
Abstract
Enterococci can act as reservoirs for antibiotic-resistant genes that are potentially at risk of being transferred to other bacteria that inhabit in the gastrointestinal tract. The aim of this study was to determine the phenotypic and molecular characteristics of antibiotic-resistant enterococci isolated from probiotic preparations. In total, we isolated 15 suspected Enterococcus species from 5 compound probiotics, which were identified by 16S rDNA as 12 Enterococcus faecium and 3 Enterococcus faecalis. Determination of antimicrobial susceptibility by the microdilution broth method showed widespread resistance to sulfamethoxazole (100%), norfloxacin (99.3%), azithromycin (99.3%), gentamicin (86.7%), and chloramphenicol (20%). Whole genome sequencing of five resistant strains revealed that all had circular DNA chromosomes and that E. faecium J-1-A to J-4-A contained a plasmid, while E. faecalis J-5-A did not. The results of the resistance gene analysis revealed that each strain contained approximately 30 resistance genes, with the antibiotic resistance genes and the multidrug resistance efflux pump genes mdtG, lmrC, and lmrD detected in all strains. The chloramphenicol resistance genes ykkC and ykkD were first identified in E. faecalis. And there were 21, 19, 21, 21, and 29 virulence factors involved in strains, respectively. Further analysis of the gene islands (GIs) revealed that each strain contained more than 10 GIs. The above results confirm the existence of hidden dangers in the safety of probiotics and remind us to carefully select probiotic preparations containing enterococcal strains to avoid the potential spread of resistance and pathogenicity.
Introduction
Nowadays, antibiotic multidrug resistance (AMR) has become a global concern. Infections caused by AMR have emerged as a new challenge for anti-infective therapies and are one of the major threats to human health and life. Enterococci are Gram-positive (G+) bacteria, which are widely distributed in the natural environment and in the digestive tract of humans and animals (Sadowsky et al., 2011). But the accumulation of antibiotic resistance in enterococci, particularly the emergence of vancomycin-resistant strains due to inappropriate use of antibiotics, has caused great difficulties in clinical treatment (Hammad et al., 2022). And they may act as donors or reservoirs of antibiotic-resistant genes that are potentially at risk of being transferred to other bacteria that inhabit the gastrointestinal tract (Ramos et al., 2020).
Probiotics are live microorganisms which when administered in adequate amounts confer a health benefit on the host. The main strains of probiotics are lactobacilli and bifidobacteria (Hill et al., 2014). In recent years, research on the relationship between intestinal microecology and various diseases has gained popularity. Enterococcus faecalis and Enterococcus faecium have been listed as approved probiotics for human use in China, but no mention has been made of potential harm caused by resistance to these probiotics (Chinese Society of Microecology et al., 2020). The reverse is true in other countries. Probiotics containing of enterococci have been questioned due to the lack of evidence-based medicine and the safety of their application. The FAO/WHO guidelines for the Evaluation of Food Probiotics issued in 2002 pointed out that the possible hazards of enterococci included systemic infection of the human body caused by entry into blood and the transfer of drug resistance genes carried by probiotic strains, etc., and suggested that enterococci should not be used in probiotics (Reid et al., 2003).
Therefore, the issue of the safety of enterococci in probiotics, especially in terms of resistance, cannot be ignored. Limited information on the susceptibility of bacteria in combination probiotic preparations can easily lead to the misconception that there is no resistance to probiotics. Surveillance of resistance is important to assess the spread of dangerous strains. As a result, it is necessary to assess the safety and monitor the antibiotic resistance of enterococci strains when they are used commercially.
The aim of this study was to isolate enterococci from combination probiotic preparations in China, analyze the phenotype and molecular characteristics of drug resistance, and assess the risk of resistance gene transfer of enterococci in probiotics.
Materials and Methods
Origin of the samples
Enterococci were isolated from five commonly used combination probiotic preparations (Table 1) purchased by Yifeng Pharmacy Chain Co., Ltd. in Nanjing, China and they can be sold at licensed offline or online pharmacies with a doctor's prescription. At the laboratory, 0.5 g of samples was mixed with 90 mL of tryptic soy broth (TSB) incubated for 24 h at 37°C. Then, 0.1 mL of enriched broth was inoculated onto the TSB agar. After incubation at 37°C for 48h, several suspected Enterococcus colonies were randomly selected according to the appearance of the colonies and transplanted into an Enterococcus selective medium for culture. After 24 h incubation at 37°C, three Enterococcus colonies were randomly selected and transferred to TSB medium for proliferation culture. Presumptive identification of enterococci was made based on colony morphology, the pyrrolidonyl arylamidase test (Hammad et al., 2022), the catalase test, and stored in broth medium with glycerol at −80°C.
Results Of 16S rRNA Identification Of 15 Bacterial Isolate Strains
Species identification
A total of 15 isolates were obtained and identified using 16S rDNA restriction analysis. The V3–V4 hypervariable regions of the 16S rDNA gene were amplified from extracted DNA samples using two primers containing the universal sequences 27 F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492 R (5′-CTACGGCTACCTTGTTACGA-3′) (Hou et al., 2018). PCR products were sequenced on a single lane of an Illumina MiSeq platform, and the NCBI Blast program was used to compare the assembled sequence files with data from the NCBI 16S database.
Preparation of antibiotic solutions
All the antibiotics, including chloramphenicol, norfloxacin, minocycline, sulfamethoxazole, azithromycin, ampicillin sodium, amoxicillin, gentamicin, and vancomycin, were purchased from National Institutes for Food and Drug Control (Beijing, China). Each antibiotic was dissolved in an appropriate solvent with an initial concentration of 2048 μg/mL.
Antimicrobial susceptibility testing
According to the Clinical and Laboratory Standards Institute (CLSI, 2017) standard, E. faecalis ATCC29212, Escherichia coli ATCC25922, and Staphylococcus aureus ATCC29213 were used as the quality control strain, and the minimal inhibitory concentrations (MICs) of enterococci to various antibiotics were determined by broth microdilution method. Briefly, overnight bacterial cultures were diluted 1:100 in TSB and incubated at 37°C until turbidity was equivalent to McFarland standard 0.5 (108 cfu/mL). The bacterial suspension was then diluted to 1 × 105 cfu/mL, and 100 μL of the bacteria was added to each well of a 96-well plate (Corning) (Nedbalcová, 2015). Then, 100 μL antibiotic solution was inoculated in the first row of the 96-well plate and serially diluted to the 11th well by the multiple dilution method, so that the final volume of each well was 100 μL and the 12th well without drug was used as a growth control. The plates were incubated in an incubator at 37°C without stirring for 24 or 48 h, and the MIC that could inhibit the gross growth of bacteria was recorded. The results were compared directly with negative controls. The experiment was repeated three times.
According to CLSI guidelines (CLSI, 2017) and EFSA guidelines (EFSA Panel on Additives and Products or Substances used in Animal Feed [FEEDAP], 2012), strains with MIC values higher than the reference breakpoint values were considered drug-resistant strains.
Whole genome sequencing
The total DNA of the bacteria was extracted using the cetyltrimethylammonium bromide method to meet the amount of DNA required for whole genome sequencing (Healey et al., 2014). The WGS (Whole Genome Shotgun) strategy was used for the whole genome sequencing of strains in this study. The PacBioRS II sequencing platform was used for the whole gene sequencing analysis, which was completed by Shanghai Paisenuo Biotechnology Co., Ltd. The sequences of all five isolates in this study were submitted to the NCBI GenBank database. The accession numbers were retrieved as CP123615 (J-1-A), CP123609 (J-2-A), CP123611 (J-3-A), CP123613 (J-4-A), and CP123624 (J-5-A).
Antibiotic resistance gene analysis
The BLAST software was used to compare the gene coding protein sequence with the sequence in the Comprehensive Antibiotic Resistance Database (CARD) (McArthur et al., 2013) to detect the genes related to antibiotic resistance in the genome.
Virulence gene analysis
BLAST software (Stephen et al., 1990) was used to compare the gene coding protein sequence with the amino acid sequence (Set A) in VFDB (Virulence Factor Database) (Chen et al., 2016) to detect the virulence factor related genes in the genome.
Data analysis
Use SPSS 22.0 software to perform statistical analysis on the measured data and calculate the average value and standard deviation of each index. The study flowchart can be seen in Supplementary Figure S1.
Results
The identification information and results of all isolates are presented in Table 1. Fifteen strains of suspected Enterococcus species isolated from samples of five compound probiotics (J-1-A, B, C; J-2-A, B, C; J-3-A, B, C; J-4-A, B, C; J-5-A, B, C) were identified by 16S rDNA, and all isolates belonged to the Enterococcus spp. Of these, J-5-A, B, and C were identified as E. faecalis and the rest as E. faecium. All comparison results are more than 99.5% similar.
The results of antibiotic susceptibility testing on bacterial isolates are shown in Table 2. According to the established criteria, 93.3% (14/15) of isolates were resistant to norfloxacin, 93.3% (14/15) to azithromycin, 86.7% (13/15) to gentamicin, 20.0% (3/15) to chloramphenicol, and 100% (15/15) to sulfamethoxazole.
Minimal Inhibitory Concentration (μg/mL) Data And Antibiotics Resistance Profiles Of 15 Bacterial Isolates To 9 Antibiotics Based On Antibiotic Susceptibility Testing
S, susceptible; I, intermediate; R, resistant.
None of the isolates was resistant to amoxicillin, ampicillin, minocycline, and vancomycin. Among them, the glycopeptide antibiotic vancomycin had the best antibacterial effect, and its MIC value against most of the strains was ≤1 μg/mL, regardless of E. faecium or E. faecalis.
The results showed that all strains except J-1-B were resistant to norfloxacin. It is noteworthy that all E. faecium strains were not resistant to chloramphenicol, but all three E. faecalis strains were resistant to chloramphenicol with MIC values ≥64 μg/mL. As there is no breakpoint standard for gentamicin MIC of enterococci in CLSI guidelines (CLSI, 2017), EFSA guidelines (EFSA Panel on Additives and Products or Substances used in Animal Feed [FEEDAP], 2012) criteria were used to determine resistance. All strains were resistant to gentamicin except J-1-C. In addition, as neither the CLSI nor EFSA have breakpoints for the MIC of azithromycin and sulfamethoxazole against enterococci, all isolates except J-3-B were resistant to azithromycin according to the CLSI guidelines (CLSI, 2017) criteria for the reference bacteria S. aureus.
The basic genomic characteristics of the five isolates are presented in Table 3. The Guanine and Cytosine content, sequence length, protein coding sequence, number of tRNA, CRISPRs, plasmid number, and sequence type of the four strains of E. faecium J-1-A, J-2-A, J-3-A, and J-4-A were similar. In particular, J-3-A and J-4-A are almost identical in all other respects except for sequence length and protein-coding sequence, indicating a high degree of homology. Although J-2-A is E. faecium, it is different from the other three E. faecium strains in that the sequence length and Protein-coding sequences are the shortest, but the plasmid length is the longest, suggesting that the homology of J-2-A is not as good as that of the other three strains. J-5-A is E. faecalis. Compared to the other four strains of E. faecium, it has the lowest percentage of GC, the longest sequence length and protein coding sequence length, less tRNA, and no plasmids in the genome. It is suggested that J-5-A may have genetic characteristics different from E. faecium.
General Features of The Genomes of Five Isolate Strains
The CARD contains a wide range of high-quality reference data on the molecular basis of antimicrobial resistance, with an emphasis on the genes, proteins, and mutations involved (Jia et al., 2017). There was no significant difference in the number of antibiotic-related genes among the five strains. The number of antibiotic resistance genes from J-1-A to J-5-A was 29, 29, 31, 31, and 30, respectively, and 1.126%, 1.128%, 1.187%, 1.188%, and 1.057% of total protein-coding genes, respectively. The number of antibiotic target genes was 16, 16, 16, 16, and 17; the proportion of total protein-coding genes was 0.622%, 0.621%, 0.613%, 0.613%, and 0.599%, respectively (Fig. 1). In addition, none of the plasmids contained genes associated with antibiotics.
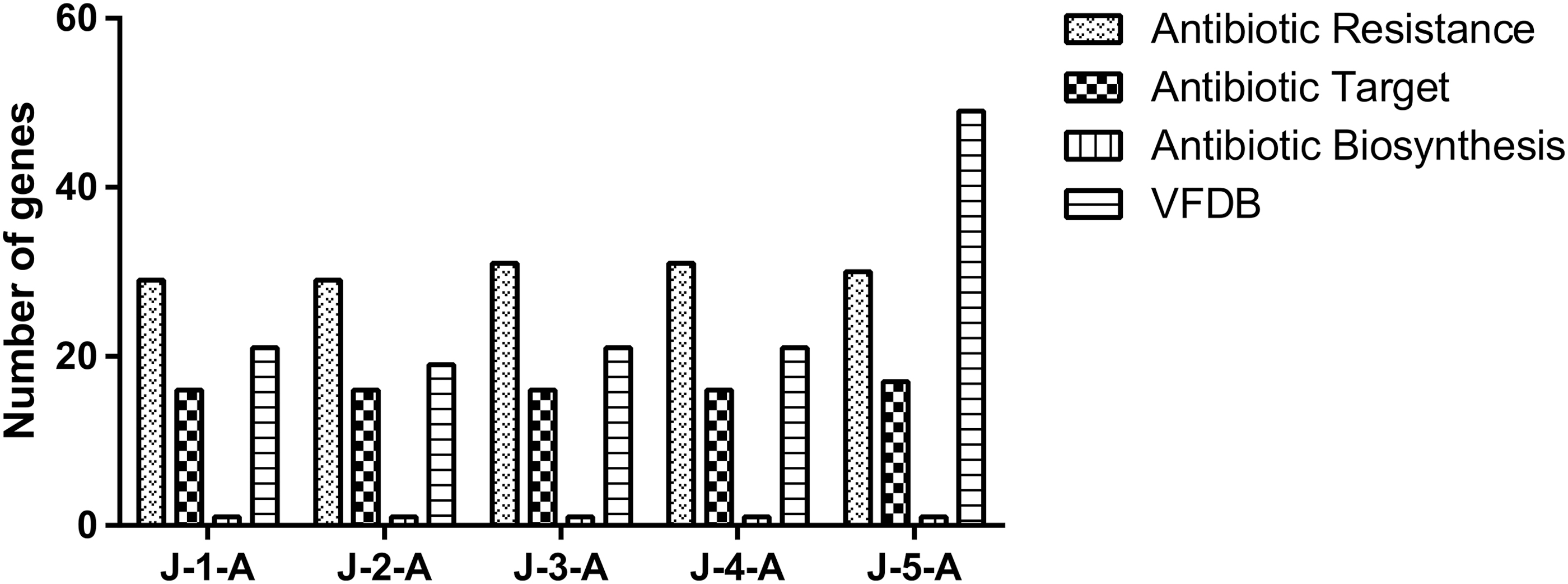
Number of antibiotic-associated genes and virulence factors.
Streptomycin resistance gene strA; lincosamide and streptogramin A resistance gene lsaA; the multidrug resistance efflux pump genes mdtG; and multidrug transporters lmrC and lmrD were all detected in J-1-A to J-5-A strains. Macrolide resistance gene msrC and polymyxin resistance gene pmrE were detected in all strains except J-2-A strains. In addition, chloramphenicol resistance genes ykkC, ykkD and the multidrug efflux pump gene emeA were also detected in J-5-A. No resistance gene was found in all plasmids.
The virulence factors for J-1-A to J-5-A were analyzed by querying the VFDB, and the number of virulence factors found in each was 21, 19, 21, 21, and 29 (Fig. 1). No virulence factor was found in all plasmids.
Gene island (GI) prediction (Fig. 2) revealed 14 GIs in J-1-A and the 9th GI was the longest one, the GIs contained 1 drug resistance gene (strA) and 3 virulence factors (wbtB, tufA, bsh); 17 GIs in J-2-A and the 6th GI was the longest one, the GIs contained 1 drug resistance gene (strA) and 4 virulence factors (acm, gndA, tufA, bsh); 15 GIs in J-3-A and the 11th GI was the longest one, the GIs contained 1 drug resistance gene (strA) and 3 virulence factors (wbtB, tufA, bsh); 15 GIs in J-4-A and the 11th GI was the longest one, the GIs contained 1 drug resistance gene (strA) and 3 virulence factors (wbtB, tufA, bsh); 13 GIs in J-5-A and the 5th GI was the longest one, the GIs contained 2 drug resistance genes (strA, pmrE) and 2 virulence factors (tufA, bsh). GI was not found in all plasmids.

GIs forecast in the genome. Outermost layer the predicted results of the gene island after the integration of the four methods and total 54 GIs were predicted. Second layer represents the results predicted by the IslandPath-DIMOB method; Third layer, the SIGH-HMM method; Fourth layer, the IslandPick method.
Discussion
Almost all strains in this study were resistant to norfloxacin, but the common plasmid-mediated quinolone resistance gene (qnr gene) and Quinolone efflux pump gene oqxA/B were not detected (Kosikowska et al., 2022). However, mutations in the parC, parE, gyrA, and gyrB genes, which determine quinolone resistance in bacteria, can also cause resistance (Aldred et al., 2014; Nathania et al., 2022). The results of this study are not certain to be related to these mutations, but a mutation in the gyrA gene has been found in quinolone-resistant enterococci isolated from yogurt (Li et al., 2015).
In addition, E. faecalis also has some inherent resistance to aminoglycoside drugs, which may be related to the change of ribosomal target sites caused by the methyl transferase encoded by aminoglycoside 6′-N-acetyltransferase encoded by the conserved gene aac(6′)-li located in the chromosome (Jang et al., 2020). This is consistent with the results of this study, where all strains were resistant to gentamicin and this gene was also found in strains J-1-A to J-4-A. At the same time, intrinsic resistance in enterococci is also associated with some ATP-binding cassette superfamily efflux pump genes. For example, lsaA gene can mediate the resistance of E. faecalis or E. faecium to macrolide-lincomycin streptavidin antibiotics (Singh et al., 2002). Furthermore, it has been suggested that enterococci may represent a major reservoir of the lsaA gene (Mišić et al., 2022). The data in this experiment also showed similar results, and the gene lsaA and another streptomycin resistance gene strA were found in all strains.
Interestingly, none of the E. faecium isolates was chloramphenicol resistant, but all three E. faecalis isolates were chloramphenicol resistant. This may be due to the two chloramphenicol resistance genes ykkC and ykkD found only in E. faecalis J-5-A, the first time such resistance genes have been identified in E. faecalis. These two genes, first identified in Bacillus subtilis, encode homologues of the small multidrug resistance family of drug efflux pumps (Dong et al., 2022). No sulfamethoxazole resistance genes were found in J-1-A to J-5-A, but all strains were extremely insensitive to sulfamethoxazole. The reason may be that enterococci have the ability to directly absorb folic acid from the environment, which leads to the inherent resistance of sulfamethoxazole because it cannot compete to inhibit the tetrahydrofolate synthesis pathway of bacteria (Christaki et al., 2020).
Notably, the emeA gene was detected only in J-5-A. The multidrug resistance efflux pump encoded by emeA belongs to the most common major facilitating family in bacteria and is a multidrug efflux transporter, which can produce resistance to a variety of antibiotics (Panthee et al., 2021). Furthermore, the emeA gene can actively remove fluoroquinolones, making E. faecalis more susceptible to developing resistance to fluoroquinolones (Esfahani et al., 2020).
In addition, in the gene sequences, we find that the number of CRISPRs in J-1-A to J-5-A is the same three, but the number of CRISPR spacers is significantly different, with J-1-A to J-4-A both containing four spacers and J-5-A containing 11 spacers. The mechanism of CRISPRs is similar to RNAi of eukaryotes, the spacer is used to identify and silence exogenous genetic material, effectively prevent the transformation of conjugative plasmids, and to some extent reduce the spread of drug resistance genes among pathogenic bacteria (Gill et al., 2005). This means that the larger the number of spacers in a bacterium, the more difficult it is to obtain resistance genes through lateral transfer.
Meanwhile, there were plasmids in J-1-A to J-4-A, but no drug resistance genes and virulence factors were found in the plasmids, indicating that these strains had little risk of spreading drug resistance through plasmids.
Studies have shown that E. faecalis tends to be more virulent than E. faecium, and its ability to acquire virulence genes is also higher compared with E. faecium (Yang et al., 2015). The present experimental results are also consistent with this view, as the number of virulence factors found in E. faecalis was much higher than that in E. faecium.
The virulence factors found in E. faecium (J-1-A to J-4-A) were almost the same in function and number, and some of them play a crucial role in the pathogenicity of Enterococcus spp. Endocarditis antigen (efaA) was detected in all strains. EfaA is a lipoprotein that exists in both E. faecalis and E. faecium. It is an adhesive antigenic component expressed on the surface of enterococci after growth in serum and is prone to provoke an inflammatory response (Low et al., 2003).
Notably, the collagen adhesins ace (E. faecalis) and acm(E. faecium) were found in J-2-A and J-5-A strains, respectively, which are widely distributed in enterococci and play important roles in bacterial colonization and pathogenesis by binding to specific proteins in the extracellular matrix through recognition and adhesion (Strateva et al., 2016).
In addition, the sprE and gelE genes were found only in J-5-A. Their expression is regulated by their upstream regulatory gene system, a two-component quorum sensing regulatory system that senses cell density and regulates virulence, participates in inflammatory processes, and is associated with the spread of enterococci around the site of infection (Kanak et al., 2023).
GIs can be involved in a variety of microbial activities, in particular as one of the important mechanisms of bacterial pathogenicity and resistance (Kim et al., 2009). The predicted GI of E. faecium was different from that of E. faecalis. The GI of strains J-1-A, J-3-A, and J-4-A was highly similar, with many identical GIs, including the largest. Furthermore, the first GI of all strains was highly similar, perhaps the fixed GI of Enterococcus spp. It is important to note that the virulence factor bsh and the streptomycin resistance gene strA are present in the GIs of all strains, and the former can help enterococci rapidly adapt to different environments. The latter may be one of the reasons that enterococci are susceptible to acquired streptomycin resistance.
Therefore, resistance and virulence factors must be identified if Enterococcus spp. is to be used as a probiotic (Yuksekdag et al., 2021). It should be noted that the results of this study do not determine whether these strains are multidrug resistant (Magiorakos et al., 2012). The presence of these resistance genes does not necessarily mean that they will be expressed, and the transfer of resistance genes also requires certain conditions. However, this safety risk in probiotics cannot be ignored.
Conclusions
Fifteen strains of Enterococcus spp. isolated from five combination probiotic preparations sold in China were tested for antibiotic resistance in this study. The results showed that almost all the isolated strains were resistant to at least one antibiotic. In a subsequent genome-wide analysis, it was found that each strain contained many resistance genes and virulence factors that were at risk of horizontal transfer. These results suggest that enterococci used as probiotics can act as reservoirs of antibiotic-resistant genes and may transfer resistance and pathogenicity to other strains, potentially posing a threat to human health. It is suggested that there are some hidden dangers in the safety of probiotic preparations and that it is necessary to further explore whether Enterococcus spp. is still suitable for use as a strain in probiotic preparations.
In view of the lack of uniform indicators and standards for the safe use of enterococci in China, we call for improved development of relevant standards and a well-established quality control inspection system, especially for probiotic preparations that can be used by infants.
Footnotes
Authors' Contributions
X.J.: Conceptualization (lead); writing-original draft (lead); formal analysis (lead); writing-review and editing (lead); Methodology (equal); Software (equal). R.C.: Methodology (equal); Investigation (equal). R.W.: Investigation (equal); Software (equal). Y.C.: Project administration (lead); Resources (equal). Z.D.: Resources (equal). Z.H.: Software (equal). X.G.: Resources (equal). L.T.: Software (equal). L.Z.: Supervision (lead); Resources (supporting).
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Scientific Research Program of Jiangsu Drug Administration (no. 202123) and Key science and technology project of Nanjing Market Supervision and Administration (no. Kj2021049).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.