
Abstract
Biofilm-associated foodborne Salmonella infections in poultry have become increasingly challenging for veterinarians, particularly in developing countries, and warrant thorough investigation. We assessed the biofilm-forming tendency of poultry isolates of Salmonella enterica, namely Salmonella Typhimurium (n = 23), Salmonella Infantis (n = 28), and Salmonella Heidelberg (n = 18), in nutrient-rich Rappaport-Vassiliadis Soya (RVS) peptone broth and nutrient-deficient diluted Tryptone Soya Broth (TSB). Seven of the tested isolates exhibited moderate biofilm formation in diluted TSB, whereas two showed such formation in RVS. In addition, the Congo red agar assay revealed curli and cellulose production in seven isolates. Fourteen specific biofilm-associated genes were analyzed identifying sdiA and seqA to be the most prevalent (100%), and glyA the least prevalent (69.5%). The prevalence of the genes bcsA and csgA was significantly lower in moderate and weak biofilm formers, respectively, as compared with nonbiofilm formers in RVS peptone broth. Furthermore, the compounds carvacrol and 2-aminobenzimidazole (2-ABI) effectively inhibited biofilm formation by Salmonella serovars in RVS peptone and TSB media, respectively. Whereas the antibiofilm activity of 2-ABI against Salmonella has not been reported previously, we determined its most effective concentration at 1.5 mM among tested antibiofilm treatments. These findings indicate that Salmonella strains prevalent in poultry farms have the potential to form biofilms, and the tested compounds should be further explored as supportive or alternative antimicrobials.
Introduction
S
The extracellular polymeric substance (EPS) matrix of biofilms typically comprises polysaccharides, extracellular DNA, proteins, and lipids, providing protection against environmental stresses (Gonzalez-Machado et al., 2018). Moreover, this matrix reduces penetration of antibacterial compounds (Idrees et al., 2021; Soto, 2014), thus enhances antimicrobial resistance from 100 to 1000-fold as compared with planktonic counterparts (Ceri et al., 1999), and the bacteria within biofilms can exchange plasmids through conjugation (Rabin et al., 2015). Biofilm formation capacity can vary even among bacterial isolates of the same serovar due to differential responses to environmental conditions (Roy et al., 2021; Wang et al., 2013).
Factors such as nutrient availability, temperature, and time play crucial roles in the extent of biofilm formation by zoonotic Salmonella (Gemba et al., 2022; Wang et al., 2013). In addition, Salmonella can form biofilms on inert materials used in kitchen and poultry processing units such as stainless steel, polystyrene, and acrylic surfaces (Ban et al., 2014). The poultry industry is a major sector in Pakistan, fulfilling 35.4% of the country's total meat requirements and exports (Pakistan Economic Survey 2020–21). Zoonotic serovars such as Salmonella Heidelberg, Salmonella Infantis, Salmonella Enteritidis, and Salmonella Typhimurium are pathogenic to both poultry and humans (Stevens et al., 2009).
The production of cellulose and curli fibers by Salmonella results in a tightly packed matrix enhancing bacterial protection (Cookson et al., 2002; Solano et al., 2002). Strong biofilms formed by Salmonella under alkaline pH, room temperature, and limited nutrient availability exhibit high resistance to the chlorine treatments commonly used to sanitize food contact surfaces in the food processing units (Yang et al., 2016).
Several genes involved in different stages of biofilm formation have been reported in Salmonella. The genes sdiA (Smith et al., 2011) and LuxS (Sun et al., 2020) are associated with quorum sensing detecting chemical signals from neighboring bacteria. Some protein synthesizing genes such as csgA, csgB (Brombacher et al., 2006), and csgD (Prigent-Combaret et al., 2001), and exopolysaccharides coding genes such as wcaA (Crawford et al., 2008) and bcsA (Castiblanco and Sundin, 2018) contribute to bacterial adhesion. The genes such as fliC (Ling et al., 2021), seqA, fimH (Hancox et al., 1997), and bapA also help bacteria to adhere to the host surface (Latasa et al., 2005).
The pfs is reported to promote the autolysis of bacteria leading to the release of extracellular DNA in the EPS matrix, thereby assisting in bacterial dispersion (Bao et al., 2015). It is crucial to explore antibiofilm compounds that can target various stages of the bacterial biofilm cycle. Such compounds can inhibit bacterial cell attachment to solid supports (Alasil et al., 2014), prevent the formation and maturation of attached bacterial microcolonies (Millot et al., 2017), interfere with the quorum sensing of bacterial cells (Singh et al., 2017), or disperse the preformed biofilms to avoid the persistence of biofilm structures that complicates infections (Nguyen et al., 2019).
Carvacrol, a natural substance found in thyme and 2-aminobenzimidazole (2-ABI), a synthetically produced substance, has been reported as quorum sensing blockers inhibiting biofilm formation (Burt et al., 2014; Rogers et al., 2009; Tan et al., 2019). However, the antibiofilm activity of 2-ABI on Salmonella isolates has not been previously determined. This study was designed to characterize the biofilms formed by Salmonella enterica isolates from local poultry farms in terms of nutrient availability, determine the prevalence of biofilm-related genes, and to evaluate the antibiofilm activity of a natural (carvacrol) and a synthetic (2-ABI) compound.
Materials and Methods
Bacterial isolates
A total of 69 S. enterica isolates belonging to three foodborne zoonotic serovars: Salmonella Typhimurium (n = 23), Salmonella Infantis (n = 28), and Salmonella Heidelberg (n = 18), were taken from the institutional glycerol (30%) stock cultures stored at −20°C. As no human or animal was involved as research subject in the study, thus the Institutional Review Board approval was waived. The stocks were revived in sterile Tryptone Soya Broth (TSB; CM0129; Oxoid, United Kingdom) tubes at 37°C for 24 h and isolated on MacConkey agar (CM0007; Oxoid) and Salmonella-Shigella (SS) agar (CM0099; Oxoid) plates. The colony morphology was observed and well-isolated colonies were stabbed and streaked on Triple Sugar Iron (TSI) agar (CM0277; Oxoid) slants (Gunasegaran et al., 2011).
The molecular identification was done by polymerase chain reaction (PCR), targeting Salmonella genus-specific invA gene (Moussa et al., 2010), serovar Typhimurium-specific stm gene (Liora et al., 2013), serovar Infantis-specific fljB gene (Ghoddusi et al., 2015), and serovar Heidelberg-specific ACF gene (Park and Ricke, 2015). Further molecular confirmation of the representative isolates of each serovar was done by targeting 16S rRNA gene (Chen et al., 2015). The 16S rRNA gene amplicons were purified using Thermo Scientific, GeneJET PCR Purification Kit, and sent for commercial nucleotide sequencing (Macrogen, Republic of Korea). The nucleotide sequences were compared with other available sequences in the NCBI database, using blastn and were submitted to NCBI GenBank.
Biofilm formation assay
Each of the Salmonella isolates was inoculated in TSB and incubated at 37°C with 180 rpm for 18 h. The optical density (OD) of the culture was measured at 600 nm and adjusted to 1.0 using sterile TSB. The biofilm-forming ability was checked in two media: nutrient-enriched Rappaport-Vassiliadis Soya (RVS) (Nawaz et al., 2020) and nutrient-deficient TSB (diluted 1/20 in water) (Stepanović et al., 2004). The bacterial culture was diluted in RVS and in diluted TSB media as 1:100 and pipetted in 96-well flat-bottomed polystyrene plates (Cat No. 31396; SPL Life Sciences) as 200 μL per well (triplicate wells for each isolate).
The sterile media were taken as negative controls and a biofilm-forming Pseudomonas aeruginosa isolate was used as a positive control. The plates were incubated at 37°C for 48 h for biofilm formation. The contents of each plate were discarded and the plate was washed once with 0.9% (w/v) NaCl. The biofilm biomass was stained with 200 μL of 0.1% (w/v) crystal violet per well and incubated at room temperature for 15 min (O'Toole, 2011). The contents were discarded and the plate was again washed with 0.9% (w/v) NaCl. The bound crystal violet dye was dissolved by adding 200 μL per well of 30% (v/v) glacial acetic acid (Coffey and Anderson, 2014) and incubated at room temperature for 15 min.
The plate contents were carefully shifted to another blank 96-well plate and OD of the plate was measured at 630 nm (Ghellai et al., 2014) using plate reader (Dia 710 microplate reader; Diamate Bio Technologies Ltd., United Kingdom). The Salmonella isolates were categorized into different groups of biofilm formation according to the previously reported classification (Stepanović et al., 2000). The experiments were performed twice at two independent occasions.
Congo red agar assay
Salmonella isolates were also characterized by Congo red agar (CRA) assay that detects the production of cellulose and curli protein (Reichhardt et al., 2016). The agar was prepared as 37 g/L Brain Heart Infusion broth (CM1135; Oxoid), 50 g/L sucrose (S391; Phyto Technology Laboratories), 10 g/L Bacteriological Agar No. 1 (LP0011; Oxoid), and 0.8 g/L Congo red dye (Winlab, United Kingdom). For the assay, 5 μL of the overnight culture was carefully inoculated on the solidified agar plates in the form of spots (Kaiser et al., 2013).
Four isolates were inoculated on each plate and subjected to 48 h incubation and the colony morphology was observed. The isolates exhibiting red growth with moist surface were classified as curli-negative, whereas those with whole or partially blackened colonies were classified as curli-positive isolates (Kaiser et al., 2013; Mathur et al., 2006). The experiment was performed twice.
Detection of biofilm-related genes
Fourteen biofilm-associated genes mostly reported in Salmonella were targeted by PCR including sdiA, glyA, pfs, csgB, fliC, wcaA, dam, seqA, bapA, fimH, bcsA, csgD, csgA, and LuxS (Table 1). For each PCR, a 25 μL mixture was prepared containing 2.5 μL of 10 × PCR buffer, 1.5 μL of 25 mM MgCl2, 1 μL of 10 pmol/μL of each of the forward and reverse primers, 0.2 μL of 5 U/μL of Taq polymerase, 12.8 μL of 10 mM Tris-HCl, and 5 μL of template DNA.
Primer Sequences for Biofilm-Associated Genes with Annealing Conditions
The PCR amplification conditions were set as initial denaturation at 94°C for 5 min, 25 cycles of denaturation at 94°C for 1 min, primer annealing (specific for each primer), and extension at 72°C for 1 min, followed by a final extension step at 72°C for 7 min. The primer sequences along with their annealing conditions and amplicon sizes are given in Table 1.
Evaluation of antibiofilm compounds
Two antibiofilm compounds carvacrol (CAS No. 499-75-2) and 2-ABI (CAS No. 934-32-7) supplied by Sigma-Aldrich were evaluated for their biofilm inhibition activity on Salmonella isolates. The stock solutions of the compounds were prepared in absolute ethanol. For carvacrol, 100 mM stock solution was prepared (Burt et al., 2014), whereas for 2-ABI, stock solution was prepared as 75 mM solution. For working solutions of both compounds, the stock solutions were diluted in the respective medium to make 1 and 2 mM carvacrol and 0.4 and 1.5 mM for 2-ABI (Amer et al., 2021; Tan et al., 2019).
The biofilm inhibition effect of the compounds was tested on eight Salmonella isolates that exhibited moderate biofilm formation in RVS, diluted TSB or both media. The OD600nm of the overnight culture of the isolates was adjusted to 1.0. The turbid broth was diluted as 1:100 in RVS or diluted TSB or both as required. For each isolate, upper 3 wells of a 96-well plate column were filled with 200 μL of diluted bacterial culture only (bacterial control), whereas the lower 3 wells contained the solvent along with the bacterial culture (solvent control).
For the antibiofilm assay, 100 μL of the diluted bacterial culture and 100 μL of working solutions for each of the 2 antibiofilm compounds were added (6 wells for each tested concentration). The plates were incubated at 37°C for 48 h and the biofilm detection assay was performed with 0.1% (w/v) crystal violet staining as described earlier. The biofilm inhibition percentage was calculated as (1-OD630/OD630-SC) 100 (Siddique et al., 2020).
Where OD630 is the OD of the bacterial biofilm formed when treated with the antibiofilm compound and OD630-SC is the OD of the biofilm formed in the presence of solvent without any antibiofilm agent.
Statistical analysis
The data were statistically analyzed using GraphPad Prism8 and SPSS software. Any significant effect of medium and serovars on biofilm formation was tested by independent t-test. The most effective antibiofilm treatment against Salmonella was determined by one-way analysis of variance followed by an Least Significant Difference test. The chi-square test was used to identify any significant association between the presence of biofilm-associated genes and biofilm formation. All the tests were performed at a 5% level of significance.
Results and Discussion
Identification of Salmonella isolates
All 69 isolates taken from glycerol stocks were successfully revived and detected as S. enterica based on colony morphology, that is, colorless colonies on MacConkey agar and colorless colonies with or without black center on SS agar plates. Typical reactions were observed on TSI agar slants: acidic/yellow tube bottom and alkaline/pink slant portion with the blackening of the medium in the center due to the production of H2S gas. Molecular identification of the Salmonella isolates was confirmed using PCR that successfully amplified a 284 bp fragment of the invA gene.
Furthermore, serovar-level PCR confirmed 23 isolates as Salmonella Typhimurium, 28 isolates as Salmonella Infantis, and 18 isolates as Salmonella Heidelberg. Phylogenetic analysis of the representative isolates from each serovar provided additional confirmation of their identity and the corresponding sequences were submitted to GenBank with the following accession numbers: MZ683222 (Salmonella Typhimurium), MZ683260 (Salmonella Infantis), and MZ683219 (Salmonella Heidelberg).
Biofilm formation varied with Salmonella serovars and strains
This study presents the assessment of biofilm formation by local poultry isolates of Salmonella based on the use of crystal violet assay for its quantification. The assay is considered the gold standard for biofilm detection, being the easiest, most cost-effective, and simplest approach (Haney et al., 2018). In RVS medium, 31 isolates (44.9%) were classified as nonbiofilm producers, 36 (52.2%) as weak biofilm producers, and 2 (2.9%) as moderate biofilm producers. In diluted TSB, 10 isolates (14.4%) were classified as nonbiofilm producers, 52 (75.4%) as weak biofilm producers, and 7 (10.2%) as moderate biofilm producers.
The diluted TSB medium was found significantly more conducive to biofilm production than RVS medium for Salmonella Typhimurium and Salmonella Infantis, whereas for Salmonella Heidelberg, the influence of medium on biofilm formation was not significantly different as estimated by t-test (p > 0.05) (Fig. 1).
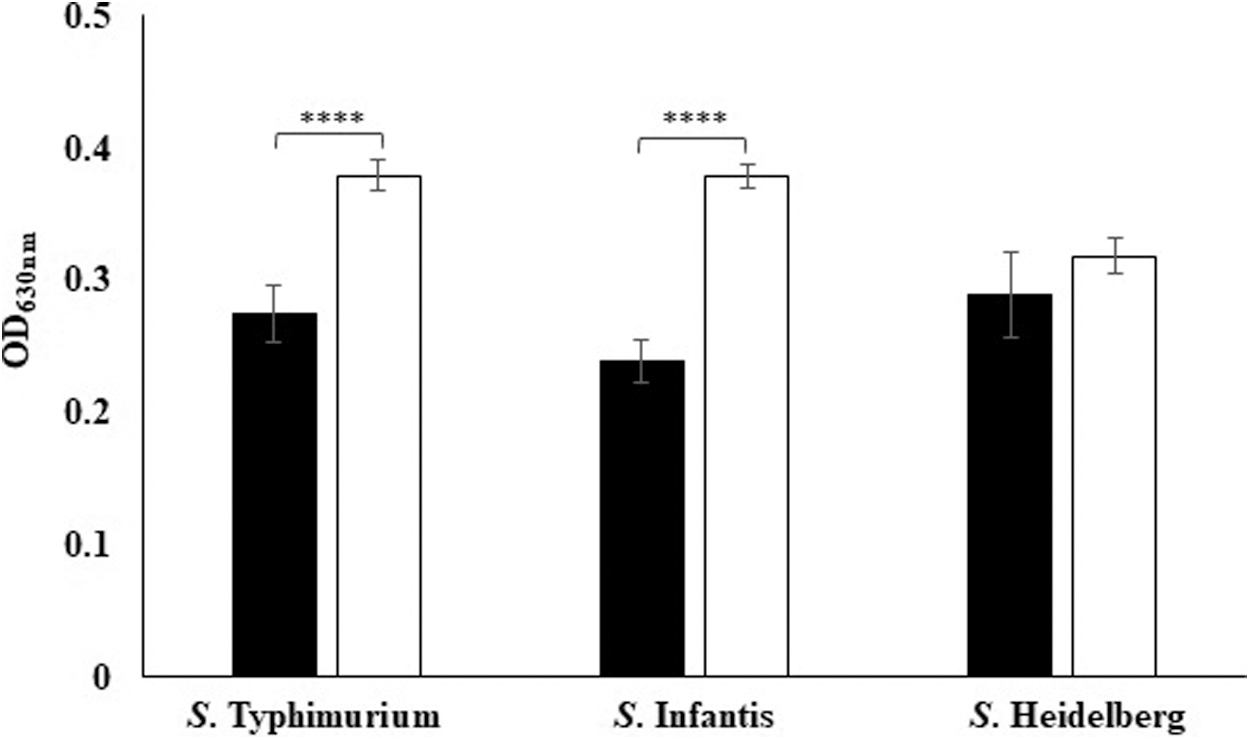
Salmonella biofilms in diluted TSB and RVS media. OD is the average OD of Salmonella isolates that is indicative of biofilm formation. The error bars represent standard error and **** indicates p < 0.0001. ■, RVS; □, diluted TSB. OD, optical density; RVS, Rappaport-Vassiliadis Soya; TSB, Tryptone Soya Broth.
Overall, Salmonella demonstrated better biofilm formation in diluted TSB than RVS medium. The results were in accordance with a previous study indicating that stress induced by limited nutrients triggers Salmonella to form biofilms (Shatila et al., 2021). Another study showed that better biofilm formation by Salmonella in diluted TSB was attributed to enhanced production of curli and cellulose, resulting in a thicker biofilm matrix (Castelijn et al., 2012). However, some studies have reported better biofilm formation by Salmonella when sufficient nutrients were available (Chandra et al., 2017).
Four Salmonella Typhimurium, three Salmonella Heidelberg, and none of the Salmonella Infantis isolates exhibited moderate biofilms in diluted TSB. In RVS medium, one Salmonella Typhimurium isolate and one Salmonella Infantis isolate formed moderate biofilms, whereas all Salmonella Heidelberg isolates showed weak biofilm formation. The biofilm-forming ability was not significantly affected by different serovars of Salmonella (p > 0.05) (Fig. 2). An earlier study also showed that serovar is not a determining factor for biofilm formation (Piras et al., 2015).

Classification of Salmonella serovars with respect to biofilm formation. Biofilm formation in , Salmonella Heidelberg.
The biofilm formation detection assay was further complemented with CRA assay whereby the production of curli protein and cellulose was detected. The CRA assay categorized 62 isolates as curli negative and seven as curli positive. The curli-positive isolates were significantly different from the biofilm formers as detected by microtiter plate assay (Table 2), making the CRA assay less reliable and less sensitive as consistent with earlier studies (Akbar et al., 2022; Hassan et al., 2011; Kord et al., 2018).
Number of Isolates Showing Curli Production and Biofilm Formation
RVS, Rappaport-Vassiliadis Soya; TSB, Tryptone Soya Broth.
Prevalence of biofilm-associated genes
All 14 targeted genes were detected in different Salmonella isolates. The prevalence of biofilm-associated genes was as follows: 100% for sdiA and seqA, 98.5% for pfs and dam, 95.6% for csgB and fliC, 94.2% for LuxS, 91.3% for csgA, 88.4% for wcaA, 85.5% for csgD, 79.7% for bapA, fimH, and bcsA, and 69.5% for glyA. Out of the 66 fliC-positive isolates, 1488 bp fragment was obtained for 48 isolates, whereas for 18 isolates, the fragment size was ∼1300 bp for which the nucleotide sequencing of both amplicons can elucidate the reason. The amplified PCR product of 1 representative isolate for each of the 14 biofilm-associated genes is shown in Figure 3a and b.
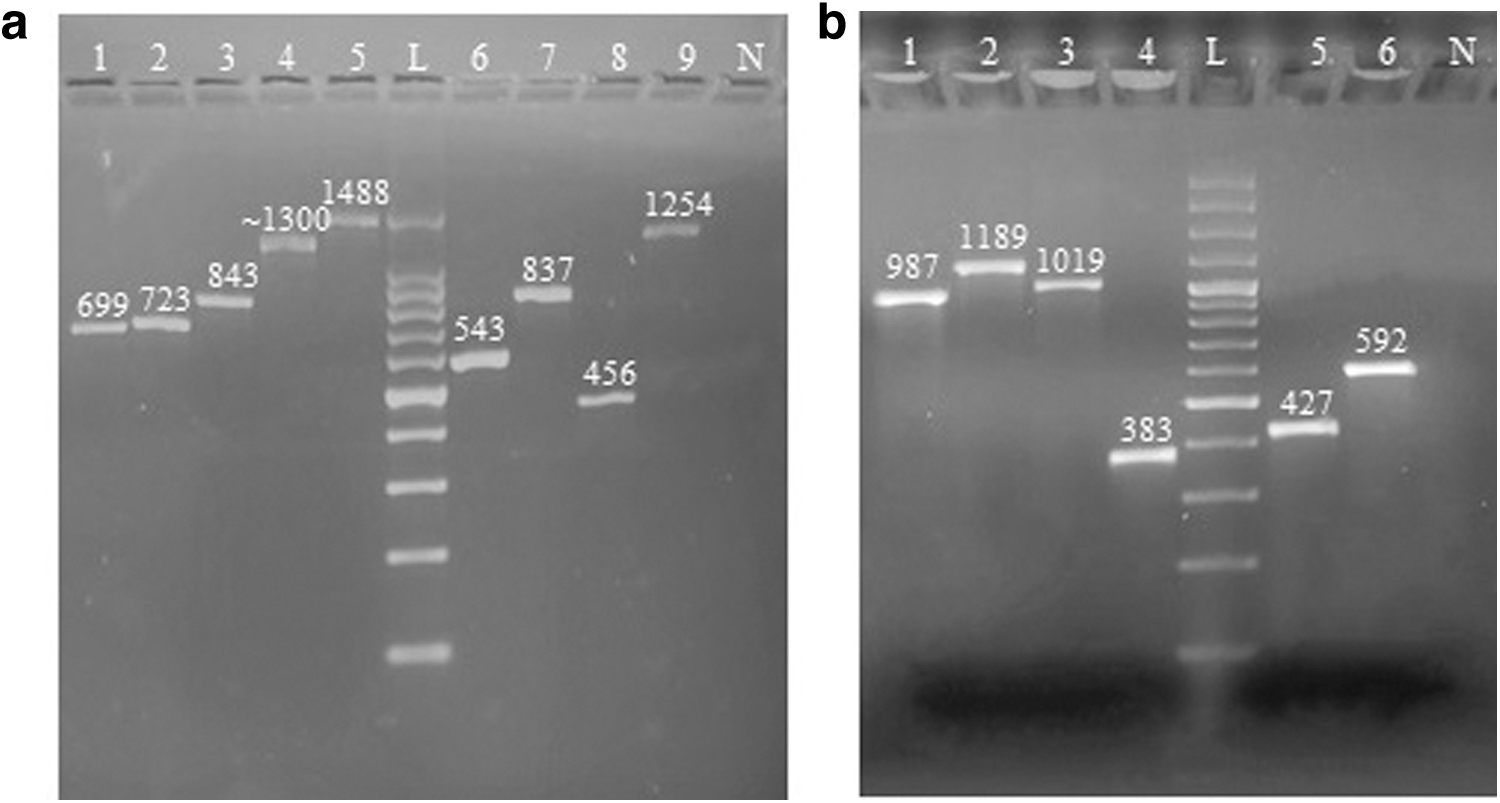
Genotypic detection of biofilm formation in Salmonella isolates.
In a previous study (Yin et al., 2018), fliC (1488 bp) and wcaA genes were reported in 100% of the biofilm-forming Salmonella isolates, whereas bcsA was reported as least prevalent at 2.6%. No significant difference (p > 0.05) was found in prevalence of detected genes among various serovars of Salmonella. However, statistical analysis showed that bcsA and csgA genes were significantly more prevalent in nonbiofilm formers in RVS medium, whereas bcsA and csgD were more common in nonbiofilm formers in diluted TSB medium.
A previous study (Jamalli et al., 2016) on biofilm formation by Escherichia coli reported that bcsA was significantly more common in biofilm formers than in nonbiofilm producers. Another study (Jahromy and Karizi, 2019) reported the presence of csgA and csgD genes as nonassociated with the biofilm-forming ability of uropathogenic E. coli. Although the targeted genes (csgA, csgD, and bcsA) were more prevalent in some isolates, they still exhibited no or weak biofilm formation, most probably due to low-level expression under laboratory-based nutrient and incubation conditions (Rédei, 2008). In contrast, the genes were found to be less prevalent in the isolates showing moderate biofilm formation, indicating the involvement of some different regulatory pathway in biofilm formation.
Antibiofilm activity of carvacrol and 2-ABI
Antibiofilm assay with carvacrol and 2-ABI showed a dose-dependent inhibition of biofilm formation. The solvent used, absolute ethanol (Burt et al., 2014; Tan et al., 2019), alone did not show noticeable biofilm inhibition effect. The biofilm inhibition was significantly different (p < 0.05) between two concentrations of carvacrol with an average percentage inhibition of 23.3% ± 1.05% at 1 mM carvacrol and 34.3% ± 1.1% at 2 mM carvacrol. Similarly for 2-ABI, significantly higher biofilm inhibition was observed at 1.5 mM (40.9% ± 2.04%) as compared with 0.4 mM (29.9% ± 2.03%).
Among the four treatments, 1.5 mM of 2-ABI was found to be more effective (p < 0.05) than other treatments against Salmonella biofilms (Fig. 4a). This concentration has previously been reported to cause 90% and 99.9% biofilm inhibition in Staphylococcus aureus and Candida albicans isolates, respectively (Tan et al., 2019). A previous study (Burt et al., 2014) reported that carvacrol inhibited 100% of biofilm formation by Salmonella Typhimurium at 1.5 and 2 mM concentrations. Another study reported that carvacrol inhibited 99% of biofilm formation by C. albicans and Streptococcus mutans when incorporated with denture soft liner material (Baygar et al., 2018).

Biofilm inhibition effect of antibiofilm compounds on Salmonella. , Salmonella Infantis (n = 1); □, Salmonella Heidelberg (n = 3);
The biofilm inhibition by carvacrol in this study is comparatively low, possibly due to multiple reasons including variations in the growth medium, biofilm-forming substrates, and the molecular mechanisms involved in the studied isolates. For Salmonella Typhimurium and Salmonella Heidelberg, 1.5 mM 2-ABI inhibited biofilm formation by 47.3% ± 2.9% and 31.3% ± 4.4%, respectively, whereas all other tested treatments had lower inhibitory effects.
For Salmonella Infantis, the highest biofilm inhibition (43.0% ± 1.3%) was observed with the 2 mM carvacrol treatment (Fig. 4b). Regarding the influence of nutrients on biofilm inhibition by different treatments, the RVS medium allowed maximum biofilm inhibition (33% ± 2.2%) by carvacrol at both concentrations, whereas in diluted TSB, the highest biofilm inhibition (45.2% ± 2.3%) was observed with 1.5 mM of 2-ABI (Fig. 4c). Both compounds, carvacrol and 2-ABI, showed antibiofilm activity on the polystyrene plates showing their antibiofilm potential, particularly on surfaces that are prone to biofilm formation, including feeding and water supply channels and utensils.
The antibiofilm effect of the tested compounds on or within any living body can further be evaluated. The assay could not differentiate between viable and dead cells within the bacterial biofilm, rather it only detected the variation in the extent of formed biofilms, so it is yet to be studied that whether the tested compounds have only antibiofilm activity or both antibiofilm and antibacterial activities. Conducting the expression analysis of biofilm-associated genes can help to identify the genes with a considerable role in biofilm formation by Salmonella. Moreover, it is imperative to test more small molecules on preformed biofilms to assess their potential for biofilm eradication and in combination with antimicrobials to kill bacterial cells released from biofilms before they can contaminate other surfaces.
Conclusion
Foodborne Salmonella isolates possess a notable ability to form biofilms that renders the associated infections challenging to treat, and leads to substantial economic losses to many industries, including poultry farming. A synthetic compound, 2-ABI, has shown promising antibiofilm activity against Salmonella biofilms. The study provides valuable insights for complementing the antimicrobial therapy with antibiofilm compounds, to inhibit bacterial biofilms. Detection of biofilm-associated genes was carried out to screen the local isolates. The role of the detected biofilm-associated genes in biofilm formation can be studied by comparing gene expression profiles of biofilm-forming and nonbiofilm-forming isolates.
The study can be further extended for the comparative gene expression analysis of biofilm-associated genes under the effect of antibiofilm compounds that can determine the roles of various genes and their encoded proteins in bacterial biofilm formation that can ultimately contribute to designing different strategies for biofilm control.
Footnotes
Authors' Contributions
F.N. performed the biofilm experiments and wrote the initial draft; Ab.A. and Y.S. designed primers and contributed to molecular detection of biofilm-associated genes; M.M.K. and P.S. contributed to phenotypic detection of biofilm and provision of antibiofilm compounds; W.R. contributed to designing and execution of antibiofilm experiments; whereas A.A. supervised the overall experiments, conceptualization, and finalized the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
The research study was partially funded by Higher Education Commission (HEC), Islamabad, Pakistan, under National Research Program for Universities project number NRPU-6111 and the Federal Ministry of Education and Research, Germany (BMBF InnoProfile-Transfer 03IPT611X and BMBF project 03PSZZF1A).