
Abstract
The aim of this study was to assess the antimicrobial activity of oregano essential oil (OEO) against Shigella flexneri and eradication efficacy of OEO on biofilm. The results showed that the minimum inhibitory concentration (MIC) and the minimum bactericidal concentration (MBC) of OEO against S. flexneri were 0.02% (v/v) and 0.04% (v/v), respectively. OEO effectively killed S. flexneri in Luria–Bertani (LB) broth and contaminated minced pork (the initial population of S. flexneri was about 7.0 log CFU/mL or 7.2 log CFU/g), and after treatment with OEO at 2 MIC in LB broth or at 15 MIC in minced pork, the population of S. flexneri decreased to an undetectable level after 2 or 9 h, respectively. OEO increased intracellular reactive oxygen species concentration, destroyed cell membrane, changed cell morphology, decreased intracellular ATP concentration, caused cell membrane depolarization, and destroyed proteins or inhibited proteins synthesis of S. flexneri. In addition, OEO effectively eradicated the biofilm of S. flexneri by effectively inactivating S. flexneri in mature biofilm, destroying the three-dimensional structure, and reducing exopolysaccharide biomass of S. flexneri. In conclusion, OEO exerts its antimicrobial action effectively and also has a valid scavenging effect on the biofilm of S. flexneri. These findings suggest that OEO has the potential to be used as a natural antibacterial and antibiofilm material in the control of S. flexneri in meat product supply chain, thereby preventing meat-associated infections.
Introduction
S
S. flexneri is widely found in food and food processing environments, such as fresh fruit and vegetables, raw oysters, meat, unpasteurized milk, and factory processing equipment surface (Garedew et al., 2016). Planktonic S. flexneri cells can easily adhere to the surface of food materials, food processing equipment, and the inner wall of pipelines, and eventually form bacterial biofilms (Chanin et al., 2019; Kang et al., 2018b; Nickerson et al., 2017). Biofilms are defined as communities of microorganisms that attach to surfaces and are a prevalent mode of growth for microorganisms in nature (Shi and Zhu, 2009). Previous studies have found that 65% of bacterial infections are involved in biofilms (Ramage et al., 2012; Srey et al., 2013), which form a barrier that can prevent or lessen bacterial contact with antimicrobial agents, so they can render their inhabitants more resistant to disinfectant, and also protect viable cells against various environmental stresses such as acid and heat (Crabbe et al., 2019).
Existing control measures used to reduce the risk of Shigella spp. include ionizing radiation (Mahmoud, 2009), ultraviolet radiation (Rajkowski, 2010), and ozone binding with organic acids (Singla et al., 2011). However, these methods are constrained by factors that affect the sensory properties and nutritional content of food, high equipment costs, and low consumer acceptance. Detergents and biocides are common practices used to control biofilms formed by S. flexneri (Yu et al., 2020). Due to the characteristics of the biofilm, which leads to the development of tolerance to detergents and biocides, S. flexneri present inside the biofilm is recalcitrant to the current treatment strategies, and hence, it causes barrier in the food processing industries (Luciano et al., 2016). Therefore, it is crucial to find a safe, effective, and green method to inhibit S. flexneri and its mature biofilm in food and food processing.
Recently, the improvement of living standards has resulted in increasing attention on food processing methods, which has natural, green, and less influence on food sensory (Pandey et al., 2016). Essential oils and other natural bacteriostatic agents and biofilm scavengers have become the focus of research, and studies have shown that some essential oils have strong antibacterial and antibiofilm activities against different microorganisms (Agarwal et al., 2008; Saharkhiz et al., 2012). Oregano essential oil (OEO) is a natural plant essential oil extracted from Origanum vulgare L. (Leyva-López et al., 2017). It is generally recognized as safe (GRAS) by the Food and Drug Administration (FDA) (SP, GRAS-182.10). In addition, OEO has demonstrated anti-inflammatory (Leyva-López et al., 2016), antioxidant (Quiroga et al., 2013), and preventing atherosclerosis (Saljoughian et al., 2017) effects. OEO has antibacterial effect on Staphylococcus aureus, Salmonella enteritidis, and Escherichia coli (Şahin et al., 2004). However, its antimicrobial and antibiofilm activities against S. flexneri have not been studied in any depth.
Based on the factors described above, the antibacterial and antibiofilm ability of OEO against S. flexneri should be more concerned. On the one hand, this study took a brief evaluation about the antimicrobial effect of OEO against S. flexneri and explained the possible mechanism of action from different aspects. On the other hand, to minimize the negative impacts of S. flexneri biofilm in the food processing environment, the efficacy of OEO in inhibiting mature biofilm of S. flexneri was evaluated by measuring the effect of OEO on S. flexneri biofilm on the surface of stainless steel materials commonly used in food processing environment.
Materials and Methods
Reagents
OEO (density 0.939 g/mL at 25°C, CAS 8007-11-2) was acquired from Sigma–Aldrich (Shanghai, China). Luria–Bertani (LB) was purchased from Land Bridge Technology Co. (Beijing, China). All other chemicals and reagents were of analytical grade.
Bacterial strains and growth conditions
S. flexneri ATCC 12022 used in this study was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA); the strain was stored in LB broth with 20% glycerol (v/v) at −80°C. Before each experiment, stock cultures were streaked on LB agar at 37°C for 18 h. A loopful of strain was inoculated into 30 mL of LB broth and incubated for 18 h with shaking at 130 rpm at 37°C. The biofilms were formed by culturing S. flexneri in 1/8 LB broth with 0.15% glucose (w/v) for 24 h.
Antibacterial activity and mechanism of OEO against planktonic S. flexneri cells
Minimum inhibitory concentration and minimum bactericidal concentration
The minimum inhibitory concentration (MIC) of OEO against the test S. flexneri was determined with the broth dilution method based on the Clinical and Laboratory Standards Institute guidelines (CLSI, 2009), with minor modifications. S. flexneri was diluted to a concentration of 3 × 108 CFU/mL with LB broth. Then, 200 μL bacterial suspension and different concentrations of OEO were added to the 96-well plate to get final concentrations of 0 (Control), 0.005% (v/v), 0.010%, 0.015%, 0.02%, 0.03%, 0.04%, 0.06%, and 0.08%. Samples were incubated at 37°C for 24 h, and the optical density (OD)600nm values were determined using a microplate reader (Model 680; Bio-Rad Laboratories, Hercules, CA, USA) at 0 and 24 h. To determine the minimum bactericidal concentration (MBC), 100 μL of each was plated on an LB agar plate and cultured for 48 h.
Growth curves
Growth curve assays were conducted as described by Zheng et al. (2019), with minor modifications. Each well of a 100-well plate was inoculated with 125 μL of diluted bacterial strain culture (∼2 × 106 CFU/mL). An equal volume of LB broth containing OEO was added to each well to achieve final concentrations equivalent to 0 (Control), 4 MIC, 2 MIC, MIC, 1/2 MIC, 1/4 MIC, 1/8 MIC, 1/16 MIC, 1/32 MIC, and 1/64 MIC. Samples were incubated in a fully automated Bioscreen C Plate Reader (Labsystems, Helsinki, Finland) at 37°C, and the OD600nm values were measured at 1 h interval for 48 h.
Antibacterial curve assay
The antibacterial curve assay was carried out to investigate the effect of OEO on the survival situation of S. flexneri as described in the previous report with some modifications (Wang et al., 2015). S. flexneri was adjusted to the concentration of ∼1 × 107 CFU/mL in LB broth, respectively, and treated with OEO at different concentrations of 0 (Control), MIC, 3/2 MIC, and 2 MIC. The S. flexneri cell survival situation in LB broth was monitored at 0, 1, 2, 4, and 6 h by counting the viable cells on the LB agar plates for 24 h at 37°C.
Inactivation effect of OEO on S. flexneri in minced pork
The experimental method of Juneja et al. (2021) was used to test the inactivation effect of OEO on S. flexneri in minced pork, with some modifications. The pork was put into a sterile sealed bag for irradiation sterilization and then thoroughly minced under sterile conditions. The S. flexneri diluted to an OD600nm of 0.5 in 0.1% Buffered Peptone Water (BPW) was mixed in the minced pork. Then, the OEO was added to the mixture to achieve final concentrations of 0 (Control), 2 (5 MIC), 4 (10 MIC), and 6 μL/g (15 MIC), respectively. The S. flexneri cell in pork was monitored at 0, 1, 2, 4, 6, and 8 h by counting the viable cells on the LB agar plates for 24 h at 37°C.
Reactive oxygen species
The effect of OEO on intracellular reactive oxygen species (ROS) level of S. flexneri was evaluated according to Kang's method (Kang et al., 2018a), with some modifications. The suspension of S. flexneri was prepared according to the Bacterial Strains and Growth Conditions section. Dichlorodihydrofluorescein diacetate (DCFH-DA; Beyotime Institute of Biotechnology, Shanghai, China) (final concentration 5 μM) was added to sterile phosphate-buffered saline (PBS; pH 7.2) and bacteria solution and then cultured at 37°C in dark for 10 min. OEO was added to samples to make the final concentration of 0 (Control), 1/4 MIC, 1/2 MIC, and MIC and incubated at 37°C in dark for 10 min. The samples were centrifuged (12,000 × g, 10 min, 4°C), washed twice, and resuspended with PBS. The fluorescence was measured at 488 and 525 nm, respectively, using a microplate reader (VictorX3; PerkinElmer, Waltham, MA, USA). Then, the samples were coated on LB agar plates and cultured for 24 h at 37°C before counting. The intracellular ROS level was calculated by using the following formula:
Confocal laser scanning microscopy
The effects of OEO on cell membrane integrity were verified by confocal laser scanning microscopy (CLSM) as described by Wang et al. (2019a), with some modifications. S. flexneri (∼3 × 108 CFU/mL) treated with OEO (0, MIC, 3/2 MIC, and 2 MIC) was incubated at 37°C for 10 min and then centrifuged at 10,000 × g for 5 min. The resulting bacterial pellets were resuspended in 0.85% NaCl (w/v) before stained with the bacterial live/dead stain in LIVE/DEAD® BacLight™ Bacterial Viability Kits (Thermo Fisher Scientific™, Waltham, MA, USA). Resuspended bacteria were mixed with 3 μL of fluorescent marker solution consisting of 1.5 μL of propidium iodide (PI) and 1.5 μL of SYTO 9 and incubated at room temperature for 5 min. Finally, sample was observed using a confocal laser scanning microscope (A1; Nikon, Tokyo, Japan) at 800 × magnification.
Field-emission scanning electron microscopy
Field-emission scanning electron microscopy (FESEM) was performed as described by Shi et al. (2016a), with some modifications. Bacterial cells (OD600nm = 0.5) were treated with OEO at concentrations equivalent to 0 (Control), MIC, 3/2 MIC, and 2 MIC and incubated at 37°C for 1 or 2 h. Cells were harvested, washed twice with PBS, fixed by sterile water containing 2.5% (v/v) glutaraldehyde overnight at 4°C, and then sequentially washed once with PBS and sterile water, respectively. The cells were dehydrated using a water–ethanol gradient (30%, 50%, 70%, 80%, 90%, 100%) for 10 min at room temperature before being fixed on FESEM supports. Finally, the samples were observed with a field-emission scanning electron microscope (S-4800; Hitachi, Tokyo, Japan).
Measurement of intracellular ATP concentrations
The intracellular ATP concentrations were measured following the previous report (Guo et al., 2019c), with some modifications. The cell solution (OD600nm = 0.5) dissolved in PBS was taken into a test tube for treating with OEO (0, MIC, and 2 MIC) at 37°C for 15 min. The bacterial cells were disrupted by ultrasonic wave for 7 min on ice to extract the ATP. The samples were immediately placed in a dry bath at 100°C for 2–3 min and centrifuged for 5 min at 8000 × g to collect supernatant. Intracellular ATP was measured by using an ATP assay kit (Beyotime Bioengineering Institute, Shanghai, China). After adding 100 μL of ATP assay mix to 100 μL of supernatant in 96-well white microtiter plates (Costar; Corning, New York, NY, USA), the samples were measured by applying a microplate reader (VictorX3).
Membrane potential determinations
With some modifications, the method described by Shi et al. (2016b) was followed. Cells were diluted with PBS and adjusted to the concentration of ∼3 × 108 CFU/mL. Afterward, 125 μL of cell suspensions was placed in 96-well black microtiter plates (Costar; Corning), and then, 1 μM of the membrane potential-sensitive fluorescent probe bis-(1, 3-dibutylbarbituric acid) trimethine oxonol [DiBAC4(3); Molecular Probes; Sigma, Louis, MO, USA] was added and incubated for 30 min at 37°C. After cultivation, 125 μL of OEO at three concentrations (0, MIC, and 2 MIC) was added to the corresponding sample. The fluorescence of the sample and its background fluorescence were measured on a microplate reader (VictorX3) at excitation and emission wavelengths of 485 and 515 nm, respectively, continuously for 30 min and at intervals of 5 min.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
Intracellular soluble proteins of S. flexneri were analyzed using sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) following the procedures as mentioned in the previous report (Zheng et al., 2019). S. flexneri prepared as described in the Minimum Inhibitory Concentration and Minimum Bactericidal Concentration section was treated with OEO (0, MIC, 3/2 MIC, and 2 MIC) at 37°C for 8 h. The samples were centrifuged at 10,000 × g for 5 min, washed twice with PBS, and the cell pellet was collected.
The lysate preparation was added into the cell pellet treated for 30 min on ice and then treated for 40 min at 37°C with shaking. Next, the supernatant was collected and the protein concentration was determined. The protein concentrations of all samples were adjusted to be consistent, and the 25 μL of 5 × SDS loading buffer was added into 100 μL of samples. The samples were boiled for 5 min and cooled at room temperature and then analyzed by SDS-PAGE. After electrophoresis, the protein bands were stained with Coomassie brilliant blue R-250 and then decolorized to obtain the separated protein bands.
The inhibitory activity of OEO on mature biofilm of S. flexneri
Assessment of biofilm biomass
The effect of OEO on mature biofilm biomass of S. flexneri was tested in 96-well microtiter plates as described by Jadhav et al. (2013), with little modification. Briefly, S. flexneri cells resuspended in 1/8 LB broth with 0.15% (w/v) glucose and adjusted to the concentration of ∼3 × 108 CFU/mL in 96-well culture plates at 37°C for 24 h (200 μL per well). After 24 h of biofilm formation, the absorbance of each well was measured at 630 nm. Then, the suspension was removed gently, and then, 200 μL of 1/8 LB broth with 0.15% glucose containing the OEO at concentrations equivalent to 0 (Control), MIC, 2 MIC, 4 MIC, and 8 MIC was added to each well, incubated at 37°C for 2, 4 and 6 h, respectively.
The suspension was removed gently, and the wells were washed with PBS, and then, the plates were air-dried before staining with 200 μL 1% (w/v) crystal violet solution for 10 min at room temperature. After the wells were washed and air-dried, the combined dye was solubilized in 200 μL of 33% (v/v) glacial acetic acid, and the plates were shaken for 20 min at room temperature. Finally, the absorbance of each well was measured by applying a microplate reader (Model 680; Bio-Rad Laboratories) at 570 nm. The Specifie Biofilm Formation (SBF) was calculated by using the following formula: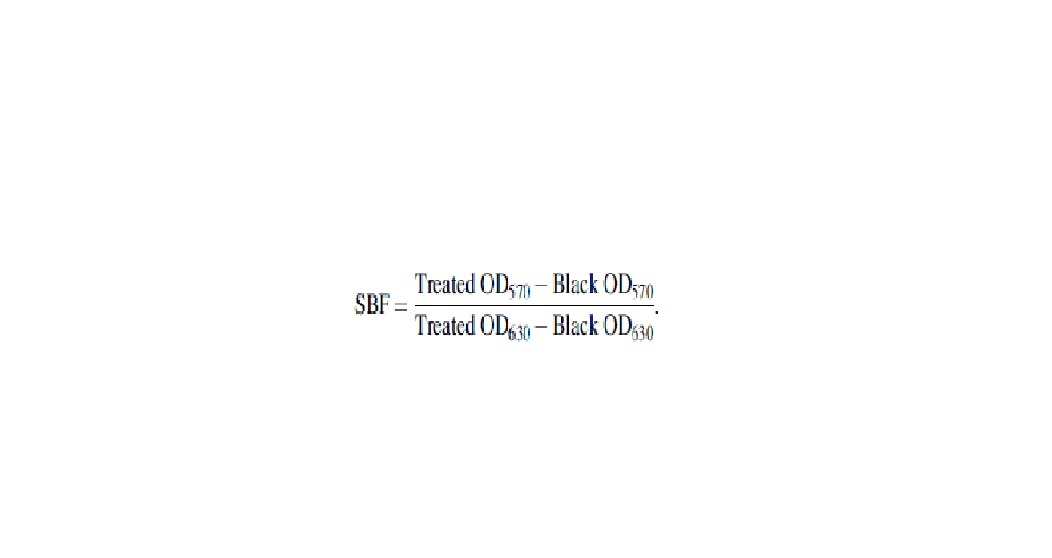
FESEM assay
The effect of OEO on S. flexneri biofilms was observed by FESEM as described by Gowrishankar et al. (2015), with some modifications. S. flexneri cells (OD600nm = 0.5) were added to the wells of 24-well tissue culture plates (2 mLper well) with embedded stainless steels (1 × 1 cm) at 37°C for 24 h. Then, the suspension was removed gently, and 2 mL of 1/8 LB broth with 0.15% glucose containing the OEO at concentrations equivalent to 0 (Control), MIC, 2 MIC, and 4 MIC was added to each well to inoculate for 2 and 4 h at 37°C. Next, the biofilms on stainless steels were fixed for 12 h at 4°C in a solution containing 2.5% (v/v) glutaraldehyde. The stainless steels were washed in PBS, washed with distilled water, and dehydrated using a water–ethanol gradient series of 30%, 50%, 70%, 80%, 90%, and 100% for 10 min at room temperature. Finally, the samples were observed by FESEM (S-4800; Hitachi).
CLSM assay
Biofilms of S. flexneri were analyzed using CLSM following the procedures as mentioned in the previous report (Peng et al., 2018). Briefly, the mature biofilm of S. flexneri was formed and treated with OEO as described in the Assessment of Biofilm Biomass section. The stainless steels were stained with Con A-FITC for 15 min at 4°C, immobilized by 2.5% (v/v) glutaraldehyde for 2 h at 4°C, and then stained with Hoechst 33258 for 20 min at room temperature. Thereafter, stainless steels with biofilms were observed using a confocal laser scanning microscope (A1; Nikon) at 400 × magnification, with an excitation wavelength of 488 nm (Con A-FITC) or 405 nm (Hoechst 33258).
Statistical analyses
All experiments were carried out in triplicate. Statistical analyses were carried out using IBM SPSS Statistics (version 20.0; IBM Corp., Armonk, NY, USA). Data are presented as the mean ± standard deviation (SD) (n = 3). Differences between means were tested by the Student t-test. p < 0.05 and p < 0.01 were considered statistically significant and extremely significant, respectively.
Results
Antibacterial activity and mechanism of OEO against planktonic S. flexneri cells
MIC and MBC
The OEO had an inhibitory effect on S. flexneri. The MIC of OEO against S. flexneri was 0.02% (v/v), and the MBC of OEO against S. flexneri was 0.04% (v/v).
Effects of OEO on S. flexneri growth
The effects of OEO at concentrations ranging from 1/32 MIC to 2 MIC on the growth of S. flexneri were determined by growth curve assay (Fig. 1). The control group without OEO treatment exhibited a typical growth curve including lag, exponential, and stabilization phases. When the concentration of OEO decreased to <1/4 MIC, there was no apparent effect on the growth of S. flexneri. S. flexneri cultures treated with OEO at 1/2 MIC showed a longer lag phase and a decrease in the maximum growth rate. The growth of S. flexneri was completely inhibited by OEO at MIC and 2 MIC.
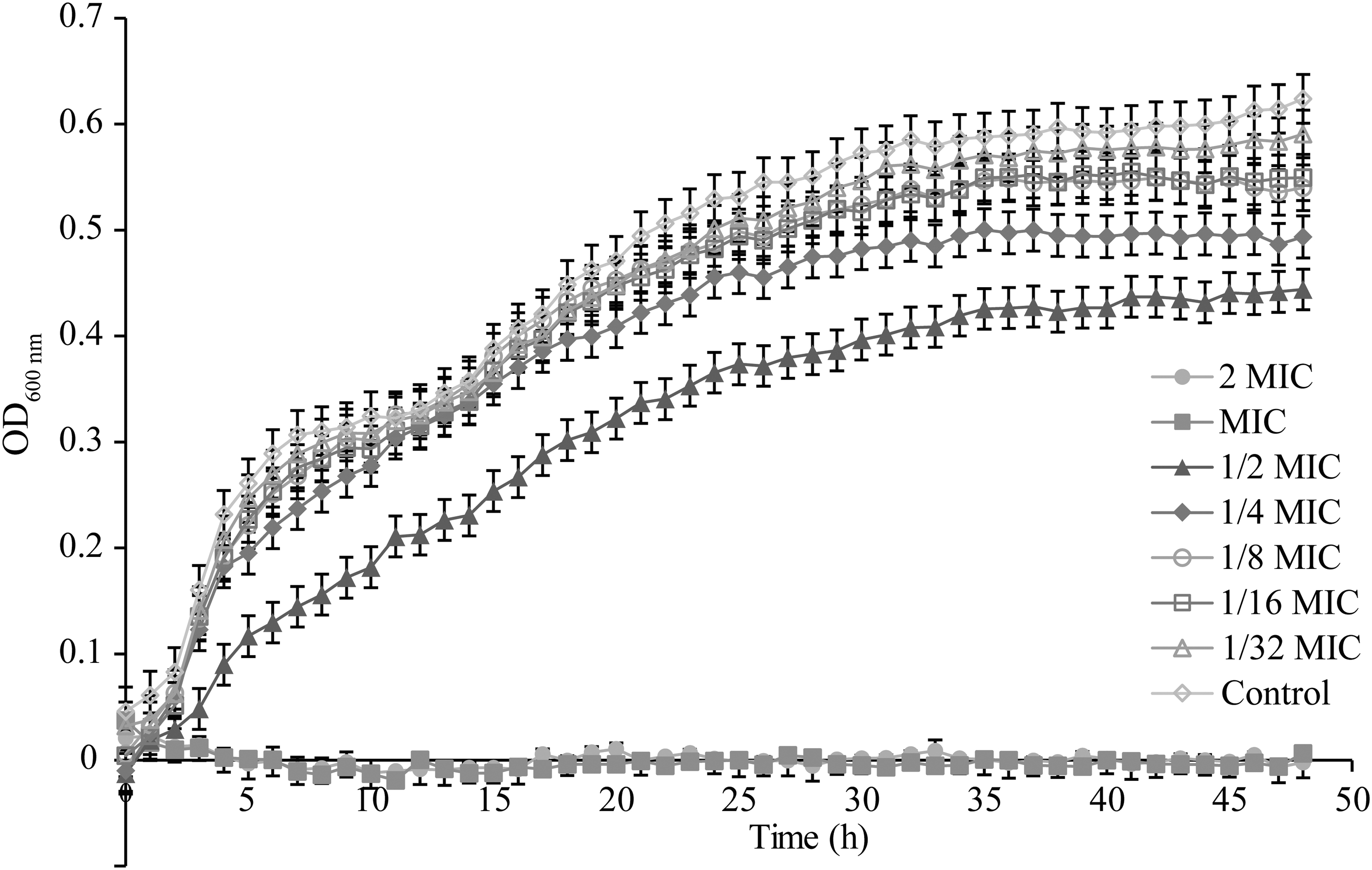
Growth curve analysis of Shigella flexneri ATCC 12022 cultured in LB broth with various concentrations of OEO. LB, Luria–Bertani; MIC, minimum inhibitory concentration; OD600, optical density at 600 nm; OEO, oregano essential oil.
Antibacterial curve assay
Antibacterial curve assay is shown in Figure 2. The initial value of S. flexneri cells in LB broth for all treatment groups was 7 log CFU/mL. The cells without OEO (Control) approximately increased to 8.4 log CFU/mL at 6 h. When S. flexneri was treated with OEO at the MIC, the viable cell quantities remained steady after 6 h, whereas the population of S. flexneri that was treated with OEO at the 3/2 MIC and 2 MIC decreased to an undetectable level (1 CFU/mL) after 4 and 2 h, respectively.
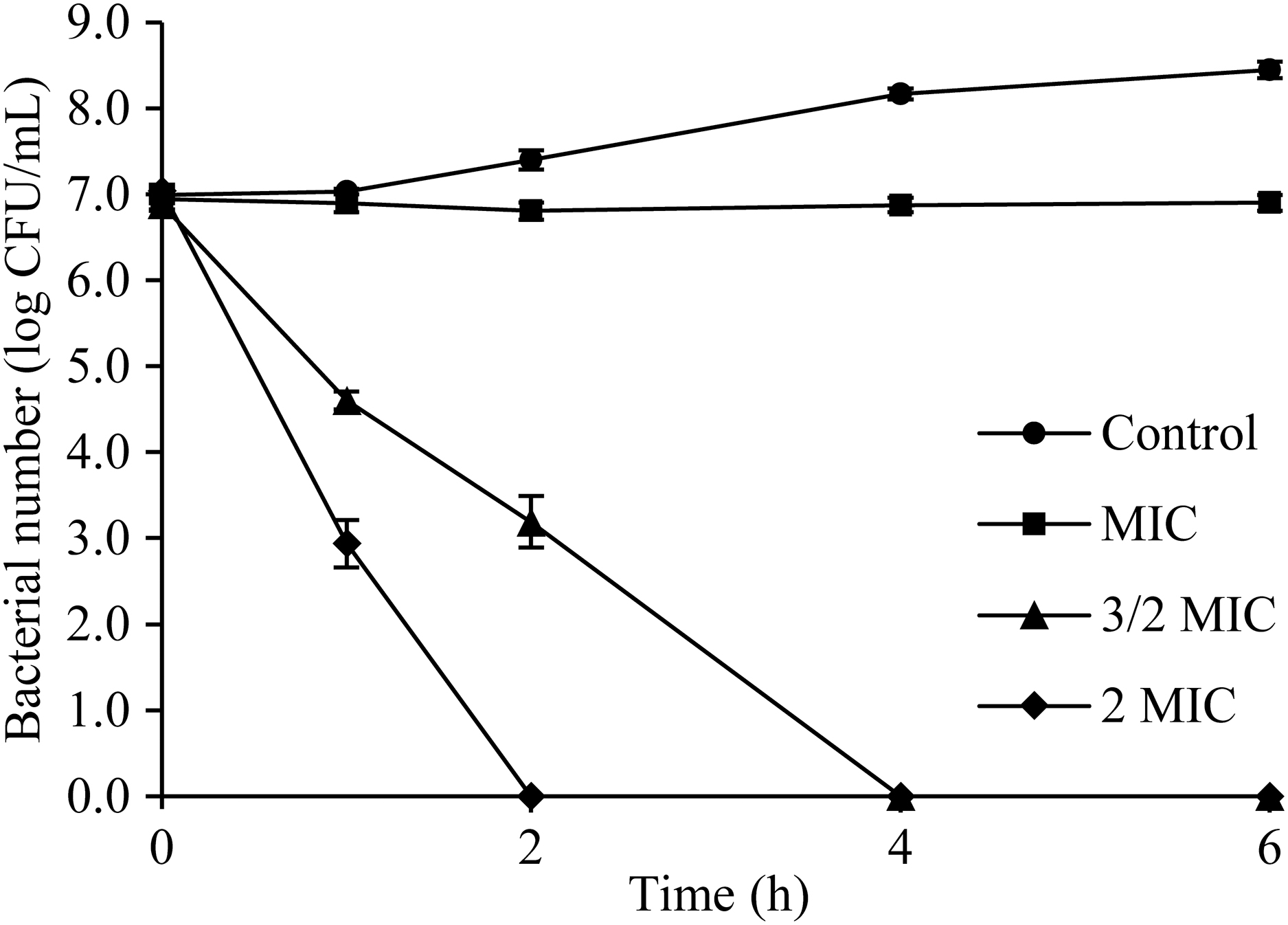
Time–kill curves of OEO against Shigella flexneri ATCC 12022 cells in LB. LB, Luria–Bertani; OEO, oregano essential oil.
Inactivation effect of OEO on S. flexneri in minced pork
The changes in the quantity of S. flexneri in minced pork treated with different concentrations of OEO are shown in Figure 3. The initial value of S. flexneri cells was the same for all the treatment groups (∼7 log CFU/g). The cells without OEO (Control) approximately increased to 8.3 log CFU/g at 12 h. When S. flexneri was treated with OEO with the concentration of 5 MIC, compared with the control, the viable cells grew after 6 h. Furthermore, OEO with the concentration of 10 MIC led to about 2.5 log CFU/g reductions of the S. flexneri population, whereas the population of S. flexneri that was treated with OEO with the concentration of 15 MIC decreased to an undetectable level after 9 h.

Time–kill curves of OEO against Shigella flexneri ATCC 12022 cells in minced pork. OEO, oregano essential oil.
Reactive oxygen species
The intracellular ROS levels of S. flexneri were significantly (p < 0.01) increased with OEO treatment in a dose-dependent manner (Fig. 4). The intracellular ROS level of S. flexneri without OEO treatment was 10.340 ± 2.800. And the intracellular ROS concentration treated with 1/4 MIC and 1/2 MIC increased to 29.835 ± 6.420 and 41.851 ± 12.971, respectively.

Effect of OEO on ROS concentration in Shigella flexneri ATCC 12022. Values represent the means of independent triplicate measurements. Bars represent the SD (n = 3). **p < 0.01. OEO, oregano essential oil; ROS, reactive oxygen species; SD, standard deviation.
Cell membrane integrity
Cell membrane integrity was observed by CLSM with results presented in Figure 4. The SYTO 9 stain generally labels all bacteria in a population. In contrast, PI penetrates only bacteria with damaged membranes, causing a reduction in the SYTO 9 stain fluorescence when both dyes are present. Thus, with an appropriate mixture of the SYTO 9 and PI stain, bacteria with intact cell membranes stain fluorescent green, whereas bacteria with damaged membranes stain fluorescent red. At the initial treatment, control cells appeared almost completely green, indicating that the cell membranes of untreated cells were intact (Fig. 5A, E).
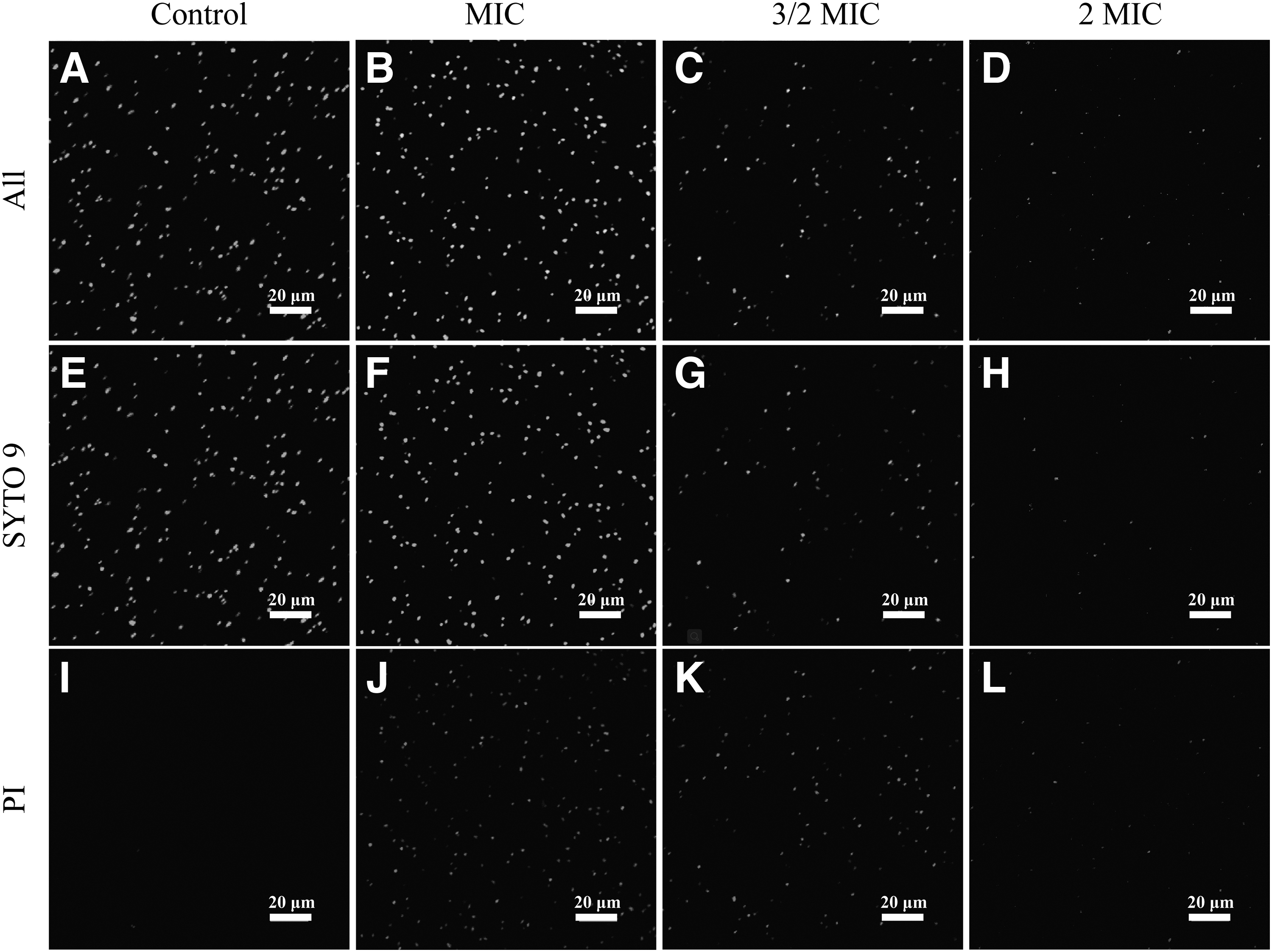
CLSM analysis of
Compared with the control, cells that were treated with OEO at MIC concentrations emitted few red fluorescence, reflecting that the cell membranes had been disrupted (Fig. 5B). With an increase in OEO treatment concentration to 3/2 MIC (Fig. 5C) and 2 MIC (Fig. 5D), the amount of fluorescence in red is obviously more than that in green, indicating that the cell membrane was seriously damaged and suggesting that OEO destroys the integrity of the S. flexneri cell membrane in a concentration-dependent manner.
Cell morphology
FESEM was used to evaluate the changes of OEO against the S. flexneri cell morphology, as shown in Figure 6. The S. flexneri cells were untreated with OEO, with a smooth and rod-shaped appearance (Fig. 6A, E). Compared with the control, cells in all treated showed (Fig. 6B–D, F–H) seriously morphological changes with a wrinkle and rough surface. The degree of cell shrinkage increased as the concentration of OEO and incubation time increased.
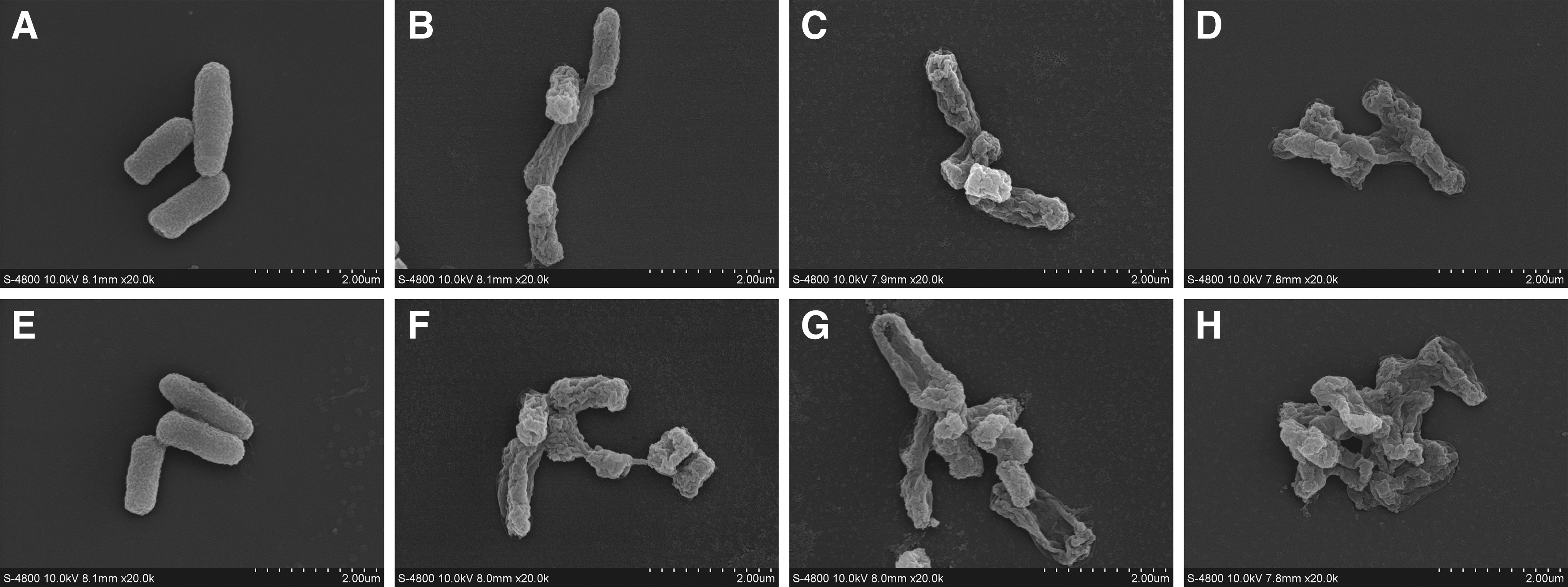
Field-emission scanning electron micrographs of Shigella flexneri ATCC 12022.
Intracellular ATP concentrations
There is a decrease of intracellular ATP concentrations of S. flexneri after the treatment with OEO (Fig. 7). Comparing control cells with cells treated with OEO at MIC and 2 MIC, there were significant reductions (p < 0.01) of intracellular ATP in the treated cells. The intracellular ATP concentration of S. flexneri without OEO treatment was 0.190 ± 0.015 μM. And OEO at MIC and 2 MIC, the intracellular ATP concentration decreased to 0.107 ± 0.015 and 0.004 ± 0.009 μM, respectively.
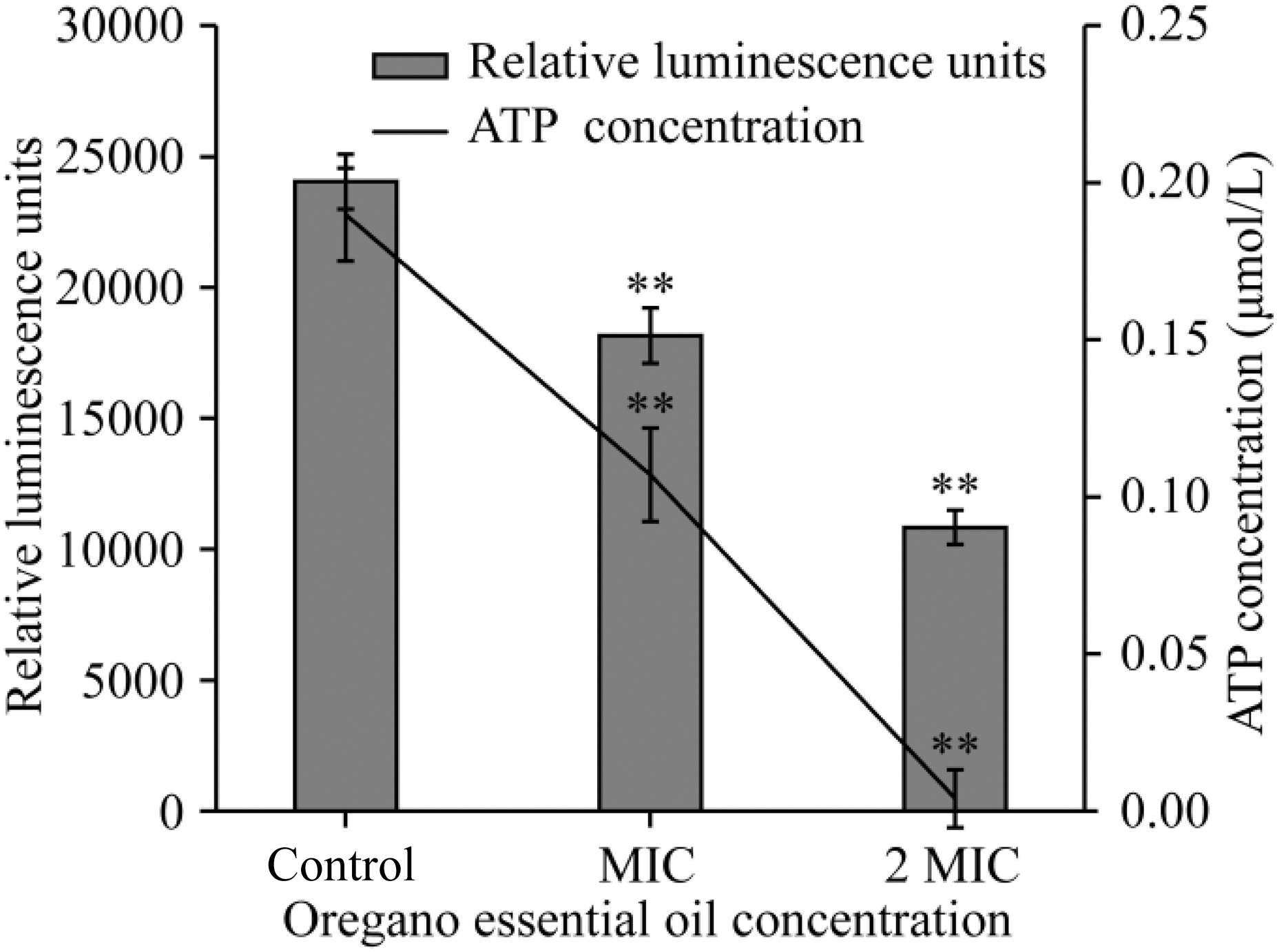
Effects of OEO on intracellular ATP production by Shigella flexneri ATCC 12022. Values represent the means of independent triplicate measurements. Bars represent the SD (n = 3). **p < 0.01. OEO, oregano essential oil.
Membrane potential
As shown in Figure 8, S. flexneri cells treated with OEO displayed prominent cell membrane hyperpolarization, as evidenced by a significant decrease in fluorescence (p < 0.01). The fluorescence of S. flexneri cells treated with OEO at MIC remained almost unchanged. While the fluorescence of cells continued to decrease at 2 MIC in 15–30 min, indicating that the degree of hyperpolarization increased.
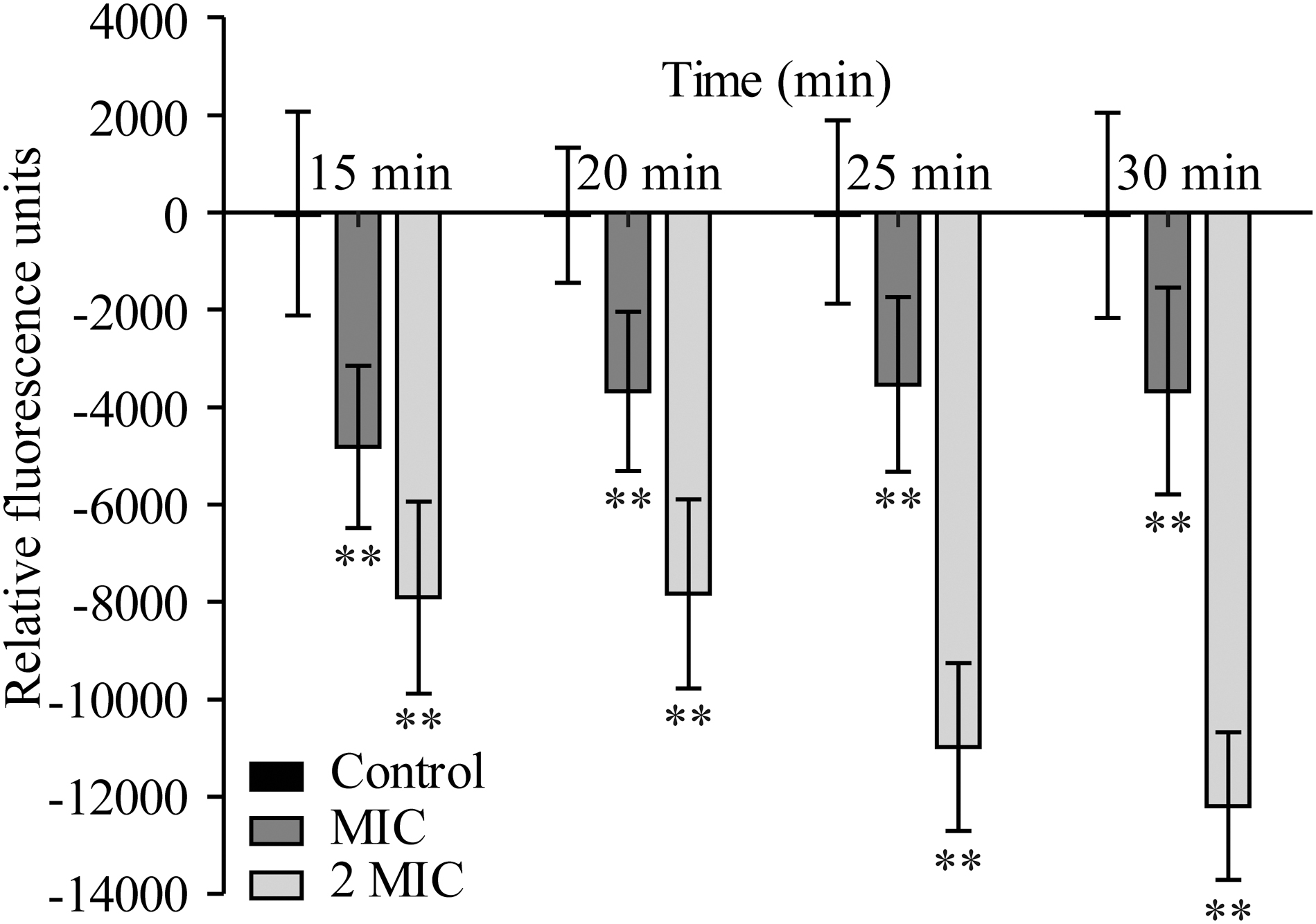
Effects of OEO on the membrane potentials of Shigella flexneri ATCC 12022. Values represent the means of independent triplicate measurements. Bars represent the SD (n = 3). **p < 0.01. OEO, oregano essential oil.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
The results of SDS-PAGE analysis of OEO against intracellular soluble proteins are shown in Figure 9. Compared with the control, the protein bands of S. flexneri treated with MIC of OEO were not significantly different. The bands (∼60 kDa) significantly changed in protein bands treated with OEO at 3/2 MIC and 2 MIC. Moreover, there were more kinds and amounts of bands below 35 kDa in the control than treated with OEO at 3/2 MIC and 2 MIC.

SDS-PAGE analysis of intracellular soluble proteins of Shigella flexneri ATCC 12022 treated with OEO. Lane Maker: marker; Control: control for 8 h; MIC: treated samples for 8 h by OEO at MIC; 3/2 MIC: treated samples for 8 h by OEO at 3/2 MIC; 2 MIC: treated samples for 8 h by OEO at 2 MIC. MIC, minimum inhibitory concentration; OEO, oregano essential oil; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis.
The inhibitory activity of OEO on mature biofilm of S. flexneri
Assessment of biofilm biomass
S. flexneri mature biofilm was treated with OEO concentration MIC at 4°C (Fig. 10A) and 2 MIC at 25°C (Fig. 10B) and 37°C (Fig. 10C) for 2 and 4 h, respectively (Table 1 and Fig. 10). The results showed that, compared with the control group without OEO treatment, different concentrations of OEO had significant scavenging effect on the mature biofilm of S. flexneri in a time-dependent and concentration-dependent manner. Meanwhile, the scavenging effect is different at different treatment temperatures: At 37°C, the biofilm was treated with MIC and 2 MIC OEO for 2 h, the SBF values were 1.37 ± 0.25 and 0.72 ± 0.15 and the clearance rates were 60% and 79%. After 4 h treatment, the SBF values were 0.93 ± 0.32 and 0.18 ± 0.11, the clearance rates were 75% and 95%, and the removal effect is the best. At 4°C, the SBF values were 1.47 ± 0.36 and 1.18 ± 0.24 and the clearance rates were 39.33% and 51.19% after 2 h treatment; the SBF values were 2.58 ± 0.53 and 2.21 ± 0.34 after 4 h treatment, the clearance rates were only 9.13% and 22.21%, and the clearance effect is poor.
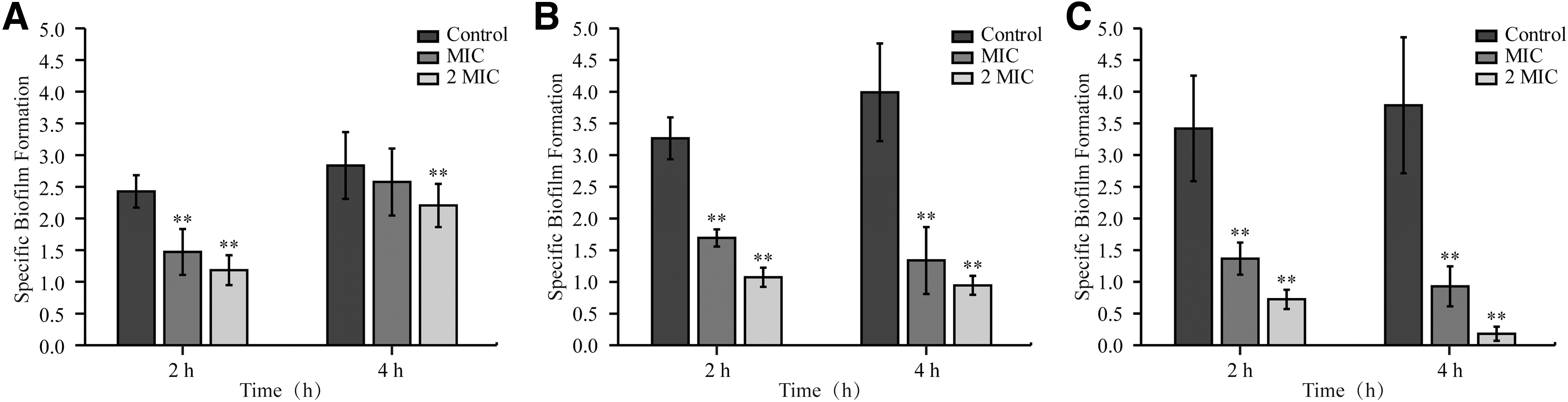
Effect of OEO on the mature biofilm of Shigella flexneri ATCC 12022 at 4°C
Effect of Oregano Essential Oil on the Mature Biofilm of Shigella flexneri ATCC 12022
p < 0.01.
MIC, minimum inhibitory concentration; SBF, Specifie Biofilm Formation.
FESEM assay
The effects of OEO treatment with MIC (Fig. 11B, E) and 2 MIC concentrations (Fig. 11C, F) on the stereoscopic structure of biofilm of S. flexneri on stainless steel surface were observed by FESEM. The results showed that the biofilm of S. flexneri without OEO treatment (Fig. 11A, D) had a dense structure and obvious three-dimensional morphology of adhesion. After the treatment of OEO, the structure of the biofilm obviously became loosened. Meanwhile, the bacterial morphology was shrunk, and the removal effect increased with the increase of the concentration of OEO. It can be seen that OEO can not only change the stereoscopic structure of the biofilm but also change the morphology of the bacteria in the biofilm and further enhance the scavenging effect of the biofilm from two aspects.

Effects of OEO on the mature biofilm of Shigella flexneri ATCC 12022 on the stainless steel surfaces at 25°C.
CLSM assay
Figure 12 shows the changes of S. flexneri biofilm treated with OEO. With the increase of OEO concentration, the amount of S. flexneri biofilm and bacteria decreased significantly. After dye staining, the biofilm was observed as green and S. flexneri as blue under laser scanning confocal microscope. The control bacteria were abundant and dense, and the biofilms were closely connected. In contrast to the control, the amount of the biofilm and bacteria treated with MIC and 2 MIC of OEO decreased, and the biofilm was dispersed in stainless steel surfaces. As the concentration of OEO increased, the phenomenon becomes more obvious, which indicates that OEO has obvious inhibitory effect on S. flexneri.

Effects of OEO on the EPS structure of Shigella flexneri ATCC 12022 mature biofilm on the stainless steel surfaces at 25°C.
Discussion
In this study, OEO was shown to exhibit inhibitory activity against S. flexneri, with MIC of 0.02% (∼0.189 mg/mL) and MBC of 0.04% (∼0.376 mg/mL). In previous studies, the inhibitory effect of some essential oils on S. flexneri was also investigated. Chimnoi et al. (2018) determined that the MIC and MBC values of essential oil of fresh leaves of Ocimum gratissimum against S. flexneri were 1 and 2 mg/mL, respectively. Noshad et al. (2018) reported that the MIC and MBC of the black zira essential oil against S. flexneri were 2 and 4 mg/mL, respectively. Therefore, compared with other reported essential oils, OEO showed stronger antibacterial activity against S. flexneri. Chan et al. (2018) determined that the MIC of carvacrol, >60% of the total ingredients of OEO, against S. flexneri was 0.313 mg/mL (Cui et al., 2019). The antibacterial effect of OEO is stronger than carvacrol, which may be caused by the synergistic antibacterial action from various components of OEO. In future studies, we will try to isolate and mix other against S. flexneri strains to evaluate the effect of OEO on the mixing of different strains.
In the antibacterial curve assay, the population of S. flexneri in LB broth that was treated with OEO at the 3/2 MIC and 2 MIC decreased to an undetectable level (1 CFU/mL) after 4 and 2 h, respectively (Fig. 2). Similarly, Chen et al. (2021) confirmed that the survival of S. flexneri treated with 2 MIC decreased from 6.0 to 0.11 log CFU/mL after 4 h. Kang et al. (2018a) reported that the quantities of S. flexneri treated by gallic acid at MBC decreased from 6.35 log CFU/mL to an undetectable level after 4 h. The population of S. flexneri in minced pork that was treated with OEO of 15 MIC decreased to an undetectable level (1 CFU/mL) after 9 h (Fig. 3).
Therefore, it can be found that the sterilization effects of OEO against S. flexneri were more pronounced in LB broth than in minced pork. The previous research reported that Zataria multiflora Boiss. essential oil played a better role in inactivating Listeria monocytogenes in broth than in the minced fish mediums (Pilevar et al., 2019). This may be attributed to the fact that the structure and nutrients of the meat inhibit the effectiveness of natural bacteriostatic agent. To improve the bacteriostatic effect of OEO in meat, methods such as making antibacterial films incorporated with OEO and encapsulating it into microcapsules to increase solubility can be further investigated.
In this study, the intracellular ROS level of S. flexneri increased in a concentration-dependent manner after treatment with OEO (Fig. 4). Similarly, Das et al. (2016) demonstrated that the existence of eugenol increased the ROS level in S. aureus and speculated the inhibitory ability of the eugenol involved in the generation of intracellular ROS. Therefore, the rise of ROS level in S. flexneri treatment with OEO may be construed as the increasing ROS generation. According to existing studies, excessive ROS production above basal levels can impair and oxidatively damage lipids to damage cell membranes (Fatma et al., 2013). Therefore, OEO induced intracellular ROS within cells, which may be of prime importance and be an indispensable reason for its antibacterial effect.
The fluorescent dye PI and SYTO 9 were used to investigate the effects of OEO on S. flexneri cell membrane integrity and proved that OEO damaged the cell membrane of S. flexneri (Fig. 5). In the previous study, Kang and Song (2018) confirmed that cinnamon leaf oil had destructive effects on the cell membrane integrity of L. monocytogenes, E. coli, and S. enteritidis by CLSM. Guo et al. (2019a) found that some green fluorescence of the Cronobacter sakazakii treated with CoQ0 was obscured by red fluorescence and that the cell membrane was destroyed. The damage of membrane integrity may be interpreted that OEO changes membrane functioning and influences protein-to-lipid ratios in the membrane (Ornella et al., 2018). Combined with the result of ROS, an emerging argument is that the increasing ROS in S. flexneri treated by OEO can oxidize lipid molecules on the cell membrane to damage the membrane integrity (Fu et al., 2021).
The FESEM images showed that OEO caused cell morphological alterations of S. flexneri, but no cell ruptured was found (Fig. 6). Similarly, Cui et al. (2019) found that the number of bacterial cells significantly reduced and some of the surface of the bacteria collapsed and shrunk after treatment with OEO. In addition, Chimnoi et al. (2018) found that the aggregation of the cell envelope and abnormal cell breaking were clearly seen in S. flexneri, which was treated with essential oil of fresh leaves, and that the alterations may be caused by an impaired membrane structure and leakage of intracellular dense material of treated. According to the results of CLSM, it can be inferred that the membrane damage of S. flexneri treated by OEO leads to the morphological changes by the loss of cell contents, which may cause irreversible damage or even death of the bacteria.
The results of this study showed that OEO significantly reduced the intracellular ATP concentration in S. flexneri (p < 0.01) compared with the control (Fig. 7). In previous study, L. monocytogenes treated with 0.31 and 1.25 mg/mL olive oil polyphenol extract showed that the intracellular ATP concentration decreased by 50.85% and 95.76%, respectively, compared with the control (Guo et al., 2019b). Kang et al. (2019) found that the intracellular ATP concentration of S. aureus treated with peppermint essential oil was significantly decreased (p < 0.05). Existing studies have concluded that the decrease in intracellular ATP concentration may be due to rapid ATP hydrolysis, which is manifested in the inhibition of ATP synthesis in bacterial cells or the increase of proton pump hydrolysis rate of ATP under the action of natural active substances, or to intracellular ATP leakage through the cell membrane by changes in membrane permeability (Mempin et al., 2013).
Through the membrane potential assay, it could be found that OEO caused the hyperpolarization of S. flexneri membrane (Fig. 8). Similarly, Yang et al. (2021) showed that the membrane potential of Salmonella typhimurium cells treated with CoQ0 indicated membrane hyperpolarization as a result of a decrease in membrane potential. Moreover, OEO was determined that caused leakage of phosphate ions from S. aureus and hyperpolarization of cell membrane (Lambert et al., 2010). The hyperpolarization of cell membrane potential may be due to diffusion of external K+ (Guo et al., 2019a).
The SDS-PAGE analysis showed that after different concentrations of OEO treatments, some S. flexneri protein bands were strong and clear, whereas some became shallow and even disappeared (Fig. 9). Meng et al. (2016) found that Juniperus rigida essential oil had a remarkable inhibitory effect on bacterial proteins of Klebsiella pneumoniae, and especially, the protein band with molecular weight <42.0 kDa got much fainter. Wang et al. (2015) reported that after lactic acid treatment, protein bands in Salmonella Enteritidis, E. coli, and L. monocytogenes weakened or even disappeared by destroying or inhibiting their synthesis.
In addition, Di Pasqua et al. (2010) showed that many proteins can be either upregulated or downregulated by the presence of thymol, with significant changes in proteins belonging to different functional classes. In the presence of low concentrations of essential oils, bacteria react by increasing their expression of the stress–response proteins to repair the damaged proteins (Lambert et al., 2010). Thus, we speculated that the OEO could probably have an effect on the cellular proteins by regulating to destroy some protein bands or producing stress proteins.
The results (Fig. 10) showed that the OEO had a good scavenging effect on the mature biofilm of S. flexneri in a time-dependent and concentration-dependent manner. Agarwal et al. (2008) reported that eucalyptus essential oil, mint essential oil, ginger grass essential oil, and clove essential oil on the biofilm reduction rate of Candida albicans was 80.87%, 74.16%, 40.46%, and 28.57%, respectively. Similarly, after OEO treatment, the biofilm of S. flexneri formed on the surface of food processing of stainless steel was loose and the bacterial morphology change was observed by FESEM (Fig. 11). Guo et al. (2019b) found that the essential oil from Citrus Changshan-huyou Y. B. destroyed the integrity of L. monocytogenes biofilm and made the bacteria to die achieving the removal of the biofilm.
Similarly, Lira et al. (2020) reported that OEO significantly decreased Salmonella in mature biofilm on stainless steel surfaces; meanwhile, the bacterial morphology was destroyed. Therefore, through the above research, we believe that OEO has a significant scavenging effect on bacterial biofilm by killing bacteria or destroying the biofilm structure and has good application potential for the removal of biofilm in food processing environment. In particular, the results (Fig. 10) that scavenging of mature biofilm was the most significant at 37°C, followed by 25°C, and the worst at 4°C.
Nostro et al. (2015) found that the polyethylene film containing OEO had the strongest bactericidal effect at 37°C, whereas the bactericidal rate was lower at a lower temperature. Previous studies have shown that the bacteria will develop the cross-protection to various stresses to survive and grow under favorable conditions and regain the ability to tolerate further severe stresses (Wang et al., 2019b). Therefore, we speculated that the possible reason for this temperature-dependent difference is that the biofilm increases its tolerance to cold when exposed to low temperatures, thus reducing the effect of OEO. However, the mechanism of OEO on biofilm of removal needs further study and confirmation.
Exopolysaccharide, as an important component of extracellular polymeric substance (EPS), protects the bacteria from external pressure by surrounding it (Kang et al., 2018a). In this study, the amount of S. flexneri and polysaccharides on the stainless steel plate could be effectively reduced by OEO (Fig. 12). Similar to the results, Yang et al. (2020) reported the green fluorescence (biofilm polysaccharide) and blue fluorescence (bacterial cells) were obviously reduced in the P. aeruginosa biofilm samples after the illumination of LED and speculated that polysaccharides in biofilms suffered oxidative damage from ROS, affecting the structure and functions of the biofilms. In the present study, the observation of CLSM may be interpreted that inactivation of S. flexneri treated with OEO resulted in exfoliation of surrounding EPS including extracellular polysaccharides. Furthermore, according to the results of ROS, the accumulative ROS may oxidatively damage polysaccharides in biofilms from S. flexneri treated by OEO.
Conclusions
In the present study, OEO demonstrated a good activity of inhibiting and killing S. flexneri and effectively restrained growth rate. Owning several mechanisms of action, OEO increased the ROS level, decreased cell membrane integrity, destroyed cell morphology, reduced intracellular ATP concentration, made the cell membrane depolarized, affected the content of intracellular soluble protein, and caused normal physiological function disorder. Furthermore, OEO effectively inactivated S. flexneri cells in mature biofilm on stainless steel surfaces at 4°C, 25°C, and 37°C, destroyed the dimensional structure of the biofilm, and reduced cells and polysaccharides in biofilm. In conclusion, OEO exerts its antibacterial action partly and also has a good scavenging effect on its biofilm and could be used in food production and processing environments to effectively control S. flexneri.
Footnotes
Acknowledgment
We thank Dr. Zhang Guoyun (State Key Laboratory of Crop Stress Biology for Arid Areas, Northwest A&F University, Yangling, China) for FESEM experimental assistance. We thank Ms. Ma Zhenzhen (Crop Biology Innovation Center, College of Agriculture, Northwest A&F University, Yangling, China) for assistance with the experiments.
Authors' Contributions
T.C.: Conceptualization, methodology, investigation, formal analysis, project administration, writing—original draft, writing—review and editing. Z.L.: Conceptualization, methodology, formal analysis, project administration, writing—original draft, writing—review and editing. P.G.: Resources, investigation, data curation, visualization. J.G.: Resources, methodology, investigation. R.W.: Resources, methodology. D.G.: Formal analysis, resources. J.Y.: Supervision, software. X.L. and X.X.: Supervision, project administration. C.S.: Project administration, supervision, funding acquisition, writing—review and editing. All authors have read and agreed to the published version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (32272445 and 31801659) and the class General Financial Grant from the Shaanxi Province Postdoctoral Science Foundation (2018BSHEDZZ150).