
Abstract
Methicillin-resistant Staphylococcus aureus (MRSA) constitutes an important cause for concern in the field of public health, and the role of the food chain in the transmission of this pathogen and in antimicrobial resistance (AMR) has not yet been defined. The objectives of this work were to isolate and characterize coagulase-positive Staphylococcus (CoPS) and coagulase-negative Staphylococcus (CoNS), particularly S. aureus, from school dining rooms located in Argentina. From 95 samples that were obtained from handlers, inert surfaces, food, and air in 10 establishments, 30 Staphylococcus strains were isolated. Four isolates were S. aureus, and the remaining ones (N = 26) belonged to 11 coagulase-negative species (CoNS). The isolates were tested for susceptibility to nine antibiotics. The presence of genes encoding toxins (luk-PV, sea, seb, sec, sed, and see), adhesins (icaA, icaD), and genes that confer resistance to methicillin (mecA) and vancomycin (vanA) was investigated. The resistance rates measured for penicillin, cefoxitin, gentamicin, vancomycin, erythromycin, clindamycin, levofloxacin, trimethoprim–sulfamethoxazole, and tetracycline were 73%, 30%, 13%, 3%, 33%, 17%, 13%, 7%, and 7% of the isolates, respectively. Seventeen AMR profiles were detected, and 11 isolates were multidrug resistant (MDR). Seven methicillin-resistant Staphylococcus isolates were detected in the hands of handlers from four establishments, two of them were MRSA. Two S. aureus isolates presented icaA and icaD, another one, only icaD. The gene vanA was found in two isolates. In relation to S. aureus, resistance to vancomycin but not to gentamicin was detected. School feeding plays a key role in the nutrition of children, and the consumption of food contaminated with MRSA and vancomycin-resistant S. aureus (VRSA) can be a serious threat to health. In particular, it was detected that the handlers were the source of MRSA, VRSA, MR-CoNS (methicillin-resistant coagulase-negative Staphylococcus), and MDR isolates. The results obtained indicate that the vigilance of this pathogen in school dining rooms should be extreme.
Introduction
Staphylococcal foodborne intoxication is one of the leading causes of foodborne diseases worldwide, resulting, mostly, from the consumption of foods contaminated with enterotoxin-producing Staphylococcus aureus (Mahros et al., 2021). S. aureus, an opportunistic pathogen, is part of the skin and mucous microbiota of humans and can survive on surfaces and foods, colonizes, and causes serious infections in humans, ranging from skin and soft tissue infections to life-threatening septicemia and toxin-mediated diseases (Wu et al., 2019).
Moreover, methicillin-resistant Staphylococcus aureus (MRSA) constitutes an important cause for concern in the field of public health, and the role of the food chain in the transmission of this pathogen and in antimicrobial resistance (AMR) has not yet been defined (Lakhundi and Zhang, 2018). MRSA is resistant to penicillin (PEN) and all β-lactam antimicrobial drugs (Stefani and Goglio, 2010). MRSA can be determined phenotypically by antimicrobial susceptibility testing to cefoxitin (FOX) and/or oxacillin and genetically by the detection of mecA. The mecA gene, located on one of at least six types of staphylococcal chromosomal cassettes (SCCmec), encodes for a modified penicillin-binding protein (PBP2a) that leads to AMR. Frequently, MRSA with the mecA gene is resistant to a variety of antimicrobials (Sadat et al., 2022).
Vancomycin (VAN) remains as one of the first-line drugs for the treatment of MRSA infections. However, vancomycin-resistant Staphylococcus aureus (VRSA) isolates have emerged in recent years. VAN resistance in S. aureus is mediated by vanA gene cluster, which has been transferred from VAN-resistant Enterococcus (Cong et al., 2020).
S. aureus is able to encode virulence factors such as superantigenic toxins, namely the classical thermostable staphylococcal enterotoxins (SEA, SEB, SEC, and SED) (Fraser and Proft, 2008). Also, some MRSA frequently carry the cytotoxic toxin Panton–Valentine leukocidin (PVL) as a virulence factor associated with more severe clinical manifestations, although conflicting data exist on the role of PVL in the pathogenesis (Saeed et al., 2018). PVL toxin is a potent staphylococcal exotoxin killing immune cells and causing tissue necrosis (Miller et al., 2005).
Coagulase-negative Staphylococci (CoNS) constitute the main microbiota of the skin. These pathogens were underestimated and are not among the classical food poisoning bacteria, yet there are reports of their association with intoxication. The potential for CoNS to serve as reservoirs of virulence genes for other bacteria has been considered (Blaiotta et al., 2004). One of the main species of CoNS is Staphylococcus haemolyticus, which accounts for 10–20% of clinical CoNS infections (Ben Saida et al., 2009; Szczuka et al., 2015).
Foodborne illness mainly affects children, and in Argentina, more than 4.5 million children eat at school (Veleda et al., 2014). The information on the possible combination of enterotoxin genes and the mecA gene in Staphylococcus strains plus the antimicrobial sensibility profiles is useful to detect the presence of foodborne-resistant strains and to analyze the potential importance of school dining rooms as a vehicle for AMR. The aims of this study were to isolate coagulase-positive Staphylococcus (CoPS) and CoNS, particularly S. aureus, from school dining rooms located in Tandil, a district situated in the southeast of Buenos Aires Province, Argentina, and to investigate the presence of virulence genes and the AMR profiles.
Materials and Methods
Sample collection
A total of 95 samples were obtained from 10 school dining rooms from Tandil, a district located in the SE of the province of Buenos Aires, Argentina. The first stage involved a simple random sampling to choose 10 of 23 schools dining rooms. Samples were collected between August 2018 and November 2019 from handlers (Acta 111/18, CA-Tandil, Argentina) (N = 40), inert surfaces (N = 35), food (N = 10), and air (N = 10). Handlers and inert surfaces samples were aseptically collected using swabs moistened with Letheen broth (Britania, Argentina). Individual swabs were rubbed onto the hands of the food handlers during food preparation and inert surfaces such as utensils, tables, and refrigerators. Food samples were placed in sterile containers. Air samples were collected by sedimentation technique. Petri dishes with Plate Count Agar were exposed to the environment for a determined period of time. All samples were transported and refrigerated in the laboratory and processed within 24 h of sampling.
Isolation and identification of Staphylococcus spp.
Swabs were incubated in Letheen broth overnight with shaking at 37°C. Twenty-five grams of food samples was cultured in 225 mL Luria-Bertani broth with shaking (120 rpm) at 37°C for 18 h. Air samples were incubated at 37°C for 24 h. An aliquot of bacterial cultures was inoculated into Baird Parker agar supplemented with yolk and potassium tellurite (Britania, Argentina) and incubated at 37°C for 24 h. Pure culture of isolates was obtained by subculture on Brain Heart Infusion (BHI; Britania, Argentina) broth. Staphylococcus was identified by phenotyping methods based on colony characteristics, pigment production, Gram staining, catalase, coagulase and DNAse tests, sugar fermentations, and urease test (Barrow and Feltham, 2003). Stock cultures were stored at −80°C in BHI broth containing 20% (v/v) glycerol.
Molecular characterization
The DNA template was obtained by boiling bacterial colonies suspended in 100 μL of Tris EDTA buffer for 10 min, as proposed by Thiran et al. (2018) with modifications. After centrifugation at 10,000 rpm for 5 min, the supernatant was suspended in 500 μL of sterile water. A duplex polymerase chain reaction (PCR) was performed to confirm the gender and to identify S. aureus, amplifying the fragments of genes 16S ARNr and nucA, respectively (Brakstad et al., 1992; Ciftci et al., 2009). In addition, a total of eight virulence and adhesin-encoding genes, luk-PV, icaA, icaD, sea, seb, sec, sed, and see, were detected according to previous studies (Bekir et al., 2012; Lina et al., 1999; Mehrotra et al., 2000).
The genes amplified in this study that were used for the identification and characterization of Staphylococcus spp. are summarized in Table 1. S. aureus ATCC 25923 (kindly supplied by Dra. F. Buzzola, Instituto de Investigaciones en Microbiología y Parasitología Médica, Laboratorio de Patogenia Bacteriana, Universidad de Buenos Aires) and S. aureus (sea +, seb +, and sec +) (kindly supplied by Dr. Gerardo Leotta, Laboratorio de Microbiología de Alimentos, Instituto de Genética Veterinaria “Ing. Fernando N. Dulout,” Facultad de Ciencias Veterinarias, Universidad Nacional de La Plata) were used as positive controls, and double distilled water was used as negative control.
Primers Used for the Identification and Characterization of Staphylococcus spp.
Phenotypic antimicrobial susceptibility testing
The isolates were tested for susceptibility to nine antibiotics using a diffusion method according to the Clinical and Laboratory Standards Institute (CLSI, 2023) instructions (Supplementary Table S1). The antimicrobials PEN, FOX, gentamicin (GEN), erythromycin (ERY), clindamycin (CLI), levofloxacin (LVX), trimethoprim–sulfamethoxazole (SXT), and tetracycline (TET) were determined using the disk diffusion method. Minimum inhibitory concentration (MICs) of VAN were determined with an Epsilometer test (E-test) (BioMérieux, Marcy l'Étoile, France) for Gram-positive strains. Detection of β-lactamase production by the disk diffusion method (PEN zone-edge test) and inducible CLI resistance (ICR) was performed. S. aureus ATCC 25923 was used as control. The results were interpreted according to the CLSI criteria (CLSI, 2023). Staphylococcus isolates resistant to three or more antimicrobial classes were categorized as multidrug resistant (MDR) (Magiorakos et al., 2012).
Detection of AMR genes
The methicillin resistance specific gene (mecA) was amplified by PCR according to Ciftci et al. (2009) and VAN resistance gene (vanA) according to Dutka-Malen et al. (1995).
Detection of hemolytic activity
The isolates were streaked onto the blood agar plates (trypticase soy agar containing 5% (v/v) cow blood (Britania, Argentina) and incubated at 37°C for 24 h. The presence of zones of clearing around the colonies was interpreted as β-hemolysis activity (positive) and the absence as γ-hemolysis activity (negative). Greenish zones around the colonies were interpreted as α-hemolysis.
Data analysis
A clustering analysis (UPGMA [unweighted-pair group method with arithmetic mean]) was performed using the software BioNumerics version 6.6 (Applied Maths).
Results
Thirty Staphylococcus isolates were obtained from the hands of food handlers, air, food, and inert surfaces from school dining rooms (Table 2). In addition to S. aureus, several CoNS were recovered, Staphylococcus fleurettii, S. haemolyticus, Staphylococcus devriesei, Staphylococcus capitis, with three to five isolates from each one, and Staphylococcus arlettae, Staphylococcus cohnii, Staphylococcus gallinarum, Staphylococcus hominis, Staphylococcus muscae, Staphylococcus rostri, Staphylococcus simulans, with one isolate from each one. Two isolates could not be determined at specific level (Fig. 1).
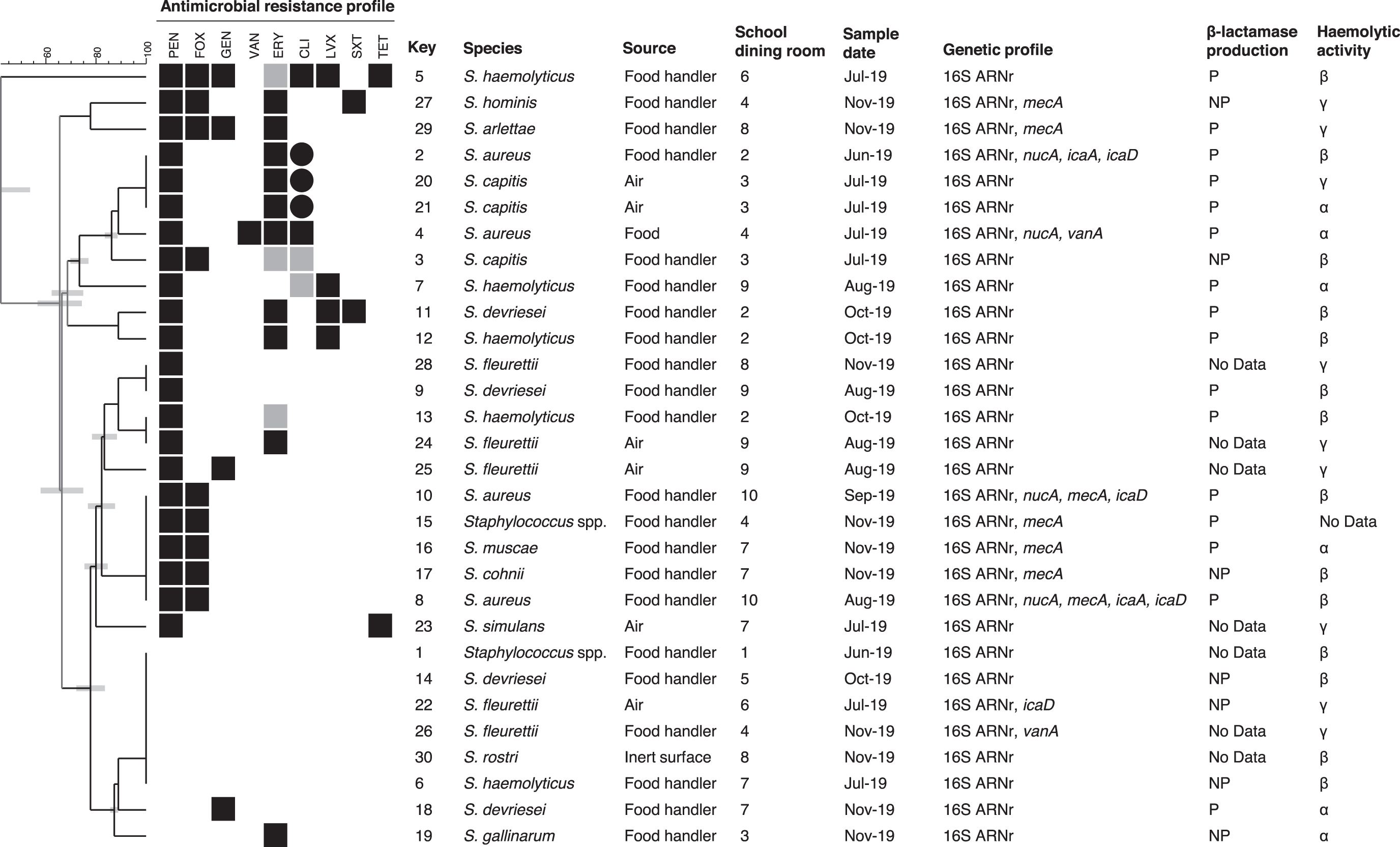
Cluster analysis of Staphylococcus spp. isolated from school dining rooms in Argentina based on antimicrobial resistance profiles. Resistance isolates for each antibiotic tested are indicated with black boxes, isolates with intermediate sensibility are indicated with gray boxes, and susceptible ones are indicated with white boxes. Besides, ICR is indicated with circles. Isolate denomination, species, source, school dining room, sample date, genetic profile, β-lactamase production, and hemolytic activity are indicated. Genes not found in any of the studied isolates: sea, seb, sec, sed, see, luk-PV. ICR, inducible clindamycin resistance; NP, non-β-lactamase producer; P, β-lactamase producer.
Source and Genetic Profiles of Staphylococcus Spp. and Staphylococcus aureus Isolates from School Dining Rooms in Argentina
N, number of isolates.
All isolates were tested for the presence of some genes encoding toxins and adhesins. None of the isolates possessed enterotoxin genes or PVL gene. Four isolates contained intercellular adhesin icaD gene (three S. aureus and one S. fleurettii) and two icaA genes (S. aureus) (Table 2).
Regarding antimicrobial sensibility, 17 profiles were detected (Fig. 1). Resistance to PEN, FOX, ERY, CLI, LVX, GEN, SXT, TET, and VAN was found (Table 3). Only 6 isolates were sensitive to all the antibiotics investigated, and 11 isolates (37%) were MDR, resistant to 3 or more antimicrobial classes. Multidrug resistance CoNS isolates were nine and belonged to five species (S. haemolyticus, S. hominis, S. arlettae, S. capitis, and S. devriesei). Two of them were, also, methicillin-resistant coagulase-negative Staphylococcus (MR-CoNS) (S. hominis and S. arlettae isolates).
Antimicrobial sensitivity of Staphylococcus spp. and Staphylococcus aureus isolates recovered from dining rooms in Argentina
Antibiotic with no described value for intermediate sensibility.
N, number of isolates.
Seven isolates harbored the mecA gene (23%), and two of the four S. aureus were MRSA (Table 2). Both MRSA isolates came from the same school, but they were isolated from two food handlers on different date. Five MR-CoNS (S. arlettae, S. cohnii, S. hominis, S. muscae, Staphylococcus spp.) were detected by PCR, and they came from three schools (Fig. 1). Only one Staphylococcus spp. isolate was resistant to VAN, but the gene vanA was detected in two isolates (Tables 2 and 3). Three inducible CLI-resistant isolates from two school dining rooms were found, and they came from food handler and air samples.
Around 50% of isolates (n = 16) presented β-hemolytic activity. In relation to β-lactamase producers, 94% of them were resistant to PEN, whereas 57% of non-β-lactamase producers were sensitive to PEN (Fig. 1).
Discussion
To the best of our knowledge, this is the first report of AMR patterns among CoNS collected from non-health care environments in Argentina. The study allowed to obtain Staphylococcus isolates, of which 13% were S. aureus, 80% were CoNS, and 2 could not be identified to species level. The CoNS isolates belonged to 11 species, with S. haemolyticus and S. fleurettii as the most prevalent ones. The first species is the second more frequently isolated CoNS, after Staphylococcus epidermidis (Szczuka et al., 2015). Previous studies have found different CoNS species colonizing food handlers, reflecting differences in the distribution of microbiota among populations of people (da Silva Sdos et al., 2015; Rall et al., 2010; Udo et al., 1999). Five isolates were MR-CoNS. In previous studies, MR-CoNS were reported (Seng et al., 2017; Xu et al., 2015).
Contamination with S. aureus by food handlers is a frequent occurrence, given the high rate of S. aureus on human skin and in mucous membrane (Castro et al., 2016; Fernandes et al., 2022). Four sampled individuals carried S. aureus on their hands, which could contribute to the cross-contamination of food. Equipment and environmental surfaces can also be sources of contamination with S. aureus (Bhatia and Zahoor, 2007).
In this study, most of the isolates resistant to FOX were tested positive for mecA (seven of nine). The finding of two mecA-negative may be attributed to the existence of mecC gene, not detected in this study. This was in agreement with the results reported previously by García-Álvarez et al. (2011).
S. aureus isolates obtained in this study did not show serious pathogenic potential since they did not have toxin genes; however, they possessed adhesion genes and were resistant to several antibiotics. S. aureus has a remarkable ability to form biofilm, which increases the resistance of the bacterium to antimicrobial therapy, and contributes to its persistence in food processing environments and increases the cross-contamination risk (Archer et al., 2011; Vazquez-Sanchez et al., 2013). Biofilm formation requires the synthesis of an exopolysaccharide called polysaccharide intercellular adhesin (PIA), dependent on the intracellular adhesion (ica) operon.
The production of PIA is one of the mechanisms of biofilm production not only in S. aureus but also in other Staphylococcus species such as S. epidermidis (Doulgeraki et al., 2016; Mirzaei et al., 2016). The ica operon comprises four genes, among them icaA and icaD (Cramton et al., 1999) are detected, here, in S. aureus and S. fleurettii isolates. The icaD-carrying S. fleurettii isolate was obtained from air, which could be explained by the formation and spread of pathogenic bacterial aerosols (Kozajda et al., 2019). Airborne transmission plays an important role in the spread of microorganisms such as CoNS, S. aureus, and MRSA in environments such as hospitals, farms, public places, and homes (Kluytmans et al., 1995; Liu et al., 2012; Shiomori et al., 2001).
Regarding Staphylococcus resistance, more than 73% of the strains were resistant to PEN and more than 33% were resistant to ERY, a macrolide antibiotic frequently used to treat S. aureus infections. S. aureus isolates with high level of PEN resistance had been detected from most of the host species (Rao et al., 2022). On the contrary, these authors observed differences between host species in TET and CLI resistance. Other results obtained in a study on S. aureus isolated from foods of a Chinese school canteen exhibited lesser PEN resistance (59%) but similar levels to those obtained in the present study for ERY (41%) (Wang et al., 2020). The authors found an isolate resistant to VAN. The emergence of VRSA is a growing problem, and the widespread use of VAN makes resistance to the drug, a significant human health concern (Mahros et al., 2021).
In the present work, two strains that are vanA-positive were detected and, one of these, an isolate of S. aureus obtained from food, was phenotypically resistant to VAN. Gene vanA-carrying enterococci and staphylococci but VAN-susceptible in traditional phenotypic susceptibility tests that could revert to a VAN-resistant phenotype upon VAN treatment were previously reported (Li et al., 2022). SXT resistance has been demonstrated in S. aureus isolates from dairy cattle and milk products (Abdeen et al. 2020; Rao et al., 2022). In this study, resistant isolates to this combined antibiotic came from food handlers and belonged to S. devriesei and S. hominis.
This study reports the presence of the methicillin-resistant staphylococci within non-health care environments. Food handlers carried high levels of MRSA strains on their hands, which represents an increased risk of MRSA strains entering the food chain, and this should be a cause for concern. On the contrary, Staphylococcus non-aureus species might act as a reservoir of antibiotic resistance and potentially transfer it to MRSA. In this study, two methicillin-resistant Staphylococcus spp. (MRSS) isolates, S. hominis and S. arlettae, were MDR (PEN-FOX-ERY + SXT or GEN).
School feeding plays a key role in the nutrition of children from vulnerable sectors, and the consumption of food contaminated with MRSA can be a serious threat to health. Although this study was carried out only in a district of Argentina, the results are valuable since they show the importance of school dining rooms as a vehicle for AMR and provide the basis for future comparisons. In particular, it was detected that the handlers were the source of MRSA, VRSA, MR-CoNS, and MDR isolates. The results obtained indicate that the vigilance of these pathogens in school dining rooms should be extreme.
Footnotes
Acknowledgments
The authors thank all the staff of the educational institutions sampled in this study and Maria Rosa Ortiz for her technical assistance.
Authors' Contributions
J.G.: Conceptualization, methodology, investigation, writing—original draft, project administration, funding acquisition. L.H.: Validation, investigation. A.T.: Methodology, validation, resources. A.V.B.: Conceptualization, writing—review and editing. A.M.S.: Resources, conceptualization, supervision, project administration, funding acquisition.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grant from Secretaría de Ciencia, Arte y Tecnología-Universidad Nacional del Centro de la Provincia de Buenos Aires (SECAT-UNCPBA-03-JOVIN-48H).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.