
Abstract
Milk is an important source of food, and it is also a nutrient-rich medium, which can harbor multiple microorganisms. Staphylococcus aureus is an important foodborne pathogen in food-producing animals, and there have been many reports on its infection and antimicrobial resistance (AMR), which has significant global public health concerns. This study was designed to isolate, characterize, and analyze the AMR pattern of S. aureus from milk samples collected in Chennai, India. A total of 259 raw milk samples from 3 groups: dairy farms, local vendors, and retail outlets were analyzed, and it was found that 34% (89/259) were positive for S. aureus. Positive isolates were further characterized by pulsed-field gel electrophoresis and isolates recovered from different sources, study areas, and locations showed high genetic diversity with no similarity. The presence of AMR has been further assessed by phenotypic methods as per CLSI-M100 performance standards, and all the isolates were susceptible to ampicillin/sulbactam, mupirocin, and tylosin. Additionally, all of the isolates were resistant to ampicillin. There were 28 isolates categorized as multidrug-resistant, which showed resistance to more than 2–3 classes of antimicrobials. This is the first report of inducible clindamycin resistance and mupirocin sensitivity pattern from S. aureus isolates recovered from milk. This study established the occurrence varied with genetic diversity in the isolates prevalent in the study area and divergence pattern of AMR S. aureus. The AMR in these isolates and with methicillin-resistant S. aureus could pose a serious threat to food safety and economic implications.
Introduction
S
The prophylactic and indiscriminate use of antibiotics in agriculture, fisheries, and livestock has led to the emergence of methicillin-resistant S. aureus (MRSA) and other antimicrobial-resistant bacteria, which are serious public health concerns as they compromise antimicrobial treatment in staphylococcal infection in humans (Guo et al., 2020; Vivas et al., 2019). In 2017, the World Health Organization (WHO) published a list of global priority pathogens in which MRSA infections in humans has been categorized as high-risk status (Asokan et al., 2019). Furthermore, multidrug resistant S. aureus has become a serious health, economic, and environmental problem (Vivas et al., 2019). Increase in number of antimicrobial resistant S. aureus in milk, meat, and foods of animal origin is an alarming threat to humans, animals, and the environment. The MRSA acquiring various resistance mechanisms and being a very complex pathogen is becoming clinically problematic worldwide (Argudín et al., 2010; Guo et al., 2020) and can be found in livestock and in the environment. In 2009, the European Food Safety Authority underlined the increasing public health issues represented by the presence of MRSA in food-producing animals and recommended that future research should be performed on sampling, detection, and quantification of MRSA carriage in both humans and animals, as well as on the foods of animal origin and their environment (EFSA, 2009).
In 2019, a new antimicrobial resistance (AMR) indicator was included in the Sustainable Development Goals monitoring framework and with the data provided with Global Antimicrobial Resistance and Use Surveillance System (GLASS). The GLASS factsheet on AMR documented that the clinical blood-stream infections due to MRSA across 49 nations was 12.11% (interquartile range 6.4–26.4) and also stated that people with MRSA infections are 64% more likely to die than people with drug-sensitive infections (WHO, 2021). It is necessary to develop strategies concerning both reducing the burden of spread of infectious agents and mitigating the spread of AMR. To achieve this, the One Health approach was highlighted for disease investigation and control (Guardabassi et al., 2020). Hence, it is essential to monitor the prevalence and AMR pattern among S. aureus as the organism. It is commonly encountered when switching hosts between animals and humans, leading to the emergence of newer strains, which may be pathogenic and/or AMR clones (Rainard et al., 2018). Our research is a small step with concerning foods of animal origin or animal agriculture, which may help in devising effective mitigation strategies. This will ensure safety across the food chain. Hence, this study was conducted to investigate the prevalence and characterization of antimicrobial-resistant S. aureus isolated from raw milk samples in Chennai, India.
Materials and Methods
Milk sample collection
The milk samples for the current study were collected from various raw milk retail outlets, vendors, and dairy farms in and around Chennai city, India (13.0827° N, 80.2707° E). A total of 259 raw milk samples were collected for this study, which includes 109 samples collected from small dairy farms, 60 samples from local vendors who sell milk early morning directly to customers, and 90 samples from retail outlets who sell milk in sealed packs early morning directly to customers. For each individual sample, 25 mL of milk was collected in a sterile screw cap bottle and immediately transported in a refrigerated chain to the laboratory (Kou et al., 2021).
Isolation and identification of S. aureus
Isolation of S. aureus was done as per the standard procedure (ISO standard 6888/1:1999 and 6888/2: 1999) and with small modification as described in Savariraj et al. (2019). In brief, raw milk samples were enriched on brain heart infusion (BHI) broth (HiMedia, India) with 7% NaCl (sodium chloride) overnight. The selective plating for S. aureus was done on Baird Parker Agar, containing potassium tellurite and mannitol salt agar (HiMedia). The isolated colonies were Gram-stained and subjected to biochemical tests (catalase test, coagulase test, hemolysis test, and HiStaph™ Latex Test Kit; Himedia) for initial confirmation. For further confirmation, thermonuclease (nuc) gene PCR assay was performed (Hegde et al., 2013). Unless otherwise mentioned, the S. aureus strain ATCC 25923 was used as the reference strain for the entire experiment.
Pulsed-field gel electrophoresis
The pulsed-field gel electrophoresis (PFGE) procedure was done as per previously published protocol (Golding et al., 2015) and performed following the Center for Disease Control and Prevention PulseNet standardized procedure. For each test, single colony that was isolated from selective medium was inoculated into 5 mL of BHI broth and incubated with moderate shaking at 35–37°C for 24 h. After incubation, the suspension was centrifuged, and the pellet was adjusted with sterile BHI broth to a turbidity reading of 1.1–1.3. Then, 200 μL bacterial suspension of brain heart infusion broth was centrifuged at 12,000 × g for 2–4 min, and the supernatant was aspirated. The pellet was resuspended in 200 μL of Tris-ethylenediaminetetraacetic acid (TE) buffer (10 mM Tris HCl, 1 mM EDTA [pH 8]) and equilibrated in a 37°C water bath for 10 min. To this, 4 μL of conventional (No. L-7386; Sigma) lysostaphin stock solution (1 mg/mL in 20 mM sodium acetate [pH 4.5]) was added, and 1 mL of 1.8% (wt/vol) SeaKem Gold agarose (Seakem gold®; Bio-Rad) in TE buffer (equilibrated to 55°C) was added to the cell suspension, gently mixed, and dispensed into a small disposable mold (∼100 μL each). The plugs were allowed to solidify at room temperature for 10–15 min or in the refrigerator (4°C) for 5 min. The plugs were removed and placed into a tube containing at least 3 mL of EC lysis buffer (6 mM Tris HCl, 1 M NaCl, 100 mM EDTA, 0.5% Brij-58, 0.2% sodium deoxycholate, 0.5% sodium lauroylsarcosine) and incubated at 37°C for overnight. The EC lysis buffer was poured off and washed with 4 mL of TE buffer for at least three more times, and the plugs were stored at 4°C. The plugs were digested with SmaI restriction enzyme (New England Biolabs, Inc., Ipswich) at room temperature for 4 h.
The PFGE typing was carried out in a CHEF-DRII apparatus (Bio-Rad Laboratories, Des Plaines). The Saccharomyces cerevisiae (Bio-Rad) chromosomal DNA varies between 0.225 and 2.2 Mb of predigested genomic plugs, and this was used as a clamped homogeneous electric field (CHEF) DNA size marker or PFGE standards. The 1% agarose gel was prepared and plugs were loaded and electro-phoresed with positioning in a contour-CHEF (Bio-Rad) tank and submerged in 1700 mL of 0.5 × Tris-Borate-EDTA. The total run time was 21 h at 14°C with an initial pulse time of 5 s, a final pulse time of 40 s, and a voltage of 6 V/cm or 200 V. The gels were then stained with 1.5 μg/mL ethidium bromide solution (AMRESCO X328, 10 mg/mL; Amresco, Inc., Solon, OH) for 20 min in a covered container and de-stained in sterile distilled water for 45 min. The gels were visualized and photographed in the Gel Documentation System (Alpha Imager; ProteinSimple, CA), and images were saved as a TIFF file for analysis. The PFGE patterns were generated by the typing method and were analyzed using BioNumerics software 7.6 (Applied Maths, Belgium). To generate a dendrogram with Dice's similarity coefficient, the unweighted pair group method with arithmetic means (UPGMA) was used, and clusters were selected using a cut-off at the 80% level of genetic similarity (McDougal et al., 2003). The random stratified selection method was used for selection of 40 isolates from 3 groups: dairy farms, local vendors, and retail outlets for further analysis.
Antimicrobial susceptibility testing
The phenotypic characterization to assess the antimicrobial-resistant pattern of 89 S. aureus milk isolates (dairy farm n = 37, local vendor n = 28, retail outlet n = 24) was subjected to antibiotic susceptibility test against commercial antimicrobial disks (ampicillin-10 μg, ampicillin/sulbactam, amoxicillin-10 μg, ciprofloxacin-5 μg, co-trimoxazole (trimethoprim-sulfamethoxazole—1.25/23.75 μg)-25 μg, tetracycline—30 μg, amikacin—30 μg, azithromycin—15 μg, cefotaxime—30 μg, clindamycin—2 μg, erythromycin—15 μg, imipenam—10 μg, rifampicin—5 μg, teicoplanin—30 μg, and tylosin—10 μg) (HiMedia). For detection of high-level mupirocin resistance, mupirocin 200 μg antimicrobial disks were used (HiMedia). The procedure of the above tests and interpretation of results was carried out according to Clinical Laboratory Standard Institute [31th ed. CLSI supplement M100] (CLSI, 2021).
The evaluation of β-Lactamase production by S. aureus isolates was carried out using penicillin 10 μg antimicrobial disk (HiMedia) (penicillin zone edge test). For methicillin resistance, MRSA was carried using cefoxitin 30 μg (HiMedia). Inducible clindamycin resistance was detected with a D-test using the clindamycin 2 μg + erythromycin 15 μg antimicrobial disk (HiMedia). The procedure of the above tests and interpretation of results was carried out according to Clinical Laboratory Standard Institute [31th ed. CLSI supplement M100] (CLSI, 2021).
Statistical analysis
Statistical analysis was performed using SAS (version 9.4; SAS Institute, Inc., Cary, NC) LOGISTIC and FREQ procedures. The S. aureus prevalence in 259 raw milk samples was analyzed using the logit linear model with fixed effects being sample source (dairy farm, local vendor, and retail outlet) and milk producer (1, 2, 3, 4) nested within dairy farm. The producer-nested-within-dairy-farm effect was not significant (p = 0.114); hence, it was excluded from the subsequent analyses of the 89 milk samples that tested positive for S. aureus. The standard error was at 5%, and an alpha value of 0.05 was used in this study. The antimicrobial susceptibility of S. aureus in milk samples was analyzed separately for each phenotypic characteristic using the logit linear model with the fixed effect being sample source. Exact inference was applied in the case of sparse (i.e., <5) resistant or susceptible milk samples. All tests were performed at a 5% standard error and alpha value of 0.05. Pairwise comparison of source effects was carried out based on the two-sided test for nonzero log odds ratio.
Results and Discussion
Prevalence of S. aureus
The best tools for laboratory isolation and identification of S. aureus are through classical methods, which include the collected milk samples with enrichment, selective plating, and biochemical test. A total of 89 positive S. aureus isolates were isolated from 259 milk samples (89/259: 34.4%) (34.4 ± 0.609; 95% confidence interval) and were screened based on the cultural dependent method. The isolates were further confirmed with biochemical tests and PCR assay against the nuc (thermonuclease) gene. The prevalence details are summarized in Table 1. The odds ratio directly implies that the prevalence of S. aureus in the study area is directly impacted by the source of collection. From the logistic model, the odds of recovery of S. aureus were 2.41 times more likely in local vendors compared to retail sources, similarly to 1.40 between dairy farm and the retail outlet. The results of our study with reference to the prevalence of S. aureus from milk samples were in accordance with various researchers who have recorded results around the globe. Traversa et al. (2015) reported 41.0% in milk from Italy; Sudhanthiramani et al. (2015) reported 39.09% in milk from Tirupathi, India, respectively. In contrast, a higher prevalence of S. aureus was reported in China (46.2%) (Wang et al., 2018) and Norway (75%) (Jørgensen et al., 2005), respectively. Variation in the prevalence of S. aureus may be attributed to various factors such as sampling method, period of sampling, number of samples screened, diagnostic procedures, hygienic conditions prevailing at retail outlets, and geographical location. The recorded occurrence of S. aureus in this study underlines the need for good hygiene practices during transportation and handling at retail outlets to reduce the risk of transmission of S. aureus to humans.
Prevalence of Staphylococcus aureus in Milk Samples
A, B, AB denotes the statistical correlation between the other 2 groups when tested.
S.E., standard error.
Pulsed-field gel electrophoresis
The pulsed-field gel electrophoresis is considered as the important method, which has been used for S. aureus strains due to its high discriminatory power and reproducibility. The 40 representative S. aureus milk isolates were typed using PFGE, of which only 33 (82.5%; 33/40) were typed, which includes 12 from dairy farms, 8 from local vendors, 13 from retail outlets, and 7 other isolates that were nontypeable by this method. The genetic analysis revealed different DNA banding patterns of varying distinct bands, showing high genetic diversity among the strains. The dendrogram of PFGE analysis of S. aureus isolates revealed 5 major clusters (namely A, B, C, D, E) and 27 pulsotypes, which denotes high genetic diversity and no specific or clonal lineages to an area and/or source of collection among the samples (Fig. 1). The cluster B and C had highest agglomeration of isolates from all three study sampling groups. The isolates from three groups were distributed among all five clusters, which suggest high similarity between the isolates of different groups made up of highly variable genetic makeup. The PFGE analysis of this study is concurrent with several other studies, such as Dorneles et al. (2018), which reported 34 different genotypes from 79 S. aureus milk isolates. Papadopoulos et al. (2018) characterized 11 MRSA isolates and grouped them into in 4 different clusters with 6 pulsotypes, with 1 cluster containing 7 MRSA strains (63.6%). These isolates were obtained from food handlers, equipment, and milk isolates, suggesting that similar organisms exist throughout a system. In contrast, Alba et al. (2015) in his review suggests the association with cattle and human S. aureus isolates, and showed high genetic relatedness for cattle and human isolates with MRSA isolates. The results from other studies concurred with our results, which represent that high genetic dissimilarity and belong to the many pulsotypes even though the source of collection is the same. As reported previously, the genetic variability among S. aureus isolates may be attributed to differences in geographical origin and collection site/source of isolates. In this study, there are few overlapping genotypes that belong to the same pulsotype and were isolated from three sources of collection, which is helpful in correlating the genetic cross over among different strains, with respect to location or collection source. This clearly indicates that the presences of S. aureus in the study area is not specific to an animal, region, or source, corresponding with the suggestion that organisms are highly variable with changing genotypic patterns.
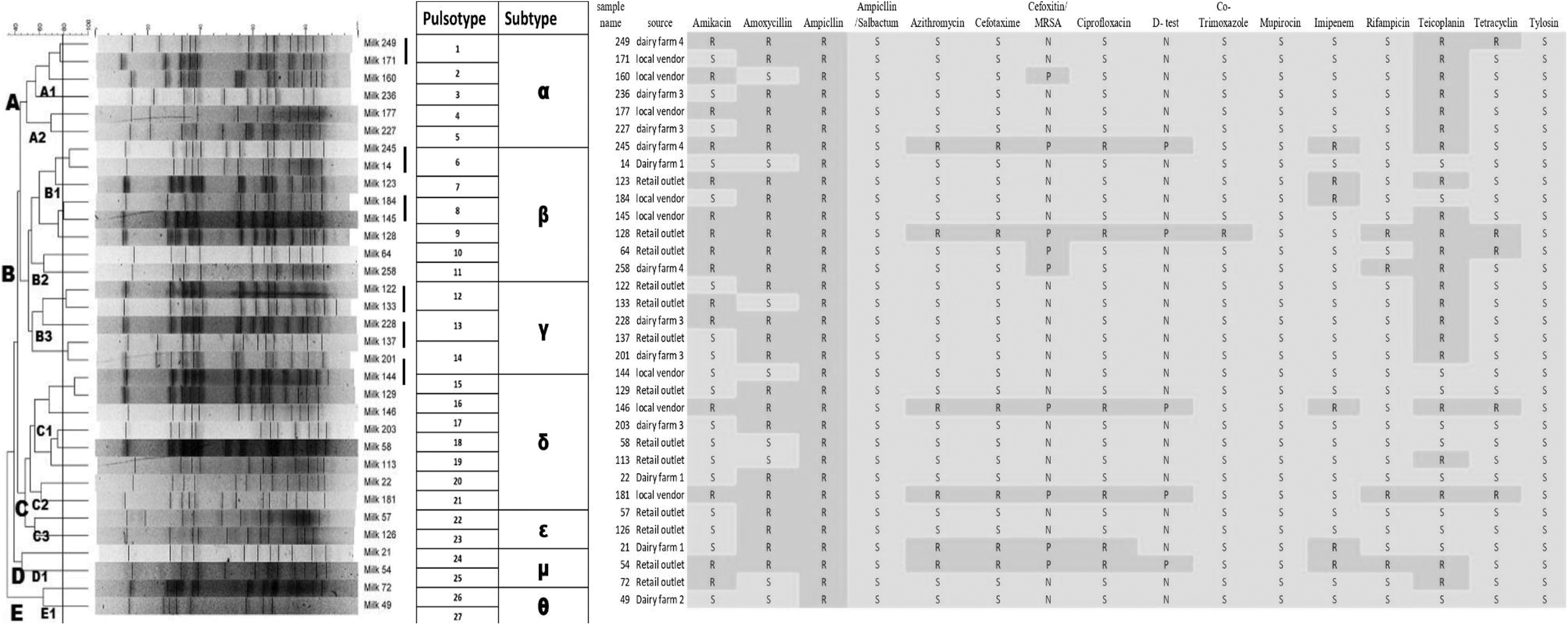
The PFGE and heat map of phenotypic-resistant markers among Staphylococcus aureus strains isolated from milk samples. PFGE, pulsed-field gel electrophoresis. R, resistant; S, sensitive.
Antimicrobial resistance: S. aureus
The phenotypic characterization was carried out using the disk diffusion method, and results of our study are summarized in Table 2, which shows the resistance rate and its odds ratio. Most of the tested antimicrobials have a p-value of >0.05, which indicates the resistance pattern is not much affected by sampling group, source, or test antimicrobial. All the isolates were sensitive to ampicillin/sulbactam, mupirocin, and tylosin. However, all the isolates were resistant to ampicillin. The resistance pattern with amikacin (aminoglycodsides group) shows an ∼50% resistance when compared to the entire isolates and odds ratio represents the direct positive relationship with source of collection. The PFGE representative samples heat map of AMR S. aureus given in Figure 1, amoxycillin (β-lactam/penicillin group) shows more than 75% resistance. Azithromycin (macrolide group) shows around 20% resistance. Cefotaxime (cephalosporins group) shows around 15% resistance and odds ratio correlating with source of collection. Ciprofloxacin (quinolones group) shows around 15% resistance and odds ratio correlating with source of collection. With co-trimoxazole (sulphonamides group), a low resistance pattern was noticed. Imipenem (carbapenams group) shows around 12% resistance. Rifampicin (ansamycins [antimycobacterial] group) shows around 15% resistance, and Teicoplanin (glycopeptides group) shows more than 50% resistance. Tetracycline (tetracycline group) shows more than 10% resistance and an odds ratio correlating with the source of collection.
The Phenotypic Characterization of Antimicrobial Resistance in Isolates of Staphylococcus aureus
A denotes the statistical correlation between the other groups when tested.
Out of 89 samples.
Exact inference.
Median unbiased estimate.
AMR, antimicrobial resistance; MRSA, methicillin resistant S. aureus; S.E., standard error.
The 28 milk S. aureus isolates showed multiple drug resistance by being resistant to more than 2 groups of classes of antimicrobials. (The complete data of 89 isolates and AMR S. aureus were showed in Supplementary Table S1 and Supplementary Fig. S1.)
Joshi et al. (2014) reported that S. aureus milk isolates were found sensitive to ciprofloxacin (97.47%), gentamicin (94.95%), ceftriaxone (91.59%), tetracycline (89.91%), and cefoxitin (62.18%) antibiotics. Similarly, Thaker et al. (2013) documented a sensitivity pattern with S. aureus milk isolates, cephalothin (100%), co-trimoxazole (100%), cephalexin (100%), methicillin (100%), gentamicin (90%), ciprofloxacin (80%), oxacillin (70%), streptomycin (60%), ampicillin (60%), and penicillin-G (0%). In contrast to our results, Liu et al. (2017) documented that 72% of isolates were multidrug-resistant. Further, Sharma et al. (2017) reported that 90% strains were multidrug-resistant, showing resistance toward penicillin-G (100%), ampicillin (77%), amikacin (57%), oxacillin (50%), ciprofloxacin (40%), and azithromycin (30%). In India, Mahanti et al. (2019) reported that AMR levels for different antimicrobials in milk samples are ceftizoxime (90.5%), ampicillin (71.4%), cefoxitin (42.9%), amoxiclav (38.1%), and erythromycin (38.1%). Furthermore, all the tested MRSA isolates were multidrug-resistant.
The presence of MRSA isolates in foods of animal origin may act as routes of exposure and lead to human transmission (Haran et al., 2012; Kaszanyitzky et al., 2004; Normanno et al., 2007). The MRSA from milk samples was found to be 27% (24/89). Several authors endorsed that the cefoxitin disc method has better sensitivity than the oxacillin disc method for MRSA detection (Boutiba-Ben Boubaker et al., 2004; Skov et al., 2003; Velasco et al., 2005). This cefoxitin has an increased affinity for expression of the mecA-encoded protein PBP2a, and also acts as an inducer of the mecA gene (Velasco et al., 2005). Indian Council of Medical Research (ICMR) AMR-Antimicrobial Resistance Research & Surveillance Network annual report in 2021 stated that MRSA rates are increasing each year from 2016 to 2021 (28.4–42.6%) (ICMR, 2021).
The penicillin resistance and penicillinase detection for β-lactamase production phenotypically was through the production of the haze zone around the penicillin-G, or the disc resistance area. Our study documented that 8% (7/89) of isolates were positive for β-Lactamase production with a haze zone. Kaase et al. (2008) has reported penicillinase detection by starch–iodine plate method; nitrocefin tests; cloverleaf assay; penicillin zone-edge. Robles et al. (2014) reported that 83% of isolates were resistant to penicillin by disc diffusion method. The high resistance to penicillin through disc diffusion assay has been reported in many studies from foods of animal origin (Liu et al., 2017; Rai and Tiwari, 2016). The 100% resistance for penicillin by disc diffusion assay may be attributed to the presence of additional protein with a reduced affinity for penicillin or other β-lactam antibiotics (Brown and Reynolds, 1980). Mahanti et al. (2019) reported antimicrobials in milk samples and also the co-resistance to tetracycline, macrolide, and aminoglycoside in addition to the β-lactams.
This study has identified inducible clindamycin resistance (D-test for clindamycin-erythromycin test) in 13% (12/89) of milk S. aureus isolates from milk samples in the study area. Earlier, the inducible clindamycin resistance was commonly reported from hospital and clinical settings. To our knowledge, this is the first report of clindamycin resistance related to a food of animal origin. Adhikari et al. (2017) reported 10% inducible clindamycin resistance from clinical samples; Prabhu et al. (2011) documented 12.4% inducible clindamycin resistance from hospital settings. The differentiation of inducible clindamycin-resistant phenotypes from others is crucial for therapeutic implications with clindamycin (Carter et al., 1985). The presence of erythromycin-induced clindamycin resistance in milk samples needs due attention, as it indicates dissemination of the resistance into foods of animal origin besides the commonly reported cases in clinical or hospital settings. Results of our study documented that four milk samples showed resistance, barring a few isolates with a zone of inhibition of 10–12 mm.
Mupirocin is a nonsystemic antibiotic that is occasionally used topically in animals and is used commonly against S. aureus for humans. It has a high degree of activity against all staphylococci, including MRSA (Carter et al., 1985). The development of resistance may have been linked to acquisition of resistance from plasmids (Rahman et al., 1987). Our results showed that there is no mupirocin resistance in foods of animal origin from the study area, but there is a chance in the near future that the resistance may be acquired from environmental contamination and pressure, as a few isolates showed a reduced zone of inhibition and fuzzy outer layer, which needs to be viewed with caution.
AMR is a major public health problem in many countries due to the persistent circulation of resistant strains of bacteria in the environment and animals. Many reports have suggested that resistance toward the less commonly used antibiotics and frequent changes in AMR patterns may be postulated as reasons for sensitivity and resistance patterns encountered with S. aureus isolates collected from different geographical foods of animal origin, and it is also well documented by various workers (Enright et al., 2002). In this study, we have identified the multiple drug-resistant AMR pattern among the isolates and with a diversified PFGE pattern with no specific similarity. Sources of collection touches on a newer dimension of microbial stress that needs to be focused on. This pattern of occurrence may raise the need for differentiating S. aureus and may also require novel tools in the future.
Conclusions
The occurrence of S. aureus in raw milk samples is an indication of potential danger prevailing in milk supply chains in our study area. Genotyping of the isolates by PFGE was helpful to understand the prevalent clonal lineages, which denote high genetic diversity and are unspecific to the source or geographical area of collection. The presence of multiple drug-resistant AMR patterns among S. aureus in milk samples are an indication of potential public health threats. Our study findings warrant the need for effective new control strategies that could be devised to counteract higher genetic diversity within MRSA and multiple drug-resistant AMR patterns among S. aureus that are prevalent from the study area.
Footnotes
Acknowledgments
The author acknowledges the Tamil Nadu Veterinary and Animal Sciences University, Chennai, India, for providing the facilities for the smooth conduct of the research. This manuscript is part of the PhD thesis submitted by the first author to TANUVAS, Chennai.
Authors' Contributions
Conceptualization, P.K., D.S.J., W.R.S., E.A., and S.S.; Formal analysis, D.S.J., P.K., W.R.S., S.G., T.M.A.S.K., N.B.R., Q.K. and R.G.A.; Investigation, D.S.J., P.K., S.S., and W.R.S.; Supervision, P.K., E.A., and S.S.; Writing—original draft, D.S.J., P.K., and R.G.A.; Writing and reviewing and editing, D.S.J., P.K. and R.G.A. All authors have read and agreed to the published version of the manuscript.
Disclosure Statement
No competing financial interests exist.
Funding Information
The research was supported by Tamil Nadu Veterinary and Animal Sciences University, Chennai, India.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.