related to glycolysis
Utilizing commercial kits (Nanjing Jiancheng Technology Co., Ltd.) measured the activities of lactic dehydrogenase (LDH), pyruvate kinase (PK), and hexokinase (HK) in accordance with the manufacturer's recommendations.
The content of pyruvic acid of R. nigricans
The kit (Nanjing Jiancheng Technology Co., Ltd.) was used to determine the content of pyruvic acid (PA) in R. nigricans. The amount of PA produced by R. nigricans was determined after the absorbance at 520 nm was measured and the supernatant was collected.
Statistical analysis
Utilizing version 9.4 (SAS Institute, Cary, NC, USA) of the SAS software package program, statistical analysis was carried out. The mean and standard error are used to express all results. The one-way analysis of variance and Duncan's multiple range test were used to assess the data's significance (p < 0.05). The experimental data were processed using Origin 9.0 for analysis and plotting. To ensure the credibility of the data, all experiments were done in triplicate and repeated at least twice.
Results
Detection of MIC
Table 3 displays the findings of the MIC analysis of FSAL against R. nigricans. After 2 d of incubation, when the concentration of FSAL was more than or equal 1.80 mg/mL, no significant colony development was seen, showing that the MIC of FSAL for R. nigricans was 1.80 mg/mL.
The Minimum Inhibitory Concentration of Flavonoids from Sedum aizoon L. on Rhizopus nigricans
Note: + indicated significant colony growth and − indicated no obvious colony growth.
FSAL, flavonoids from Sedum aizoon L.
Illumina HiSeq sequencing defining DEGs
Transcriptome data have been uploaded to NCBI (
The results are displayed in Figure 1, the transcriptome sequencing data of the treated and control samples were normalized, and then scatter plots were plotted for the expression of all genes. The red and green portions depict the DEGs' up- and downregulated genes, respectively, while the gray part shows the DEGs with insignificant differences. From Figure 1C, 221 distinct genes were found to be differentially expressed in the FSAL (1 MIC)-treated group compared with the control group, 89 of these genes showed downregulated expression, accounting for 40.3% of the total differentially expressed genes, and 132 differential genes were expressed as downregulated, accounting for 59.7%. The results demonstrated either upregulation or downregulation of gene expression in FSAL-treated R. nigricans, indicating that it had a repressive effect on R. nigricans.
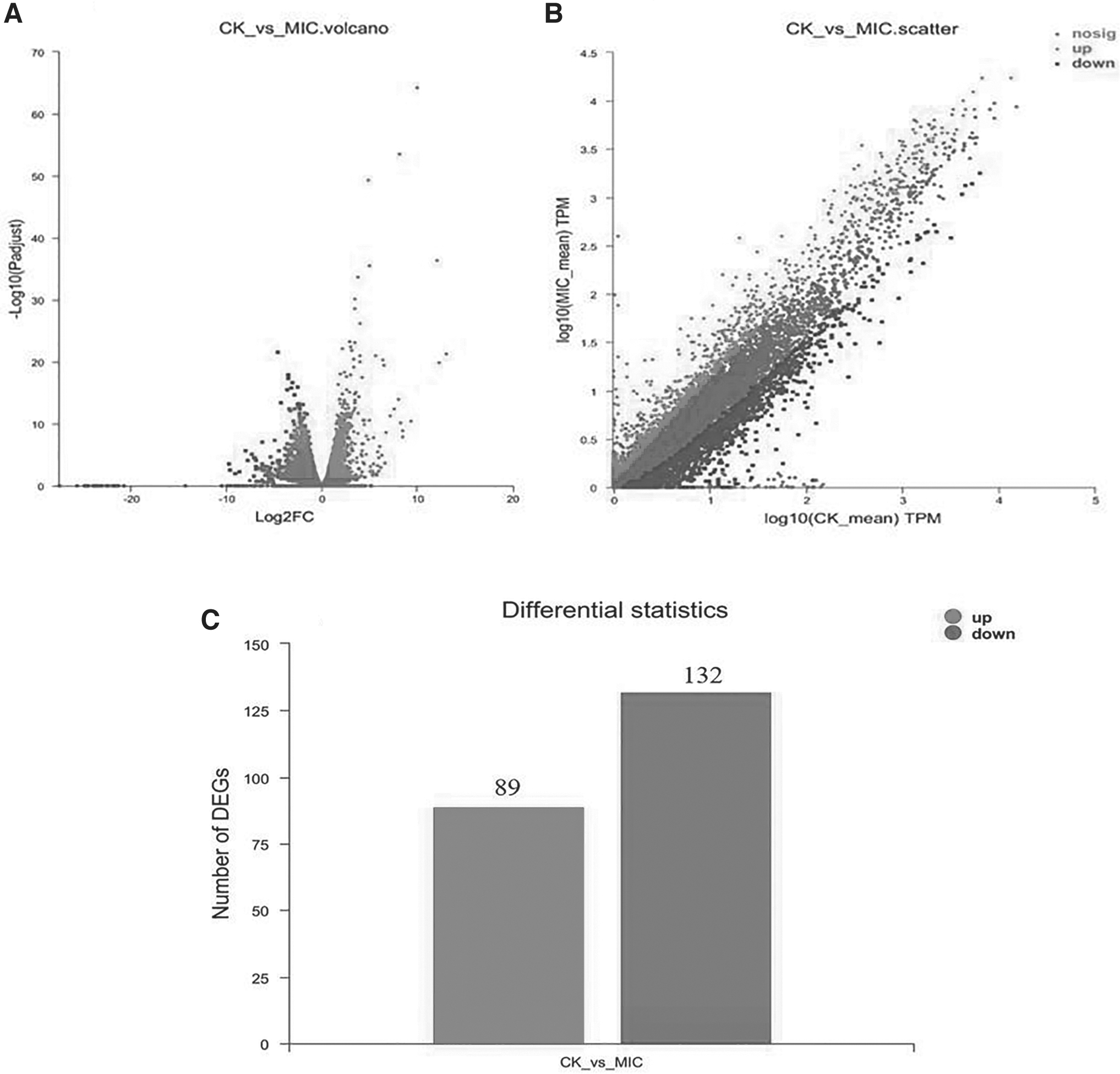
Specifics of DEGs of Rhizopus nigricans in the FSAL-treated and control groups.
Gene function classification using GO enrichment and KEGG pathway analysis
GO is a comprehensive database created by the GO Consortium to categorize and summarize all gene-related research results worldwide. The findings of the GO functional enrichment analysis are shown in Figure 2: molecular function (MF), biological process (BP) and cellular component (CC). This method enables a detailed comparison of DEG function between the FSAL treatment group and the control group. For the MF aspect of the GO functional classification, a total of 8 components were involved, and the FSAL treatment mainly affected the catalytic activity, binding, and other functions. Cellular processes, metabolic processes, and bioregulation were the key areas of emphasis for the BP category. In the functional division of GO, the CC category, a total of 6 contents were involved, and the most important ones were functions such as membrane fraction and organelle fraction. The results showed that FSAL treatment has changed the cell membrane of R. nigricans.
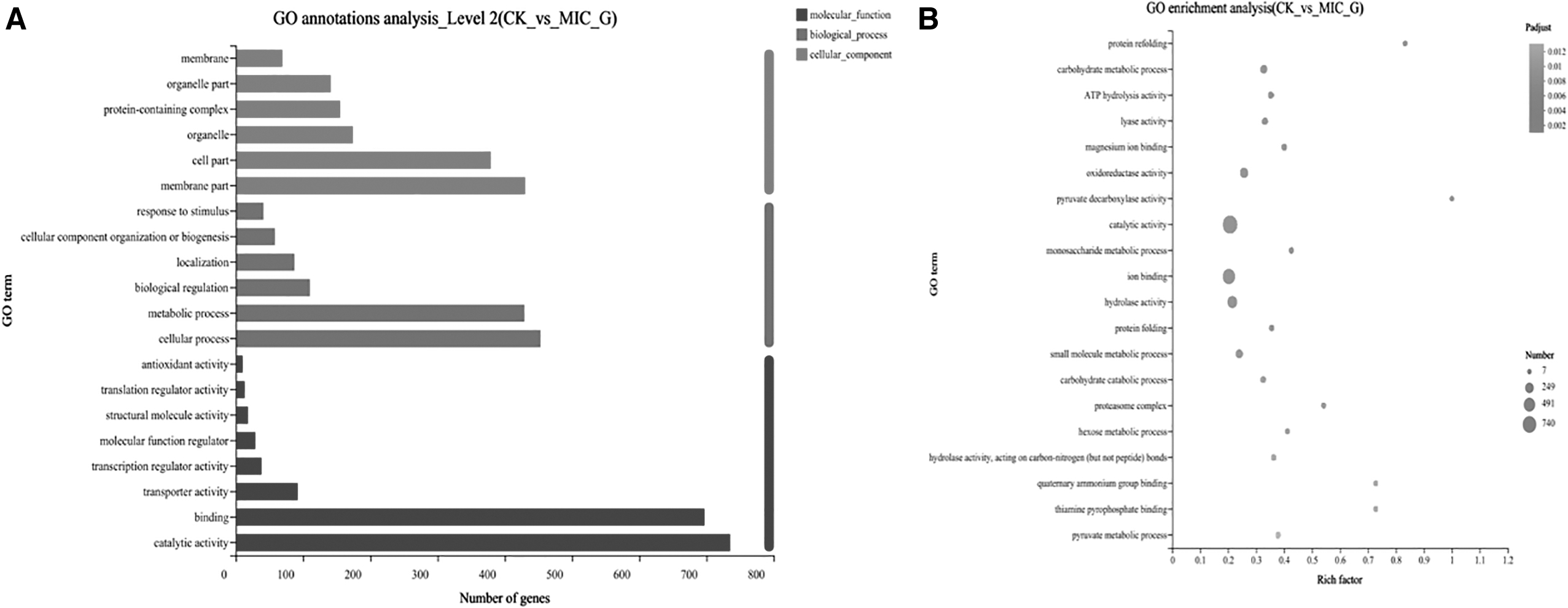
DEGs between the two sample groups were analyzed by GO.
To determine which metabolic pathways the genes in this gene set are primarily involved in, the KEGG PATHWAY enrichment analysis was carried out. When the corrected p value (false discovery rate) was <0.05, this KEGG pathway was found to be significantly enriched in the gene set using Fisher's exact test. As shown in Figure 3, DEG is more involved in glycolysis, pyruvate metabolism, longevity regulation pathway-polysaccharide, ascorbate, and aldehyde metabolism, glycerol ester metabolism, interconversion of pentose and glucuronide, methane metabolism, tricarboxylic acid cycle, and ATP-Binding Cassette transporter protein pathways, respectively. Among the top 20 most significantly enriched pathways, 6 were carbohydrate metabolic pathways, and the highest enrichment in significantly enriched pathways was in the glycolysis and pyruvate metabolic pathways, respectively. It indicated that these two pathways played important roles in the inhibition mechanism of FSAL against R. nigricans.

KEGG enrichment analysis of Rhizopus nigricans after FSAL treatment. The horizontal axis indicates the degree of enrichment and the vertical axis indicates the enriched KEGG pathway; the size of the dots indicates the number of differential genes enriched to a certain KEGG pathway; the color of the dots indicates different p-values; the rich factor indicates the number of differential genes belonging to a certain KEGG pathway and the total number of genes belonging to this KEGG pathway. ABC, ATP-Binding Cassette; FDR, false discovery rate; KEGG, Kyoto Encyclopedia of Genes and Genomes; TCA, tricarboxylic acid cycle.
Cell membrane and internal components
The cell membrane in fungi is an important cellular barrier that absorbs nutrition and exchanges material and energy with the surrounding environment (Sant et al., 2016), and once the cell membrane has been damaged, fungal cells become inactive due to a number of dysfunctions (Chueca et al., 2017).
The membrane damage and ultrastructural changes of FSAL on R. nigricans were observed using SEM and TEM. As shown in Figure 4, the SEM observations showed the effect of FSAL on the morphology of R. nigricans mycelium, in the SEM images, the mycelium of the untreated group showed normal and complete tubular structure, and the surface of the mycelium was relatively clear and smooth. While the mycelium morphology changed significantly after FSAL (1 MIC) treatment, the cells of FSAL-treated group showed folds, local depressions due to loss of contents, and a rougher surface (Fig. 4B). A uniform distribution of electron density in the cytoplasm was observed in the TEM images (Fig. 4C). However, treatment of R. nigricans with FSAL resulted in a nonuniform cytoplasm with vacuolation and other phenomena compared with the control (Fig. 4D), indicating that some intracellular material leaked out through cell membrane damage or permeability.
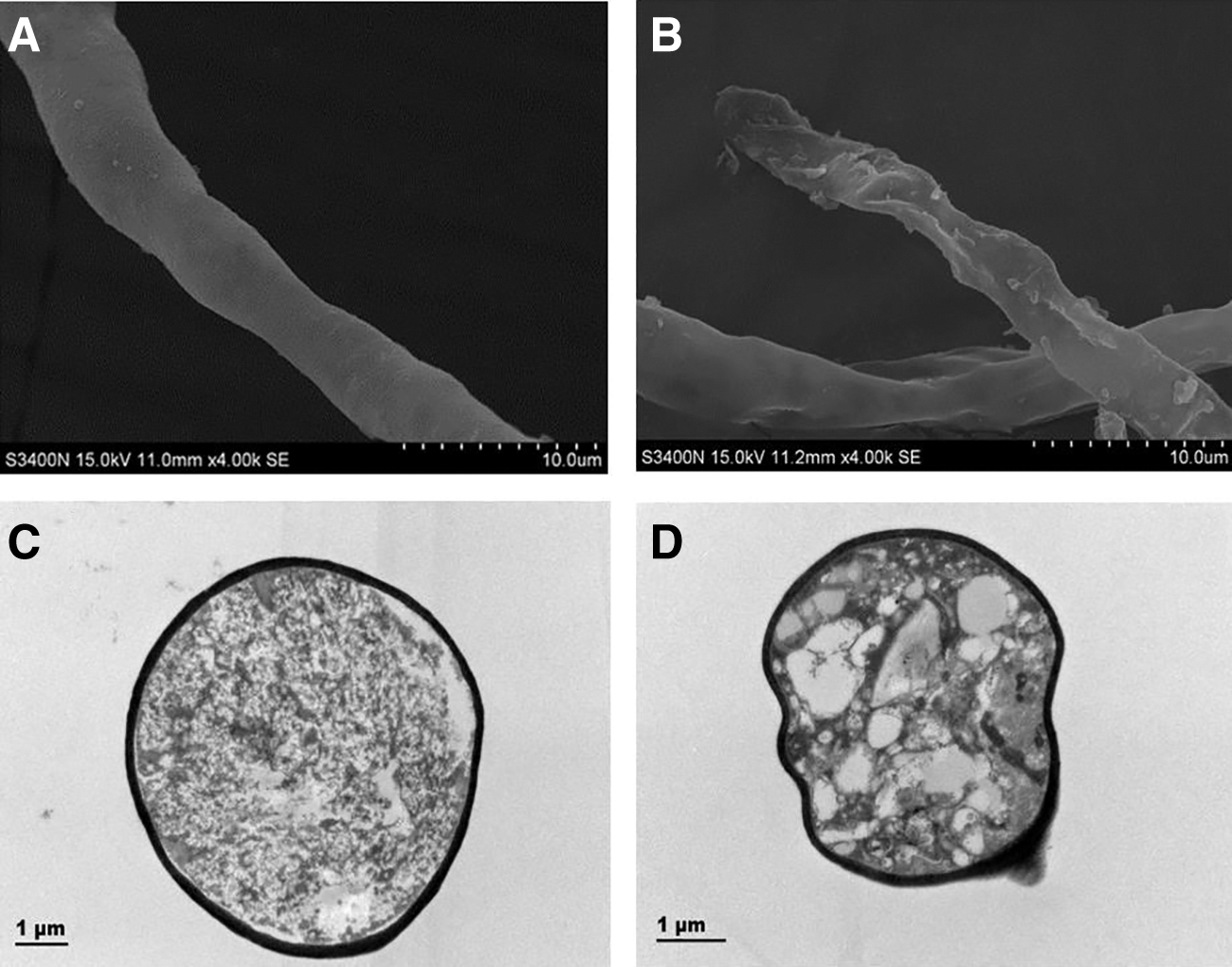
Scanning electron microscopy and transmission electron microscopy images of Rhizopus nigricans.
Meanwhile, the transcriptome results demonstrated that 15 genes associated with membranes appeared significantly increased (such as TRINITY_DN5339_c0_g1 and TRINITY_DN14896_c0_g1) and 13 genes significantly decreased (such as TRINITY_DN14267_c0_g1, TRINITY_DN13721_c0_g1, and TRINITY_DN501_c1_g1) in the expression following FSAL therapy in comparison with the control group. The changes in genes controlling biofilm formation suggest that the integrity of the membrane of R. nigricans was disrupted under the influence of FSAL. One upregulation (TRINITY_DN3838_c0_g1) and three downregulations (TRINITY_DN9445_c1_g3, TRINITY_DN9277_c0_g1) were observed in the genes of the membrane protein complex. The change of biofilm composition may cause the change of membrane fluidity and permeability.
The results showed that FSAL can significantly enhance the cell membrane permeability of R. nigricans, and disrupted the cell membrane structure and certain disruption of ionic homeostasis, thus causing abnormalities in the cell membrane and organelles of R. nigricans, and affecting the normal physiological metabolism of the cells.
ROS and MDA
ROS are aerobic metabolites produced by biological cells during metabolism (Martinez-Cisuelo et al., 2016), and the accumulation of ROS attacked the cell membrane's unsaturated fatty acids, causing lipid peroxidation that eventually ruptured the cell membrane and triggered apoptosis (Hwang and Lim, 2015; Qian et al., 2016). The fluorescence results observed by fluorescence microscopy and the measured fluorescence intensity data are shown in Figure 5A, B. It can be seen that the fluorescence intensity in the control group was lower, which was substantially different (p < 0.05) from the fluorescence intensity after 4 h of FSAL treatment. In addition, the fluorescence intensity was increased and the number of stained R. nigricans spores steadily increased with a dose-dependent impact as the FSAL treatment duration was extended. This result indicated that the treatment with FSAL contributed to the significant accumulation of ROS in R. nigricans.
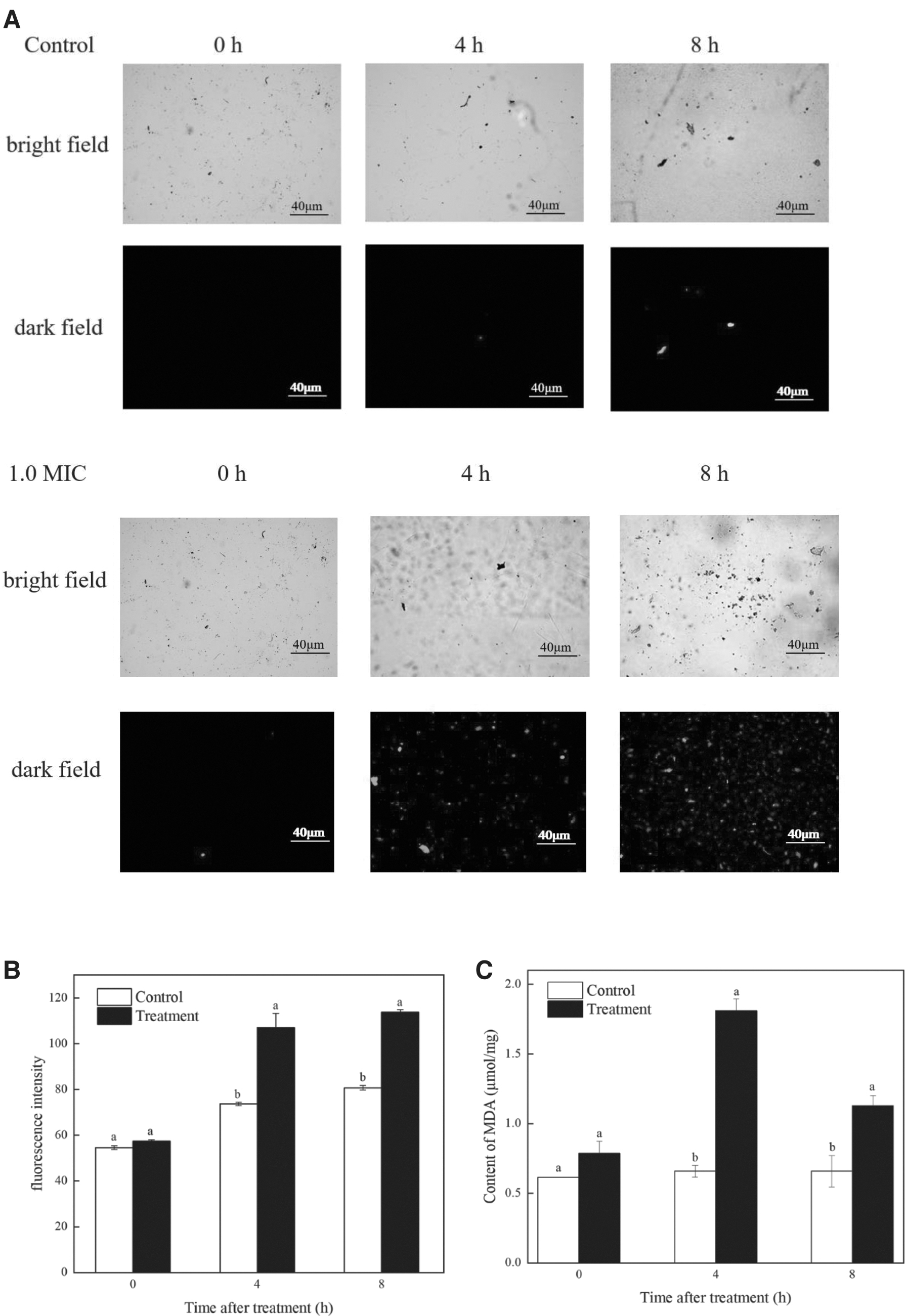
Effect of FSAL on ROS and MDA of Rhizopus nigricans.
MDA is the primary byproduct of cytoplasmic membrane lipid peroxidation damage, accumulation of MDA speeds up oxidative damage, and measurement of MDA levels can be used to assess membrane lipid peroxidation levels (Aguilar Diaz De Leon and Borges, 2020). Figure 5C shows the MDA levels in R. nigricans following FSAL treatment. The overall MDA content in the R. nigricans mycelium after FSAL treatment showed an increasing trend. When the MDA value was 1.81 mol/mg at 4 h (p < 0.05), compared with the control group, the MDA content in the FSAL-treated group was significantly higher. The data suggested that FSAL exacerbated the degree of peroxidative damage and enhanced the pace and severity of lipid peroxidation in R. nigricans.
In addition, the activity of oxidoreductase (TRINITY_DN8079_c0_g1) in R. nigricans was dramatically impacted by FSAL, according to transcriptome sequencing studies, indicating that FSAL may cause oxidative stress in R. nigricans and large amounts of ROS to be produced, causing DNA damage and intracellular lipid peroxidation, all of which result in cell death. The results supported the conclusions described above that FSAL administration can cause ROS and MDA to accumulate, boosting the Ca 2+ content in R. nigricans, creating Ca2+ imbalance, oxidative stress to the fungal, hastening cellular senescence, and even death.
Carbohydrate metabolism
Glucose oxidation is a common energy metabolic pathway in microorganisms and the ultimate metabolic pathway for biomolecules such as lipids and proteins. Inhibition of key enzymes in the glucose oxidation metabolic pathway will affect the energy production and normal growth of the organism (Gonzalez et al., 2017). The production, catabolism, and interconversion of carbohydrates in living things are all governed by numerous biochemical processes known as carbohydrate metabolism, including glycolysis/gluconeogenesis (KEGG: map00010) and pyruvate metabolism (KEGG: map 00620).
The effect of FSAL on the R. nigricans glycolytic pathway was determined by measuring the activity of the key glycolytic enzymes. For HK, the treatment group was substantially different from the control group after 4 h (p < 0.05), and maintained at a low level, as shown in Figure 6A. The HK activity of control was 3.31 and 3.63 times higher than the infected groups after 4 and 8 h, respectively. The results of PK are shown in Figure 6B. FSAL treatment could induce sustained enhancement on the activities of the PK, within the tested 4 and 8 h, and the PK enzyme activity in the FSAL increased 2.40 and 2.54 times compared with the control.
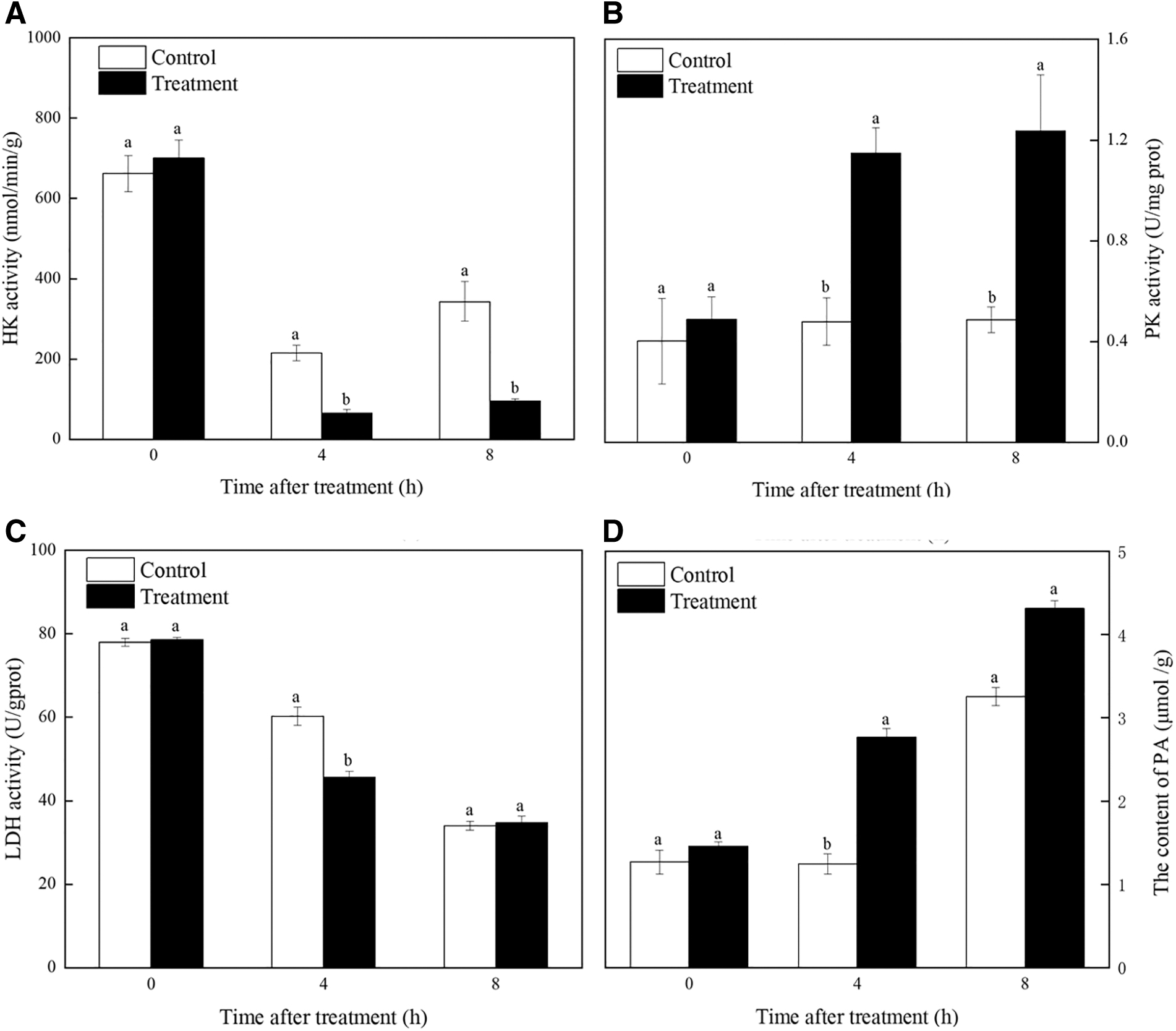
Effect of FSAL on key enzymes in glycolysis and PA content of Rhizopus nigricans.
On the contrary, the activity of the LDH enzyme in the FSAL treatment showed a decreasing trend after 4 h of incubation, and was significantly lower than the control; the LDH enzyme activity in FSAL treatments was 1.32 times lower compared with control after 4 h. However, the significant difference from the control disappeared after 8 h, respectively. As shown in Figure 6D, FSAL treatment significantly accelerated the accumulation of PA in R. nigricans.
We extracted the RNA of R. nigricans under FSAL treatments at 0, 4, and 8 h and detected the expression of HK, PFK, PK, and LDH-encoding genes by RT-qPCR. As shown in Figure 7, RT-qPCR results of HK, PFK, PK, and LDH during glycolysis were consistent with the transcriptome results, HK, LDH were repressed and PFK, PK were increased. In addition, the transcriptome results showed that PFK (TRINITY_DN3960_c0_g1), PK (TRINITY_DN5345_c1_g1), key genes of R. nigricans glycolytic pathway, were upregulated; and HK (TRINITY_DN3_c2_g2), LDH (TRINITY_DN5219_c0_g1) downregulated after FSAL treatment. In summary, FSAL interfered with key enzymes, affected the content of intermediates, and disturbed the key genes in the glycolysis process of R. nigricans.
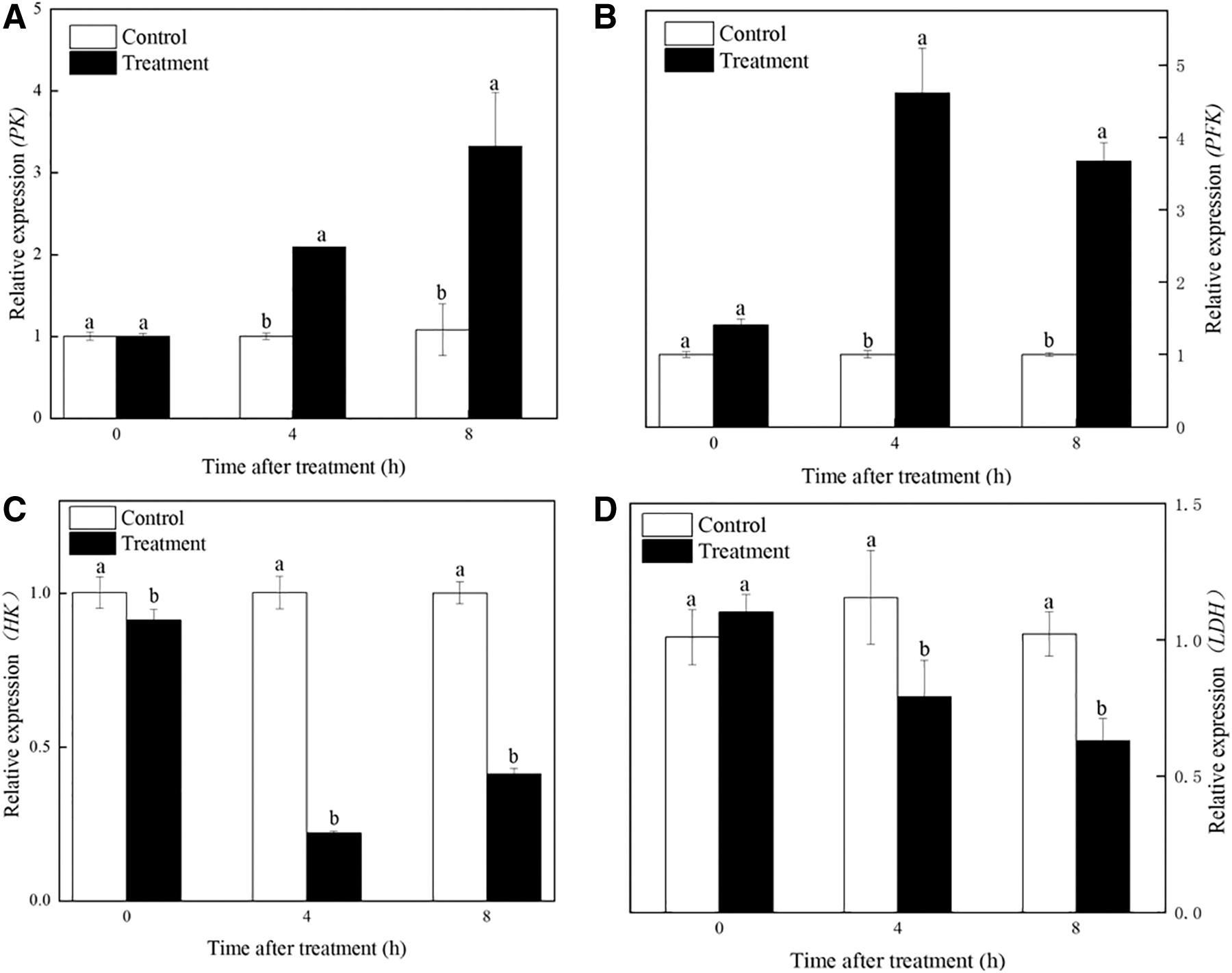
Effect of FSAL on the relative expression levels of genes related to glycolysis of Rhizopus nigricans.
Conclusion
The study was to analyze the mechanism of FSAL inhibition against R. nigricans by combining transcriptomics and bioinformatics. In conclusion, this study confirmed that FSAL had significant antifungal activity against R. nigricans with an MIC of 1.80 mg/mL. Moreover, the antifungal mechanism of FSAL on R. nigricans found that FSAL treatment induced the accumulation of ROS, increased the levels of MDA, and caused intracellular material leakage. Moreover, the activity of crucial genes and enzymes involved in glycolysis was perturbed. The transcriptome analysis further affirmed that the expression of genes involved in glycolysis, pyruvate metabolism, and other pathways was suppressed by FSAL. Therefore, R. nigricans is significantly suppressed by FSAL. The findings mentioned above offer a theoretical foundation for the FSAL inhibition mechanism, which may facilitate the development of antifungal agents with a natural plant origin and provide a basis for fruit preservation applications.
Footnotes
Authors' Contributions
Q.G.: Methodology, investigation, and writing—original draft. K.W.: Investigation and writing—original draft. X.S.: Writing—review and editing. Y.W. and Y.L.: Data curation. X.Z.: Validation and formal analysis. H.W.: Conceptualization, supervision, and project administration. F.X.: Resources and investigation.
Acknowledgments
We thank Key Laboratory of Animal Protein Food Processing Technology of Zhejiang Province and Zhejiang-Malaysia joint research laboratory for agricultural product processing and nutrition.
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This study was supported by the Natural Science Foundation of Zhejiang Province (LY16C200003) and the SRIP project of Ningbo University (2022SRIP3608).