
Abstract
Toxoplasma gondii, an important food-borne zoonotic parasite, poses a worldwide public health hazard. Domestic pigs are considered one of the main intermediate hosts in the zoonotic transmission of T. gondii. To date, seroepidemiological information on T. gondii in domestic pigs in India is very scarce, and there are no reports of occupational hazards to pig farmers in this country. Here, we aimed at estimating the occurrence of T. gondii (antibodies and parasite DNA) in slaughtered pigs and pig farmers in Central India. Seroprevalence was determined in 410 serum samples from slaughtered pigs and 103 sera from pig farmers using an in-house prepared antigen-based modified agglutination test (MAT), enzyme-linked immunosorbent assay (ELISA), and indirect-fluorescent antibody test (IFAT). Anti-T. gondii IgG antibodies were detected in 200 pigs (up to 48.8%, confidence interval [95% CI]: 40.4–52.2) and 44 pig farmers (up to 42.7%, 95% CI: 35.6–47.3) using MAT, ELISA, and IFAT. Inter-rater agreement showed an excellent agreement (kappa κ = 0.9) among the different serological tests suggesting similar detection potential of these tests. Recently acquired infections in all seropositive subjects were determined using IgG avidity testing and polymerase chain reaction (PCR). IgG avidity showed that 20 (10.3%) of slaughtered pigs and 8 (19.5%) pig farmers had a recently acquired infection. PCR for B1 and 529 repeats was performed in the heart tissues of slaughtered pigs and the blood cells of pig farmers. T. gondii DNA was detected in 14 (7.2%) slaughtered pigs and 5 (12.2%) pig farmers. Univariate analysis revealed that adult animals (>1 year), cats and rodents on the farm, and outdoor access are common factors (p ≤ 0.05) associated with T. gondii infection in pigs. Our results indicate that T. gondii is widely distributed in slaughtered pigs and pig farmers at risk of infection, highlighting a potential zoonotic transmission and health risk to consumers.
Introduction
Toxoplasmosis is one of the most common foodborne zoonoses worldwide caused by the protozoan parasite Toxoplasma gondii, causing abortion and neonatal mortality in humans and animals (Dubey, 2021). Toxoplasmosis has been ranked fourth among foodborne parasitic infections of global concern (FAO/WHO report, 2014; Torgerson et al., 2015).
According to the European Food Safety Authority (EFSA), T. gondii is a potential biological hazard that cannot be detected using meat inspection procedures (EFSA, 2014). Pigs are intermediate hosts of T. gondii, and pork represents a large proportion of the meat consumed worldwide. In Europe and the United States, ingesting infected raw or undercooked meat is a major source of human T. gondii infection (Kijlstra and Jongert, 2008).
The global pooled seroprevalence of T. gondii in pigs was estimated at 19% (reviewed by Foroutan et al., 2019). Serological tests such as indirect-fluorescent antibody test (IFAT), modified agglutination test (MAT), and enzyme-linked immunosorbent assay (ELISA) showed the highest sensitivity and specificity for detecting T. gondii infection (Hill and Dubey, 2002).
However, combining serology and molecular methods may better assess the risk of T. gondii infection from infected animals (Liu et al., 2015). The serological screening of pigs can be beneficial for assessing infection risk in meat due to a significant relationship between the T. gondii seropositivity and the presence of live parasites in their tissues (Sroka et al., 2020).
Around 0.1 million children in India are born with a possible risk of congenital toxoplasmosis yearly (Singh, 2016). The nationwide survey on T. gondii infection indicated 24% and 22% seroprevalence in humans (Dhumne et al., 2007) and pregnant women (Singh et al., 2014). The regional studies showed that veterinary personnel and abattoir workers are at risk of T. gondii infection (Deshmukh et al., 2021a; Thakur et al., 2022).
The pig is one of the economically important livestock in India, and as per the 20th livestock census, there are 9.06 million pigs in India (Animal Husbandry Annual Report, 2021). Despite global importance, surprisingly, only one regional study was conducted in the past four decades and reported 48% seroprevalence in slaughtered pigs (Thakur et al., 2019).
The lack of information regarding the prevalence and factors associated with T. gondii infection in pigs highlights important data and knowledge gaps for assessing the risk of pork to human infection. This study aimed at estimating the prevalence of T. gondii in slaughtered pigs and pig farmers in central India and at identifying factors that may favor infection.
Materials and Methods
Study design and data collection
The institutional ethics committees (National Institute of Animal Biotechnology—IAEC/NIAB/2019/48/ASD; Nagpur Veterinary College—NVC/2019/SC) have approved using animal and human samples and research protocols. Samples were collected (March–December, 2022) from Maharashtra (19.66°N, 75.3°E) and Telangana (18°N, 79°E) regions of Central India (Fig. 1). The sample size was calculated (Lwanga and Lemeshow, 1991) using the formula n ≥ Z 2. p (1 − p)/d 2, where n = sample size, z = level of confidence, p = estimated proportion of the population, d = margin of error.

Regions of Central India showing the locations of pig slaughterhouses and pig farmers sampled in the study.
The estimated sample size for slaughtered pigs was 385 at 95% of confidence, 5% of margin of error, and 50% of assumed prevalence (Thakur et al., 2019). Accordingly, 410 samples of pigs (blood and heart tissue) were collected using the random sampling method. The sampling was done multiple times to ensure that pigs belonged to different batches.
Data regarding the farms of origin, animal details, and rearing conditions were obtained from pig farmers who came to slaughterhouses through a questionnaire and a short interview. Samples were collected over 6 months, during which there were no marked changes in temperature, humidity, or precipitation patterns. Data on age, sex, presence of cats, rodents, other farm animals, and outdoor access for individual pig slaughtered were also recorded.
Blood samples of 103 pig farmers (males) who regularly supply pigs to these slaughterhouses and consume pork routinely were screened. Serum was used for antibody testing. Peripheral blood mononuclear cells (PBMCs) were collected from heparinized blood by a density gradient centrifugation method using Ficoll Histopaque (Histopaque-10771; Sigma) and used for detecting parasite DNA by polymerase chain reaction (PCR).
Digestion of tissue samples and DNA extraction
Pig heart tissues were digested with pepsin solution as previously described (Dubey, 2021). DNA from the recovered pellets and PBMCs was extracted using QIAmp DNA Mini Kit and stored at −20°C until examination.
Parasite culture
T. gondii parasites were maintained in human foreskin fibroblasts (HFF No. CRL-1634; ATCC) cells in DMEM supplemented with 10% fetal bovine serum, 10 μg/mL Gentamicin, 1% Penicillin/Streptomycin, and 2 mM
IgG ELISA
IgG-ELISA was performed using T. gondii lysate antigen (TLA) as described previously (Hebbar et al., 2022a). Before testing the test sera, TLA was standardized for purity, concentration, and incubation time. The standardization results revealed that TLA is pure, requires 1 μg/mL concentration for coating the wells, and gives >0.7 OD value for positive and <0.2 OD value for negative samples. The control and test sera were used in duplicates at 1:100 and 1:200 dilutions.
Bound IgG was detected using anti-pig IgG conjugated with horseradish peroxidase (HRP) at 1:5000 dilution (No. AP166P; Sigma) or anti-human IgG conjugated with HRP at 1:5000 dilution (No. A18805; Invitrogen). Cut-off values were calculated as the mean OD plus 2 standard deviations values obtained with negative samples.
Modified agglutination test
MAT was performed as described previously (Hebbar et al., 2022a). The test was performed using a serum dilution range from 1:25 to 1:200. The result was considered positive when a layer of agglutinated parasites was observed at serum dilutions of 1:25 or higher. The control sera (positive and negative) were included in each plate.
Indirect fluorescent antibody test
IFAT was performed as described previously (Deshmukh et al., 2021b). Serum samples were tested using 1:100 and 1:200 dilutions. FITC-conjugated anti-pig (No. SAB4600047; Sigma) or anti-human (No. 31529; Invitrogen) antibodies were used at 1:1000 dilution. The tachyzoite to bradyzoite stage differentiation was carried out by incubating the parasites in bradyzoite induction medium (RPMI pH 8.2) at 37°C for 5 days without CO2.
Coverslips were mounted with Vectashield medium (No. H-1200-10; Vector Laboratories) with DAPI on a glass slide. T. gondii morphology marker inner membrane complex 1 (IMC1) (Mitra et al., 2019) and cyst wall protein 1 (CST1) antibodies (Deshmukh et al., 2021b) were used to stain the tachyzoites and bradyzoites, respectively. The images were captured using a confocal microscope (Leica) with a 100 × objective and processed using LAS X software.
IgG avidity ELISA (IgG avidity-ELISA)
The avidity test was performed using ELISA (Hebbar et al., 2022b). Results were represented as the avidity indices (AI). AI (%) = (OD of washed urea well/OD of washed regular buffer well) × 100. Cut-off values were calculated for each dilution as the mean OD plus 2 SD values of the negative samples, with or without urea treatments. The AIs of <45%, 45% to 55%, and >55% were considered low avidity, equivocal avidity, and high avidity, respectively.
Polymerase chain reaction
For PCR amplification, 2 targets, B1 gene (Burg et al., 1989), and 529 repeats (Homan et al., 2000) were used. The primers used to amplify B1 gene (651 bp) were TCGCAGTACACCAGGAGTTG and CACTCCATCTCTCGTCTTCT, and 529-repeats (182 bp) were TGTGCTTGGAGCCACAGAAG and GCAGCCAAGCCGGAAACAT, respectively.
The following PCR cycle conditions were used (Rahumatullah et al., 2012): 95°C for 5 min followed by 35 cycles of 95°C/10 s, 57°C/10 s, 72°C/30 s, and a final extension step at 72°C/2 min. Each assay included positive (T. gondii genomic DNA) and negative (ultrapure H2O) controls. PCRs were carried out in a 20 μL reaction volume containing 200 ng of extracted DNA, 10 pmol of each primer, 0.2 mM dNTP, 1.5 mM MgCl2, and 1.5 U Taq DNA polymerase (No. M0267; NEB, United Kingdom).
Statistical analysis
The data from serological tests and univariate analysis were analyzed using GraphPad Prism and SPSS software, respectively. The kappa coefficient (κ) was calculated to assess agreements between various serological diagnostic assays. Values of p < 0.05 were considered statistically significant for univariate analysis.
Results
Seroprevalence of T. gondii infection in slaughtered pigs and pig farmers
The serum samples of 410 slaughtered pigs and 103 pig farmers were evaluated using ELISA (dilution: 1:100/200), MAT (dilution: 1:25–200), and IFAT (dilution: 1:100/200). IgG-ELISA results revealed that 47.3% (194/410) of slaughtered pigs and 39.8% (41/103) of pig farmers were seropositive for T. gondii (Fig. 2A and Table 1) for both the serum dilutions.

Seroprevalence of Toxoplasma gondii infection in slaughtered pigs and pig farmers.
Detection of Antibodies to Toxoplasma gondii in Slaughtered Pigs and Pig Farmers Using Enzyme-Linked Immunosorbent Assay, Modified Agglutination Test, and Indirect-Fluorescent Antibody Test
+, positive; Bz, bradyzoite; ELISA, enzyme-linked immunosorbent assay; IFAT, indirect-fluorescent antibody test; MAT, modified agglutination test; Tz, tachyzoite.
MAT results showed that 48.3% (198/410) of slaughtered pigs and 39.8% (41/103) of pig farmers were seropositive (Fig. 2B and Table 1). Pig MAT titers were 25 in 41 (20.7%), 50 in 40 (23.9%), 100 in 50 (25.7%), and 200 in 67 (47.3%). Pig farmers MAT titers were 25 in 1 (2.5%), 50 in 5 (10.5%), 100 in 10 (25%), and 200 in 25 (62.5%).
The IFAT results demonstrated that sera from 48.8% (200/410) of slaughtered pigs and 42.7% (44/103) of pig farmers were able to detect intracellular tachyzoite (Tz) of T. gondii (Fig. 2C, D and Table 2). All seropositive samples were also tested against in vitro-induced bradyzoites (Bz). In the case of slaughter pig samples, 99% of Tz-IFAT samples were able to detect bradyzoites (Fig. 2E), whereas 97.7% of Tz-IFAT samples of pig farmers recognized bradyzoites (Fig. 2F).
Results of Toxoplasma gondii Detection Using Modified Agglutination Test and Enzyme-Linked Immunosorbent Assay, Compared with Indirect-Fluorescent Antibody Test Results
ELISA, enzyme-linked immunosorbent assay; IFAT, indirect-fluorescent antibody test.
The seropositive sera of T. gondii almost recognized the surface and internal structures of both tachyzoites and bradyzoites. The seronegative samples showed no staining of intracellular tachyzoites (Fig. 2C, D) and bradyzoites (Fig. 2E, F). Identical results were observed for both the dilutions of serum (1:100 and 1:200).
In this study, IFAT was slightly better than MAT and ELISA in diagnosing the T. gondii antibodies. Hence, we compared MAT, and ELISA results with IFAT. Compared with IFAT, MAT and ELISA showed slightly lower sensitivity and specificity (Table 2). However, overall, excellent agreements were obtained between ELISA and MAT with IFAT (kappa value [κ] > 0.88).
Many seropositive pigs and pig farmers had a recently acquired infection
Testing of IgG avidity level and parasite DNA help detect recently acquired infection. Of the 194 seropositive pigs, 108 (55.7%) had high IgG avidity (>55%), 20 (10.3%) had low IgG avidity (<45%), and 66 (34%) had equivocal avidity (45% to 55%) (Fig. 3A, B and Table 3). Of the 41 seropositive pig farmers, 11 (26.8%) had high IgG avidity, 8 (19.5%) had low IgG avidity (<45%), and 22 (53.7%) had equivocal avidity (45% to 55%) (Fig. 3A, B and Table 3).
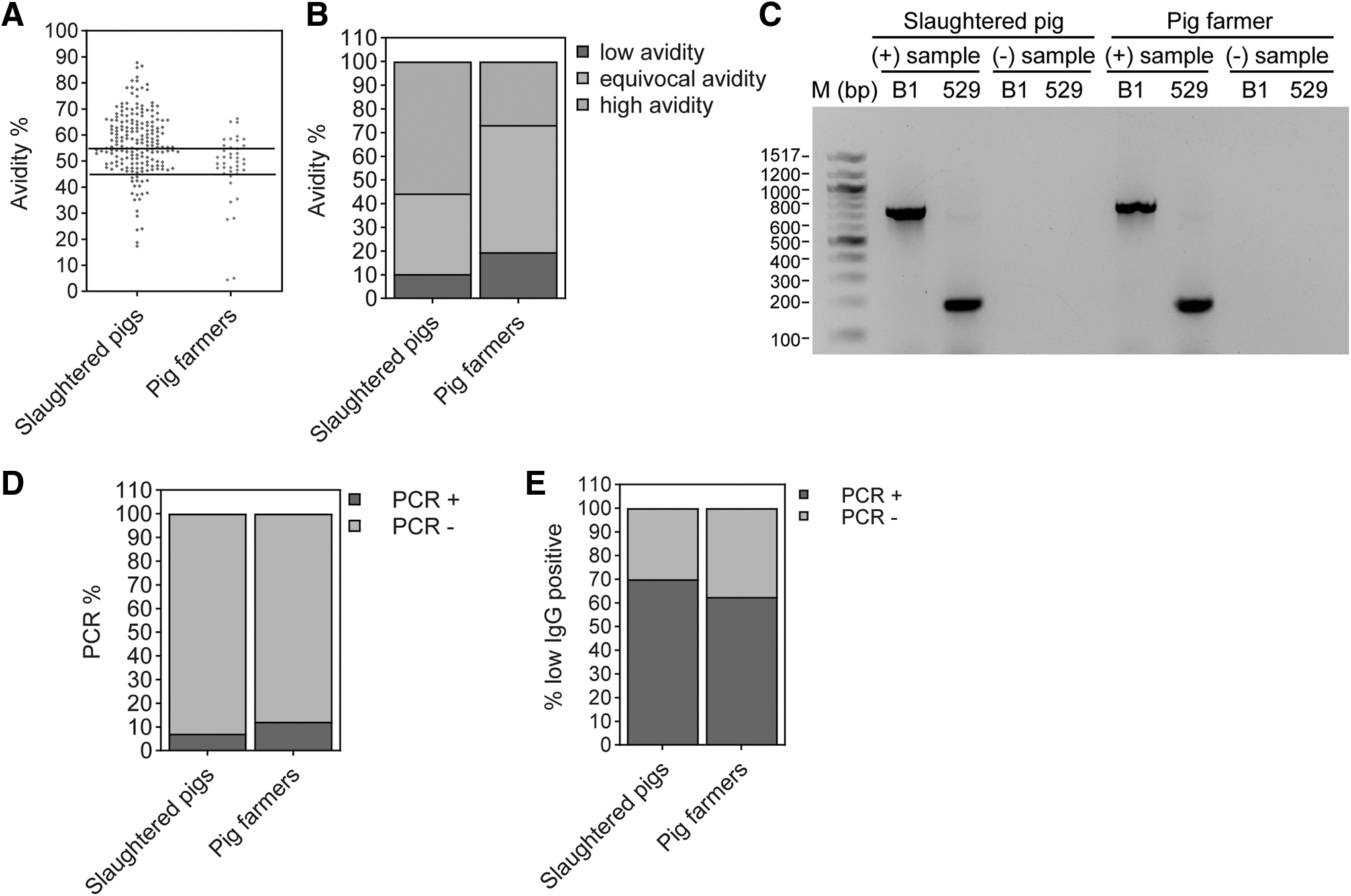
Determination of recently acquired infection in slaughtered pigs and pig farmers.
Detection of Recently Acquired Infection in Toxoplasma gondii Seropositive Pigs and Pig Farmers Using IgG Avidity and Polymerase Chain Reaction
PCR, polymerase chain reaction.
All seropositive subjects were tested for the presence of parasite DNA using PCR. Of the 194 seropositive pigs, 14 (7.2%) pigs had parasite DNA in their heart tissues, and 5 (12.2%) of 41 seropositive pig farmers had parasite DNA in their PBMCs (Fig. 3C, D). In comparison with IgG avidity, of the 20 T. gondii seropositive low IgG avidity slaughtered pigs, 14 (70%) were positive for T. gondii DNA (Fig. 3E and Table 3) and 62.5% (5 of 8) of T. gondii seropositive low IgG avidity pig farmers were positive for T. gondii DNA (Fig. 3E and Table 3).
In comparison with low IgG avidity results, 14 (70%) of 20 pigs and 5 (62.5%) of 8 pig farmers were positive for T. gondii DNA (Fig. 3E and Table 3).
Identification of factors associated with T. gondii infection in slaughtered pigs
The variables from the univariate analysis (Table 4) significantly associated with T. gondii seropositivity (p < 0.05) were: slaughtered pigs that were >1 year old (odds ratio [OR]: 2.2, confidence interval [95% CI] 1.47–2.32), presence of cats in the farm (OR: 2.85, 95% CI 1.9–4.28), presence of rodents in the farm and feed storage area (OR: 2.67, 95% CI 1.79–3.98), and outdoor access (outside farm boundaries) (OR: 2.25, 95% CI 1.46–3.46). However, the sex of the slaughtered pigs and the presence of other livestock species in the farm area did not significantly contribute to T. gondii infection in these pigs.
Univariate Analysis of Risk Factors Associated with Seropositivity of Toxoplasma gondii in Slaughtered Pigs (n = 410)
CI, confidence interval; IFAT, indirect-fluorescent antibody test; OR, odds ratio.
Discussion
This is the first study from Central India demonstrating the T. gondii seroprevalence in slaughtered pigs and pig farmers and analyzing the factors associated with infection. MAT, ELISA, and IFAT results showed that overall seroprevalence in slaughtered pigs and pig farmers was up to 48.8% and up to 42.7%, respectively. Recently acquired infection was noticed in 10.3% of slaughtered pigs and 19.5% of pig farmers; however, parasite DNA could be detected in 7.2% of slaughtered pigs and 12.2% of pig farmers.
The T. gondii seroprevalence of pigs observed in our study is more than the global pooled pig seroprevalence, 19% (reviewed in Foroutan et al., 2019). The highest pooled seroprevalence was observed in Africa (25%), followed by North America (25%), South America (23%), Asia (21%), and Australia (2%).
The seroprevalence data reported from different continents/countries may differ due to differences in the geographical locations, methods employed for testing, and production systems. However, the overall seroprevalence in slaughtered pigs and recently acquired infection was comparable with the earlier study from north India (Thakur et al., 2019).
In the IFAT, we used IMC1 and CST1 antibodies to stain the tachyzoites and bradyzoites, respectively. The IMC is a specialized structure comprising >25 IMC proteins localized beneath the parasite's plasma membrane that provide support at the periphery of the parasite (Hu et al., 2002). Antibodies against IMC proteins, including IMC1, are used as morphological markers to stain the tachyzoite stage. However, it is not a tachyzoite stage-specific marker.
During the differentiation of tachyzoite to bradyzoite stage, the parasitophorous vacuole membrane of tachyzoite transforms into a cyst wall. The cyst wall comprises several proteins, including major CST1. Staining with CST1 antibody confirms the bradyzoite stage of the parasite. The diagnostic (Deshmukh et al., 2021a) and vaccine (Eom et al., 2023) utilities of CST1 have been reported.
The IFAT is the gold-standard test for T. gondii detection in different livestock species, including pigs (Garcia et al., 2006; Lind et al., 1997); hence we adopted the IFAT as a reference test to compare its diagnostic performance with MAT and ELISA. We observed good agreements between these tests, and results aligned with the calculated sensitivity and specificity.
Given that the IFAT is slightly better at detecting antibodies to T. gondii than MAT and ELISA (Garcia et al., 2006; Hebbar et al., 2022b), we suggest that the IFAT should remain the assay of choice in the seroprevalence study. However, several studies showed that MAT and ELISA are reliable tests for T. gondii detection in pigs (Dubey et al., 1995; Gamble et al., 2005; Garcia et al., 2006).
MAT is used as a serological reference test because it showed high agreement with bioassays in mice and cats used to detect viable T. gondii in pigs (Forbes et al., 2012; Opsteegh et al., 2016). Similarly, Gamble et al. (2005) and Hill et al. (2006) obtained a higher agreement of bioassays in cats using an ELISA to detect T. gondii antibodies in pigs.
Also, very few differences were observed in identifying the number of positive samples of pigs and pig farmers using the tachyzoite and bradyzoite IFAT, indicating a robust humoral response against bradyzoite antigens. Several attempts have been made to identify the diagnostic and protective antigens from T. gondii in recent years. The IFAT results showed a strong antibody response against antigens expressed in the tachyzoite and bradyzoite stages, suggesting the tremendous potential of these antigens to be utilized in the development of diagnostic assays and vaccines.
The immunodomaint proteins expressed in the bradyzoite stage deserve more attention because bradyzoites have long-term latent infection ability. Moreover, antibodies against bradyzoite antigens can be used to distinguish between acute and chronic infection.
We observed little discrepancy between IgG avidity and PCR results. Parasite DNA was observed in 70% (14 of 20) and 62.5% (5 of 8) of low IgG avidity slaughtered pigs and pig farmers, respectively, which could be attributed to limited tissue sample size, small number of parasites in the original material, and random distribution of tissue cysts. T. gondii DNA detection efficiency can increase if the parasites in tissue samples are multiplied by bioassay or cell culture before DNA analysis, which was not possible in this study.
Interpretation of IgG avidity results is complicated, as persistent low avidity results have been reported in serum from some patients with past infections for months. The results of gray-zone avidity should be interpreted cautiously unless antibody titers have perfect stability in two sera taken 2–3 weeks apart.
In this study, we observed (1) older animals (>1 year), (2) the presence of cats, (3) rodents on the farm, and (4) access to outdoor as potential factors associated with T. gondii infection in pigs. (1) The prevalence of T. gondii increases with the age, explained by the maximum persistence of tissue cysts in pigs and the probability of pigs getting into contact with the parasite increases with age (Castillo-Cuenca et al., 2021; Hill et al., 2010; Rostami et al., 2017). (2) The movement of cats on the farm poses a risk of acquiring T. gondii infection as one infected cat can shed millions of oocysts and contaminate a wide field area quickly.
After primary infection, the oocyst excretion occurs mainly in seronegative cats, followed by a long-lasting humoral immune response against the parasite. However, oocyst excretion does not occur in all cats that come into contact with infective forms of parasite regardless of their T. gondii infection status. Similar results were reported previously (Meerburg et al., 2006; Papatsiros et al., 2016; Weigel et al., 1995). (3) Rodents play an essential role in the life cycle and epidemiology of T. gondii because they are considered the primary source of infection for domestic cats (Garcia-Bocanegra et al., 2010; Hill et al., 2006; Kijlstra et al., 2008; Papatsiros et al., 2016). (4) The outdoor access and scavenging leads increase the chance for pigs to contact contaminated soil, pasture, and water with T. gondii infective stages (Limon et al., 2017; Pablos-Tanarro et al., 2018; Wallander et al., 2016).
The gender (Hill et al., 2014; Roqueplo et al., 2017) and presence of other farm animals (cattle, sheep, and goat) (Djokic et al., 2016; Limon et al., 2017) were not significantly associated with seropositivity (p = 0.52), which corroborates the results of previous studies.
The study showed that 42.7% of pig farmers are seropositive, and 12.2% carry T. gondii infection, indicating possible occupational hazards. The relation between antibodies to T. gondii in pig farmers and pigs is not necessarily causal because if the soil and environment of the farm are contaminated with oocysts, the farmer is exposed to oocysts in the dust during work in the piggery. In the studied area, tribal communities rear pigs and prefer pork to other meats.
Generally, there is no raw pork consumption habit in these communities, but there are possibilities of unsafe handling and preparation of pork leading to undercooked smoked pork. In India, pigs are mostly reared in poor hygienic conditions and have free access to garbage and stray animals or rodents (Chauhan et al., 2016). Therefore, pig farmers should be educated about good hygienic practices to raise healthy and disease-free pigs, which will help prevent food-borne zoonoses and economic losses.
Conclusion
This is India's first T. gondii prevalence concurrent report of slaughtered pigs and pig farmers, indicating potential occupational hazards. Serological surveillance seems a valuable tool to guide and monitor the control of T. gondii in pork production due to the inability to detect T. gondii tissue cysts during routine post-martem examinations at slaughterhouses.
Pig farmers should be advised about the potential danger of having cats and rodents on the farm and allowing pigs outside the farm area. This study will contribute to developing improved control strategies against T. gondii in this country. Further molecular studies on genotypes circulating in pig farms must be conducted to increase the knowledge of the epidemiology of T. gondii in India.
Footnotes
Authors' Contributions
M.R.: Data curation, Investigation, Analysis, and Writing manuscript. V.M.: Data curation. P.M.: Investigation, Funding, and Writing manuscript. Y.U., V.S., and W.K.: Resources and investigation. A.S.D.: Conceptualization, Funding acquisition, Investigation, and Writing manuscript.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the DBT (Grant no. BT/PR40763/ADV/90/289/2020 and Grant no. BT/PR41294/MED/29/1545/2020) to A.S.D. The authors acknowledge the facilities and the technical assistance of the NIAB microscopy imaging center.