
Abstract
This study was conducted in Bejaia, Algeria, to determine the presence of Salmonella in fresh watermelon (n = 105), soil (n = 23), and irrigation water samples (n = 17) collected from two different farms. After isolation, antimicrobial susceptibility testing, serotype determination, multilocus sequence typing, antimicrobial resistance genes detection, and whole genome sequencing were performed. Twenty watermelon samples (19%) were contaminated with Salmonella, but none were found in the soil or irrigation water. Among the 20 Salmonella isolates, 2 serovars were identified (Salmonella Liverpool and Salmonella Anatum), belonging to sequence types ST1959 and ST64, respectively. Ten Salmonella isolates showed significant resistance to nalidixic acid, ofloxacin, and ciprofloxacin but were susceptible to all other antibiotics. The coexistence of point mutations (parC:p.T57S) in Quinolone Resistance-Determining Regions and the qnrB19 gene may contribute to quinolone resistance. The study identified 164 virulence genes in the Salmonella isolates. Our study found Salmonella in fresh watermelon during the preharvest season in Bejaia, Algeria. Our study indicates a relatively high prevalence of Salmonella on watermelon samples before harvest. Although we cannot directly compare our results with previous studies, it is crucial to recognize that the absence of comprehensive comparative data underscores the need for further research and surveillance.
Introduction
Vegetables and fruits are essential to a healthy and balanced diet (Slavin and Lloyd, 2012). However, because these products are primarily consumed raw and often undergo minimal processing that could otherwise reduce their microbiological contamination, they can become sources of infection and transmit pathogens that cause human food poisoning (Kowalska, 2023). Health agencies such as the World Health Organization, European Food Safety Authority, Food and Agriculture Organization, and French Agency for Food Safety (AFSSA) strongly encourage the consumption of these foods to protect against various illnesses, including cancer and cardiovascular diseases (Abaza, 2017).
Consuming fresh vegetables, root vegetables, sprouts, and fruits in their raw form can expose humans to foodborne pathogens, including antibiotic-resistant bacteria. In recent years, exposure to antimicrobial-resistant pathogens through the food chain has become increasingly associated with foodborne disease outbreaks (Pérez-Rodríguez and Mercanoglu Taban, 2019; Rahman et al., 2021). Disease outbreaks in Germany, the United States, Canada, Australia, and Finland have been linked to multidrug-resistant Escherichia coli and Salmonella spp. recovered from vegetables (Rahman et al., 2021).
Watermelons have been linked to numerous foodborne illnesses in the past decade. Unlike thermally processed foods, watermelons are consumed fresh and are, therefore, at greater risk of bacterial contamination (Miao et al., 2018). Inadequate washing during processing can lead to foodborne illnesses such as salmonellosis, as these fruits naturally carry contaminants from soil and water (Taban et al., 2013).
Fresh products are susceptible to bacterial pathogen contamination at several production and supply chain stages, including farming, transportation, and handling. Direct contact with fecal waste, including biosolids or animal manure as fertilizer and wastewater irrigation, can result in contamination during farming. Similarly, contamination can occur during the transportation and handling of produce (Chee-Sanford et al., 2009; Jung et al., 2014).
Watermelon can get contaminated with Salmonella through exposure to contaminated soil, irrigation water, air, or animal feces. The bacteria can survive on the surface of the watermelon rind and spread to other parts of the fruit during handling. Consuming contaminated watermelon can lead to diarrhea, fever, and abdominal cramps, especially severe in vulnerable groups (Iwu and Okoh, 2019; Parnell et al., 2005).
Salmonella, a well-known pathogen belonging to the Enterobacteriaceae family, is a leading cause of foodborne illnesses worldwide (Reddy et al., 2016). Its impact on public health and food safety cannot be overstated. Salmonella infections can cause various digestive symptoms, whereas severe cases can lead to hospitalization and fatalities, particularly among vulnerable populations such as young children, older people, and people with weakened immune systems (Iwu and Okoh, 2019; Parnell et al., 2005).
Although antimicrobial treatment is typically not required for human nontyphoidal salmonellosis, it becomes essential in systemic infection (Gales et al., 2002). Moreover, Salmonella contamination of food products can result in substantial economic losses and erode consumer confidence in food safety (Reddy et al., 2016). Therefore, comprehensive knowledge of Salmonella's prevalence, sources, and antimicrobial resistance is vital for implementing effective preventive strategies, making monitoring programs essential (Miranda et al., 2009).
There is a lack of research on Salmonella's prevalence in Algeria's fruits and vegetables. One study on vegetables reported no presence of Salmonella (Mesbah Zekar et al., 2017). Our study aims to detect and serotype Salmonella in watermelon samples collected from two farms in Bejaia, Algeria, and characterize the antibiotic resistance and genome structure of the Salmonella enterica isolates obtained from the samples.
Materials and Methods
Study area
The study was conducted in two watermelon production farms, Sameoun and Amizour, with surface areas of ∼12 and 6 hectares, respectively. These selected farms represent the significant contributors to watermelon production in the area and were located about 18 km apart. During preharvesting, random samples of healthy Citrullus lanatus watermelon fruits were collected from the farms. The watermelon plants were spaced 10 m apart in the fields, ensuring adequate airflow and minimizing the risk of cross-contamination between plants.
Collection of samples
In this study conducted from April to June 2022, a total of 145 samples were collected, including watermelons (n = 105), soil (n = 23), and irrigation water (n = 17). The sampling process of watermelon was executed using sterile swabs to collect samples from the watermelon surface, including the stem, bottom, and sides of the watermelon fruits. To identify potential sources of contamination, soil samples were also collected using a soil auger, whereas irrigation water samples were collected using a sterile container.
Samples were gathered from different areas across the farm premises and transported in a cooler equipped with ice packs, ensuring a temperature range of 2–8°C throughout transit. Upon arrival at the laboratory, the samples underwent processing for microbiological analysis to identify the potential presence of Salmonella.
The soil samples were obtained from watermelon fields that had been previously amended with animal-based soil amendments. Each soil sample represented a pooling of 10 individual samples harvested from different areas within the farms. This pooling approach was employed to comprehensively represent soil microbial populations and potential variations across the farms (Armalytė et al., 2019; Osborne et al., 2011; Walder et al., 2022). For the irrigation water samples, we aimed to capture a comprehensive representation of the various sources of water used by farmers for irrigating watermelon plants. These sources included well water (n = 10) and surface water (n = 7). A volume of 250 mL was collected for analysis.
Microbiological analysis
Salmonella spp. was isolated from swab samples following the ISO 6579:2002/Amd 1:2007 guidelines. The surface swab was briefly taken and then transferred into ∼10 mL of buffered peptone water (BPW) (Fluka, St Louis). According to established protocols, 25 g of soil and 25 mL of water were added to 225 mL of BPW for soil and water samples. All samples were then incubated at 37°C for 18–24 h. The sample was then subcultured into selective enrichment in Rappaport-Vassiliadis broth (Fluka) and incubated at 41.5°C for 24 h.
After incubation, aliquots were streaked onto selective agar media such as xylose lysine deoxycholate (XLD) agar (Fluka) and incubated at 37°C for 24 h. Presumptive colonies on XLD media were then transferred to MacConkey agar plates and incubated at 37°C for 24 h to obtain pure cultures. Species identification was accomplished by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (Biotyper MBT Compass V4.2.100; BRUKER).
Antimicrobial susceptibility testing
The disk diffusion method on Mueller–Hinton agar (Bio-Rad, France) was used to perform antimicrobial susceptibility testing on all confirmed strains. The results were interpreted based on the Clinical and Laboratory Standards Institute recommendations guidelines (CLSI, 2019). A total of 20 antibiotics (Bio-Rad) were tested, including ampicillin (10 μg), amoxicillin/clavulanic acid (20 + 10 μg), ticarcillin (75 μg), piperacillin/tazobactam (30 + 6 μg), cefoxitin (30 μg), cefotaxime (30 μg), ceftazidime (30 μg), aztreonam (30 μg), imipenem (10 μg), ertapenem (30 μg), streptomycin (10 μg), gentamicin (10 μg), tobramycin (10 μg), amikacin (30 μg), nalidixic acid (30 μg), ofloxacin (5 μg), sulfonamides (250 μg), tetracycline (30 μg), trimethoprim (5 μg), and trimethoprim/sulfamethoxazole (1.25 + 23.7 μg).
In addition, the minimum inhibitory concentrations (MICs) of ciprofloxacin were determined using E-test strips. In our study, we used the E-test method as a convenient and reliable alternative to the traditional agar dilution method for determining MIC values, which has been shown to provide comparable results to the agar dilution method (Jorgensen and Ferraro, 2009).
Whole genome sequencing and in silico analysis
The Bacterial DNA Extraction kit for SaMag automaton (Sacace Biotechnologies) was used to extract DNA from all identified strains of Salmonella. Libraries were carried out with the Ion Xpress Plus Fragment Library kit on the AB Library Builder system (ThermoFisher Scientific, Waltham, MA). Whole genome sequencing was then performed with Ion Torrent technology on an S5 sequencer following the manufacturer's instructions.
The quality of the fastq raw reads was evaluated using FastQC v0.11.9 and MultiQC v1.13. After alignment against the reference genome (GenBank NZ_CP025554.1) using bwa v0.7.17-r1188 and Samtools v1.15.1 algorithms with default parameters, a minimum mean depth 30x was established. The reads were then assembled into contigs using SPADES v3.13.0 software, and the resulting reads are available under the Sequence Read Archive accession number PRJNA950207.
Serovar prediction and multilocus sequence typing were performed with SeqSero (Zhang et al., 2015) and pubMLST (Jolley et al., 2018), respectively. Core genomic single nucleotide polymorphisms (SNPs) were identified using Snippy v4.4.5 (https://github.com/tseemann/snippy) against the reference strain Salmonella Enteritidis ATCC BAA-708 strain (NZ_CP025554.1). Fasttree v2.1.10 and Figtree v1.4.4 software were used to construct a phylogenetic tree based on SNP comparison.
In addition, in silico analysis was performed on the assembled genome sequences to detect the presence of acquired antibiotic resistance genes (ARGs), plasmid replicon types, and virulence genes. The ResFinder 4.1 database (https://cge.cbs.dtu.dk/services/ResFinder/) with a 90% identity and 80% coverage setting was used to detect ARGs, whereas plasmid replicon types were identified using the Plasmidfinder database (https://cge.food.dtu.dk/services/PlasmidFinder/) with the same settings. To screen the presence of virulence genes in the Salmonella genome sequences, the virulence factor database (VFDB) was used with the default setting (www.mgc.ac.cn/VFs/main.htm) (Liu et al., 2019).
Results
Prevalence of Salmonella spp.
During the study period, Salmonella was detected in 20 out of 105 watermelon samples analyzed, representing a prevalence of 19%, and was absent in soil and irrigation water samples (0%). Most positive samples originated from Semaoun farm (85% or 17 out of 20), whereas the remaining positive samples were from Amizour farm (15% or 3 out of 20).
Serotyping and phylogeny of Salmonella isolates
Isolated Salmonella strains were subjected to serotyping, identifying two serovars: Salmonella Liverpool (10/20; 50%) and Salmonella Anatum (10/20; 50%). Further analysis using multilocus sequence typing (MLST) revealed that the Salmonella Liverpool strains belonged to ST1959, whereas the Salmonella Anatum strains belonged to ST64 (Table 1).
Genomic Characteristics and Resistance Profile of Salmonella enterica subsp. enterica Serotypes Found in Watermelon Samples
CIP, ciprofloxacin; MIC, minimum inhibitory concentration; NAL, nalidixic acid; OFL, ofloxacin; QRDR, Quinolone Resistance-Determining Region; R, resistant; S, susceptible.
The phylogenetic relationships between isolates, which confirmed previous serotyping results, are illustrated in Figure 1. Our analysis revealed two distinct clusters in the phylogenetic tree. The first cluster represented Salmonella Anatum isolates (n = 10), whereas the second cluster represented Salmonella Liverpool isolates (n = 12). Notably, within each serotype cluster, all isolates displayed a relatively low SNP difference number below 100. This finding indicates a potential common contamination source for the isolates within each serotype.
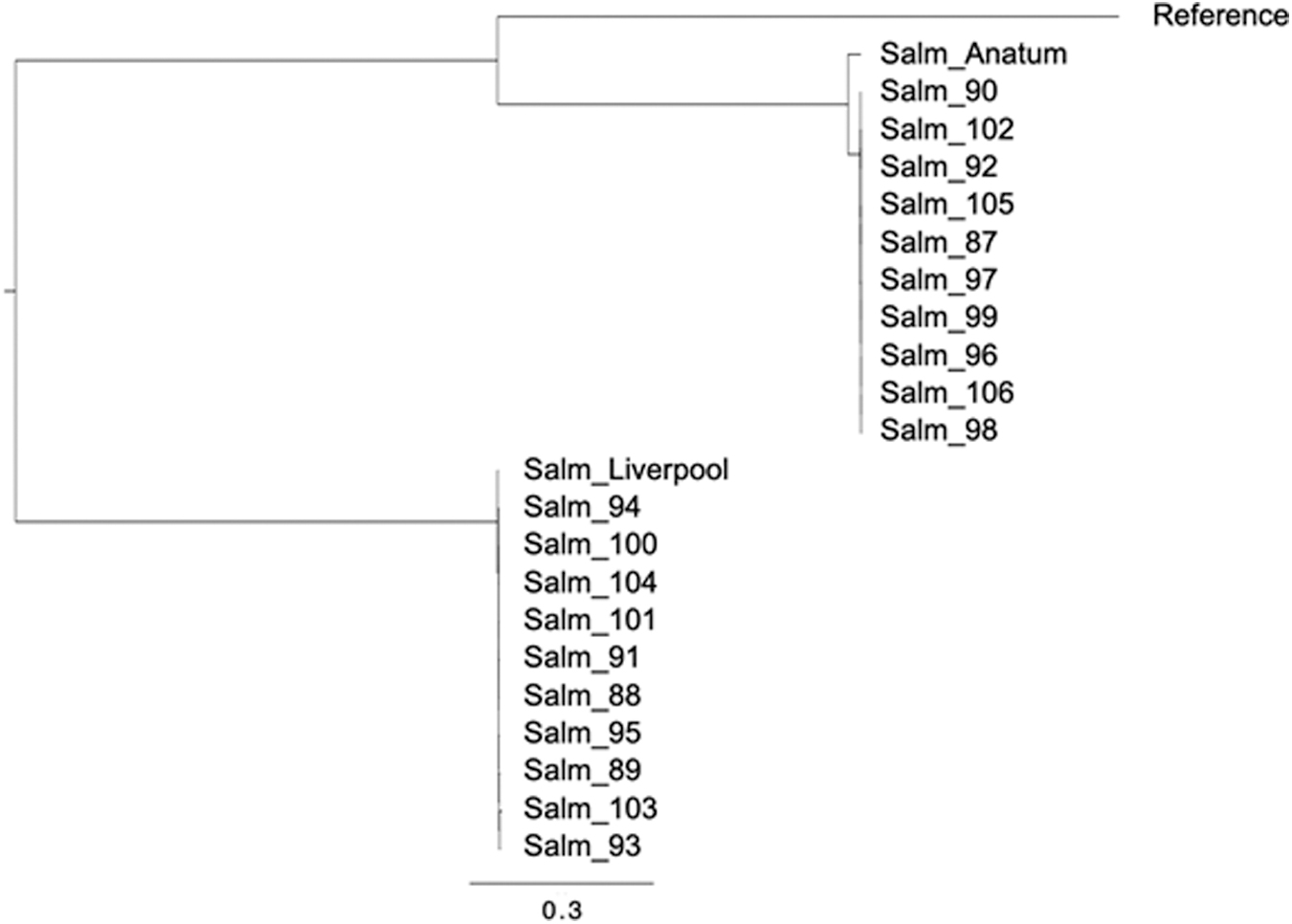
Phylogenetic tree based on chromosomic SNP differences between Salmonella isolates. SNP differences were generated using snippy v4.4.5 against reference genome Salmonella enteritidis ATCC BAA-708 (CP025554.1). The phylogenetic tree was created using Fasttree v2.1.10 and Figtree v1.4.4 software. Isolates Salmonella ser Anatum (CP045516) and Salmonella ser Liverpool (CP083758) were also used to confirm serotypes. Branch lengths represent the number of nucleotide substitutions per site (scale: 0.3 substitutions per site). SNP, single nucleotide polymorphism.
Antibiotic susceptibility testing and acquired resistance gene detection
All isolates were susceptible to all tested antibiotics except those of Salmonella Anatum, which were resistant to nalidixic acid, ofloxacin, and ciprofloxacin. The isolates showed ciprofloxacin MIC values of 0.08 mg/L (n = 3), 0.012 mg/L (n = 7), which are considered susceptible, and 0.019 mg/L (n = 4), 0.025 mg/L (n = 6), which are considered resistant to ciprofloxacin according to the CLSI clinical breakpoint (Table 1).
Upon further analysis through whole genome sequencing, all strains contained the chromosomal aminoglycoside resistance gene aac(6′)-Iaa. In addition, the plasmid-mediated quinolone-resistance gene qnrB19 was only identified among the Salmonella Liverpool serovar (n = 10). Furthermore, a point mutation was detected in the parC gene in the same position for all isolates (parC:p.T57S) (Table 1).
PlasmidFinder identified the presence of Col(pHAD28) plasmid in all Salmonella Liverpool strains. No plasmid was detected in the Salmonella Anatum isolates (Table 1).
Virulence genotyping
In this study, we investigated the presence of virulence genes in Salmonella Anatum and Salmonella Liverpool strains (Supplementary Table S1). Our findings revealed the detection of multiple common chromosomic operons and genes associated with virulence in both serovars. The presence of these genes suggests that they possess similar potential for pathogenicity and virulence.
Discussion
This study reported the presence of Salmonella strains in watermelons in Algeria. The study found that 19% of the watermelon samples analyzed contained Salmonella, highlighting the health risks of consuming fresh products. No Salmonella has been isolated from irrigation water and soil in the studied samples.
The prevalence of Salmonella in vegetables and fruits varies significantly across regions and countries. According to recent studies, the prevalence of Salmonella in vegetables and fruits is higher in Africa at 19.9% (Corredor-García et al., 2021). A recent systematic review detected a lower prevalence of 0.9% in China (Miao et al., 2021), whereas 17.5% of vegetable samples were positive in Vietnam (Nguyen et al., 2021). In China, the prevalence of Salmonella in foods from 2006 to 2016 ranks fifth among other pathogens (Paudyal et al., 2018).
In our study, we investigated the prevalence of Salmonella on fresh watermelon during the preharvest season. Our findings revealed the presence of Salmonella in watermelon samples, indicating a significant concern for food safety in the region. Although our study did not include comparative data from previous years or other regions, it is essential to note that, to the best of our knowledge, only one article has been published in Algeria on the presence of Salmonella in raw vegetables, which reported no Salmonella presence (Mesbah Zekar et al., 2017). Therefore, our study provides valuable additional information by highlighting the occurrence of Salmonella contamination, specifically in fresh watermelon.
Considering other sources of contamination, the study of Bounar-Kechich et al. (2012) showed Salmonella Heidelberg, Salmonella Enteritidis, Salmonella Albany, and Salmonella Typhimurium prevalence in poultry production farms. In humans, several nontyphoidal Salmonella, such as Salmonella Heidelberg, Salmonella Enteritidis, Salmonella Typhimurium, and Salmonella Anatum, have been found in Algeria (Djeghout et al., 2017; Kermas et al., 2012). The presence of Salmonella spp. in vegetable samples was detected in Morocco (Ibenyassine et al., 2007) but not in samples from Egypt (Abaza, 2017), Ghana (Abass et al., 2016), Burkina Faso (Dao et al., 2018), or South Africa (Richter et al., 2021).
Although meat products are considered high risk for Salmonella contamination, ready-to-eat vegetables and fruits, including watermelons, can also be a source of salmonellosis. Salmonella can be present on the surface of watermelon rinds and can spread to edible tissues when cut during preparation. Previous outbreaks of salmonellosis have been associated with watermelon, including precut-wrapped watermelon contaminated with Salmonella Miami and Salmonella Bareilly (GOLDEN et al., 1993).
In addition, a review of outbreaks reported by the Centers for Disease Control and Prevention's Foodborne Disease Outbreak Surveillance System in the United States from 1973 to 2011 identified 34 instances of foodborne diseases caused by various melons, including cantaloupes, watermelons, and honeydew. Salmonella was reported as the most frequent cause of these outbreaks (Chan et al., 2023).
Studies have identified several risk factors for vegetable contamination, including noncontrolled temperatures, human handling practices, soil and fertilizers applied using animal manure, wildlife, and irrigation water from both irrigation and non-irrigation sources (Corredor-García et al., 2021; Reddy et al., 2016). Climate change and global warming have been directly implicated in the increased spread of foodborne pathogens, including Salmonella.
Interestingly, no significant correlation was observed between precipitation and Salmonella infections, highlighting the specific influence of temperature changes in facilitating the transmission of this pathogen (Akil et al., 2014). Irrigation water is one of the most essential factors for Salmonella spp. contamination in vegetable growth in soil (Hernandez et al., 2021; Melloul et al., 2001). As watermelons are grown on the ground, their surfaces are in direct contact with soil or contaminated irrigation water, which is a possible risk factor for contamination (Walsh et al., 2014).
Data from the literature indicate that fresh fruits and vegetables can become contaminated with Salmonella through contact with soil, manure, compost, water, or personnel (Kowalska, 2023). The type of melon cultivar also plays a crucial role in Salmonella colonization (Korir et al., 2020). Cantaloupes melons or netted rind melons have been mainly linked to Salmonella widespread foodborne outbreaks that can be introduced through the water, the soils, or the flowers, resulting in both external contamination and internalization in the fruit (Burris et al., 2021).
Spinach has also described this process (Kumar et al., 2020). Precutting melons may facilitate additional opportunities for contamination and pathogen amplification (Chan et al., 2023). Slicing into a melon can transfer pathogens from the surface to the edible flesh, leading to cross-contamination of other melons (Ukuku et al., 2012). Bacterial pathogens can multiply on the edible flesh of cut melons held at room temperature. The risk of a family outbreak is also more critical as it is shared and consumed by one family. These findings highlight the need for rigorous safety protocols and targeted preventive measures throughout melons' production and supply chain to minimize the risk of Salmonella contamination.
This study identified two distinct Salmonella serovars: Salmonella Anatum and Salmonella Liverpool. We assigned Salmonella Anatum to ST64 and found 6074 records in Enterobase from various sources, including 59 records from food. For Salmonella Liverpool, we identified strains as ST1959 and found 392 records through a quick search on Enterobase, including only 2 records from food (https://enterobase.warwick.ac.uk/species/senterica).
These two serotypes have been previously associated with both clinical and foodborne illness cases, indicating that they are potentially essential pathogens of public health concern (Quiroz-Santiago et al., 2009). Isolates of the same serovar had a robust genetic relationship, suggesting they belonged to the same clone (SNP difference <100).
Nevertheless, the source of contamination is still ignored, as no Salmonella was isolated in the analyzed irrigation water and the soil during the study. This suggests that the contamination could be attributed to other sources or occurred before sampling. It should also be cautioned that these data do not imply that the absence of Salmonella is related to organic agricultural practices.
Antimicrobial resistance in bacteria from food can lead to resistant strains in humans (Miranda et al., 2009; Pan et al., 2019). However, little is known about the prevalence of antimicrobial-resistant Salmonella strains in fruits and vegetables (Reddy et al., 2016). In this study, Salmonella isolates from watermelons showed a low level of antibiotic resistance, with no resistance to most antibiotics tested. Quinolone resistance was observed in the Salmonella Liverpool isolates. Similarly, in a study by Miranda et al. (2009), high levels of quinolone resistance were found in isolates from vegetable samples.
We found that the phenotypic susceptibility patterns were consistent with the genotypic analysis. First, we revealed the presence of the aac(6′)-Iaa gene in all sequenced Salmonella strains. This later is naturally present in all those serovars and rarely confers phenotypic resistance to aminoglycosides (Neuert et al., 2018). Moreover, Col(pHAD28) plasmid was found in all Salmonella Liverpool, which was reported by various authors harboring different PMQR genes such as qnrS1 and qnrB19 (Hurtado et al., 2022; Li et al., 2021; Raufu et al., 2021).
Quinolone resistance in Salmonella is increasing worldwide and has been linked to the use of fluoroquinolones in food animals. The higher levels of resistance to nalidixic acid, ofloxacin, and ciprofloxacin in Salmonella isolates from watermelons may be due to the presence of the qnrB19 gene as previously reported in the literature (Ballesteros-Nova et al., 2022; Dionisi et al., 2009; Pachanon et al., 2020). Mutation T57S in the parC gene was not associated with quinolone resistance, as it has already been described in other studies (Chang et al., 2021; Qian et al., 2020).
Salmonella's pathogenicity depends on various factors, such as the serotype and the host's immune response. Many virulence genes have been identified in different serovars in pathogenicity islands, such as SPI-1 and SPI-2. These genes are essential for invasion and survival within host cells. Salmonella can also infect plants, and plants possess antibacterial peptides that protect them from bacterial infections, but it is unclear how effective they are against Salmonella. Understanding the complex factors influencing Salmonella's pathogenicity and interactions with hosts is critical in developing effective strategies for the prevention and treatment of infections (Grivokostopoulos et al., 2022; Grund and Weber, 1988; de Moraes et al., 2017).
The comparison of the virulomes of Salmonella Anatum and Salmonella Liverpool strains revealed the shared presence of a wide range of virulence genes and operons. These genes encode factors associated with adhesion, biofilm formation, toxin production, invasiveness, and other virulence mechanisms. The similarity in the virulence gene profiles suggests that both serotypes have comparable capabilities to cause disease and establish infections. These findings contribute to our understanding of the genetic determinants underlying the pathogenicity of Salmonella strains and their potential impact on human health.
Our study's main limitation was the relatively small sample size and the concentration of sampling locations in two farms. Although the sample size of 105 watermelons may seem limited, it was chosen to provide a representative, practical, and manageable data set for comprehensive analysis within the scope of our study. The choice of farms was strategic, focusing on significant contributors to watermelon production in the Bejaia region.
Another limitation of this study was the absence of identification of contamination sources despite soil and water analysis. More samples or other matrices, such as animal-based soil amendments or air, could help identify the contamination source. These types of samples should be performed in future studies. Moreover, we could not directly evaluate the contamination risk as watermelon samples during the preharvest season.
The Salmonella contamination could have originated from various potential sources, including contaminated irrigation water, soil, equipment, and air, as well as from human or animal fecal contamination. The fact that Salmonella was not detected in the water samples suggests that the contamination source may have been localized and limited to the watermelon crop. Investigating the factors that led to the contamination is essential to prevent future occurrences. This may involve examining the growing and handling practices in the farms and implementing appropriate interventions to minimize the risk of contamination.
Salmonella spp. is a significant challenge to control because it can contaminate fresh fruits and vegetables at any stage of the supply chain, from growth and harvesting to transportation and marketing, and is challenging to remove (Corredor-García et al., 2021). To minimize the risk of transmission of pathogens in fresh fruits and vegetables, implementing Good Agricultural Practices (GAPs) is crucial.
GAPs focus on reducing microbial hazards from seed to production and require incorporating measures such as record-keeping, ensuring worker health, hygiene, and training, avoiding the use of manure for soil amendments, ensuring the quality of water for production and post-harvest, controlling wildlife interaction with production areas, implementing cleaning and sanitization, pest control in packinghouses, and traceability (Bihn and Reiners, 2018). Implementing these practices can significantly reduce the incidence of Salmonella spp. in fresh fruits and vegetables, which is essential to protecting public health.
Conclusion
This study has demonstrated the presence of Salmonella in watermelon in Bejaia, Algeria, with the majority of isolates exhibiting susceptibility to the tested antibiotics except for quinolones. Our study gives new data on watermelon samples and indicates a relatively high prevalence of Salmonella before harvest. Although we cannot directly compare our results with previous studies, it is crucial to recognize that the absence of comprehensive comparative data underscores the need for further research and surveillance.
However, despite low antimicrobial resistance rates, Salmonella in watermelons poses a severe risk to public health as it is widely distributed, consumed raw, and produced in large quantities. The Salmonella serotypes detected in this study are also commonly associated with other foodborne illnesses. Therefore, it is imperative to continue monitoring the microbiological quality of watermelon to enhance our understanding of the potential risks associated with its consumption. These findings underscore the need for ongoing efforts to ensure the safety of fresh produce and protect public health.
Declaration of Interests
The authors have no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the article.
Footnotes
Disclosure Statement
No competing financial interests exist.
Authors' Contributions
Z.B. collected the samples, conducted the analysis, and wrote the article. S.M. performed the whole genome sequencing, analyzed the data, and corrected the article. V.T. contributed to the bioinformatic analysis. A.M. analyzed the microbiological data. C.D.C. conducted the antibiotic testing and also corrected the article. O.B. supervised, reviewed, and edited the article. A.T. directed, supervised, and corrected the article. All authors have read and approved the final version of the article.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.