
Abstract
Salmonella enterica serovar Typhimurium and its variants are the most common serotypes of human salmonellosis cases. Serotyping Salmonella Typhimurium and its variants has always been challenging. Our previous work found that among 14 Salmonella Typhimurium and variant strains, some different antigenic formulas had 100% pulsed-field gel electrophoresis (PFGE) similarity. The 14 strains were sorted into 3 groups; in each group, the different antigenic formulas had the same PFGE patterns. This phenomenon suggested that different antigenic formula identification might originate from a common ancestor subtyped by PFGE. To assess whether the serotyping method on Salmonella Typhimurium and variant strains reflected the genetic relationship, we improved the discrimination for the phylogenetic relationship among the 14 Salmonella Typhimurium and variant strains using Fourier-transform infrared spectroscopy (FTIR) and whole-genome multilocus sequence typing (wgMLST). We compared the wgMLST assay of 14 Salmonella Typhimurium and variant strains from this study with 50 public ST34 strain data of Salmonella Typhimurium and variant strains. We also compared flagella (H antigen)-related genes based on the whole genome of 14 strains and the other 293 ST34 public database for further understanding of this question. The phylogenetic results (PFGE) showed no regularity between the antigenic formulas and genotypes. The results of the higher discrimination power assays (FTIR and whole-genome multilocus sequence typing) also showed no regularity between the antigenic formulas and genotypes (or phenotypes). The 58 flagella encoding genes of different antigenic formulas were sorted into 13 patterns. However, a similar phenomenon was found: the same flagella encoding gene patterns could express different antigenic formulas. In conclusion, there is no consistency between the antigenic formulas and phylogenetic relationships among ST34 Salmonella Typhimurium and variant strains, even in flagella antigenic formula and flagella encoding genes.
Introduction
Salmonella spp. poses a serious threat to public health worldwide. According to a 2023 report by the World Health Organization (WHO), ∼1 in 10 people worldwide suffer from Salmonella-related illness each year, resulting in a loss of 33 million years of healthy life (WHO, 2023). A meta-analysis also found that the average annual incidence (95% confidence interval) of nontyphoidal Salmonella invasive disease reached 44.8 (31.5–60.5) cases per 100,000 people (Marchello et al., 2021). Most Salmonella infections in humans are caused by a limited number of serotypes, which are of significant public health concern.
The top five common serotypes reported by European Food Safety Authority (EFSA) are as follows: Salmonella serotype Enteritidis, Salmonella serotype Infantis, Salmonella serotype Typhimurium, monophasic Salmonella Typhimurium (1,4,[5],12:i:-), and Salmonella serotype Derby (EFSA and ECDC, 2022). Among them, Salmonella Typhimurium and its variants are the most common, accounting for 20.2% of human salmonellosis cases in the European Union in 2021 (EFSA and ECDC, 2022).
Salmonella Typhimurium and its monophasic variants have been a long-standing public health concern. Since the last decades of the 20th century, many countries have reported the emergence of the Salmonella Typhimurium monophasic variants (Switt et al., 2009), which were officially recognized in 2003 (CDC, 2005). Since then, the Salmonella Typhimurium monophasic variants have increased steadily year after year and reached the top three serotypes in humans in the European Union (EFSA and ECDC, 2022; EFSA, 2018; EFSA, 2007).
In the United States, the Salmonella Typhimurium monophasic variants have been ranked NO. 5 since 2006 until now (Collins et al, 2022; CDC, 2017; CDC, 2011a; CDC, 2011b; CDC, 2009). Meanwhile, the Salmonella Typhimurium serotype, which was the NO. 1 serotype in humans from 1970 to 2006 in the United States (CDC, 2008), had been declining during this 2006–2021 period (CDC, 2017; Collins et al., 2022), while it has been ranked NO. 2 in humans in the European Union since 2004 until now (EFSA and ECDC, 2022; EFSA, 2018; EFSA, 2005).
For over eight decades, serotyping of Salmonella has been a very useful subtyping method for studying epidemics, but the serotyping report work of Salmonella Typhimurium and its variants has always been challenging. The variants of Salmonella Typhimurium with flagellar mutations can be divided into two groups of monophasic variants and one group of nonmotile variant: the monophasic variants have either the first or the second flagellar phase missing, and the nonmotile variant has both flagellar phases missing. However, EFSA only categorized Salmonella Typhimurium monophasic variants that lacked the second flagellar phase, such as Salmonella 1,4,[5],12:i:-, Salmonella 1,4,5,12:i:-, Salmonella 1,4,12:i:-, Salmonella 4,[5],12:i:-, Salmonella 4,5,12:i:-, and Salmonella 4,12:i:- (EFSA, 2018). This was done to reduce the bias in assessing the distribution of Salmonella serotypes that cause human infections.
For serotyping, EFSA suggested confirmation by PCR based on the gene fliA–fliB detection. However, this method would fail to detect the Salmonella Typhimurium and its variants that have IS200 fragment of 1-kb in the fliB-fliA intergenic region (EFSA, 2010). Bugarel et al. (2012) improved this protocol by developing a seven-marker PCR method, yet this PCR method also lacked good discriminatory power among these strains and was time-consuming.
Based on a deep molecular study on 14 Salmonella Typhimurium and variant strains from previous isolates (Xin et al., 2020), we found that some different antigenic formulas shared 100% pulsed-field gel electrophoresis (PFGE) similarity: the H antigens of phase one “i” and of phase two “1,2” could be lost, but they had 100% PFGE similarity. This reveals that possibly there is no consistent correlation between the specific antigenic formulas and the evolutionary relationships of these strains.
To assess whether the serotyping method on Salmonella Typhimurium and variant strains reflected the genetic relationship, we improved the discrimination for the phylogenetic relationship (using Fourier-transform infrared spectroscopy [FTIR] and whole-genome multilocus sequence typing [wgMLST]) among the 14 Salmonella Typhimurium and variant strains. The 14 Salmonella Typhimurium and variant strains were subtyped as ST34 by MLST typing. Then, we compared the flagella (H antigen) encoding genes based on the whole genome of 14 strains (this study) and the other 293 ST34 strains (downloaded from NCBI public database) of Salmonella Typhimurium and variant strains for further understanding of this question. The results of this work would provide more information for the serotype identification of Salmonella Typhimurium and its variants.
Materials and Methods
Materials
The 14 Salmonella Typhimurium and variant strains isolated from pig rectal and pork samples in slaughterhouse in Khon Kaen Thailand during 2017–2018 (Xin et al., 2020) were selected to conduct a further molecular study. We then compared the whole genomes of these strains with the same MLST subtype Salmonella Typhimurium and variant strains from the 1785 published whole genomes in NCBI database (Supplementary Data S1).
Serotyping
Serotyping of 14 Salmonella Typhimurium and variant strains had been identified by WHO National Salmonella and Shigella Center (Xin et al., 2020), assigned according to the Kauffmann–White scheme (Grimont and Weill, 2007).
PFGE analysis
The subtyping of 14 isolates was performed following the CDC standard (CDC, 2017). The DNA fragments were digested by XbaI and separated by a CHEF-DRⅢ Pulsed-Field Electrophoresis System. The gels were stained with ethidium bromide and documented by ChemiDocTM XRS+ (Bio-Rad). The PFGE electrophoresis map was labeled with the positions of each band and input into the BioNumerics software (version 7.6.3). This software conducts cluster analysis and calculates genetic similarity by analyzing the distance between each marker band. The dendrogram was produced using band clustering with the dice coefficient similarity index of 1% optimization and 1% tolerance by the unweighted-pair group method with arithmetic means (UPGMA).
FTIR spectroscopy analysis
Fourteen Salmonella Typhimurium and variant strains were prepared by incubating, washing, and resuspending onto BaF2 windows for FTIR microspectroscopy. The infrared spectra were collected using Bruker IR spectrometer (Tensor 27) coupled to IR microscope (Hyperion 2000) with 36 × magnification at beamline 4.1 Infrared spectroscopy and imaging, Synchrotron Light Research Institute (Public Organization), Thailand. The spectra were measured between wavenumbers 4000 and 400 cm−1.
The data were obtained from 64 scans at a resolution of 4 cm−1. Then, all spectra of each sample were averaged into one spectrum, and transformed by first derivative and vector normalization in the range from 3800 to 1000 cm−1. The spectrum was processed using OPUS 7.5 software (Bruker Optics); each sample was transformed by applying water compensation. The hierarchical cluster analysis (HCA) and principal component analysis (PCA) were analyzed by software Unscrambler × 10.5.
Whole-genome preparation
The whole genome of 14 Salmonella Typhimurium and variant strains was sequenced by Illumina system with 10 Gbp clean data from each sample. The genome sequences were denovo-assembled by SPAdes (v3.15.4) from clean data. The average sequencing Q30 score reached 95.27%. The 1785 published Salmonella Typhimurium and variant strains were downloaded from NCBI database (accessed in March 14, 2022); information of 1785 strains is listed in Supplementary Data S1.
wgMLST analysis
All the genome data were typed by PubMLST (https://pubmlst.org/), according to the multilocus: aroC, dnaN, hemD, hisD, purE, sucA, and thrA. The aroC type 10, dnaN type 19, hemD type 12, hisD type 9, purE type 5, sucA type 9, and thrA type 2 were recognized as ST34 according to the PubMLST website database. The 14 Salmonella Typhimurium and variant strains were typed as ST34 (shown in Supplementary Data S1). Subsequently, the wgMLST of 14 Salmonella Typhimurium and variant strains with the related ST34 Salmonella Typhimurium and variant strains were analyzed by PubMLST.
The strains include 50 related ST34 strains and one reference strain (Salmonella Typhimurium str. LT2, Assembly number: GCA_000006945.2), which were selected from 1785 published whole genomes in NCBI database. The selection procedure was as follows: first, the 1785 public genome data were analyzed according to these seven alleles (aroC, dnaN, hemD, hisD, purE, sucA, and thrA) using PubMLST website database. Then, the additional 317 ST34 Salmonella Typhimurium and variant strains were classified from the 1785 public genome data. Details are shown in Supplementary Data S1. The loci were schemed according to the SalmcgMLST v1.0 locus. The dendrogram of these strains was graphed by ggplot2 in R.
Analyses of flagella (H antigen)-related genes
The flagella (major differences of H antigen between Salmonella Typhimurium and variants) encoding genes of the 14 strains of this study and 317 related ST34 Salmonella Typhimurium and variant strains were annotated by eggNOG-mapper v2 (http://eggnog-mapper.embl.de/). All antigenic formulas of the ST34 Salmonella Typhimurium and variant strains were confirmed using bioinformatics technique Seqsero2 (http://www.denglab.info/SeqSero2).
Of the 331 ST34 strains analyzed, 24 strains were found to have lost their O antigen information based on the assembled sequence. The remaining 307 ST34 strains were used for the analyses of flagella (H antigen)-related genes. Fifty-eight flagella encoding genes were selected according to the eggNOG-mapper gene description information. The presentation of the 58 flagella encoding genes was compared among the ST34 Salmonella Typhimurium and variant strains.
Results
Characterization of 14 Salmonella Typhimurium and variant strains
An interesting phenomenon was found in the three groups of Salmonella Typhimurium and variant strains; the H antigens of phase one “i” and of phase two “1,2” could be lost or both lost, but they had 100% PFGE genetic similarity. PFGE results showed that the 12 strains were divided into three groups with the 100% similarity; the other two strains “F42” and “F120” were separated individually. Three different antigenic formula strains were clustered into the first PFGE 100% similarity group (ID F15, F38, and F59), seven strains constituted the second 100% similarity group (ID F1, P1, F7, P7, F33, F153, and P212), and two strains constituted the third one (ID P286 and P287).
The first group was 93.3% similar to the second group; the third group was 71.9% similar to both the groups 1 and 2. This irregularity reveals that these 12 Salmonella Typhimurium strains and Salmonella Typhimurium variant strains were evolved from three different origins, whereas the same ancestor of three sources produced different antigenic formulas, respectively. Details are shown in Figure 1.
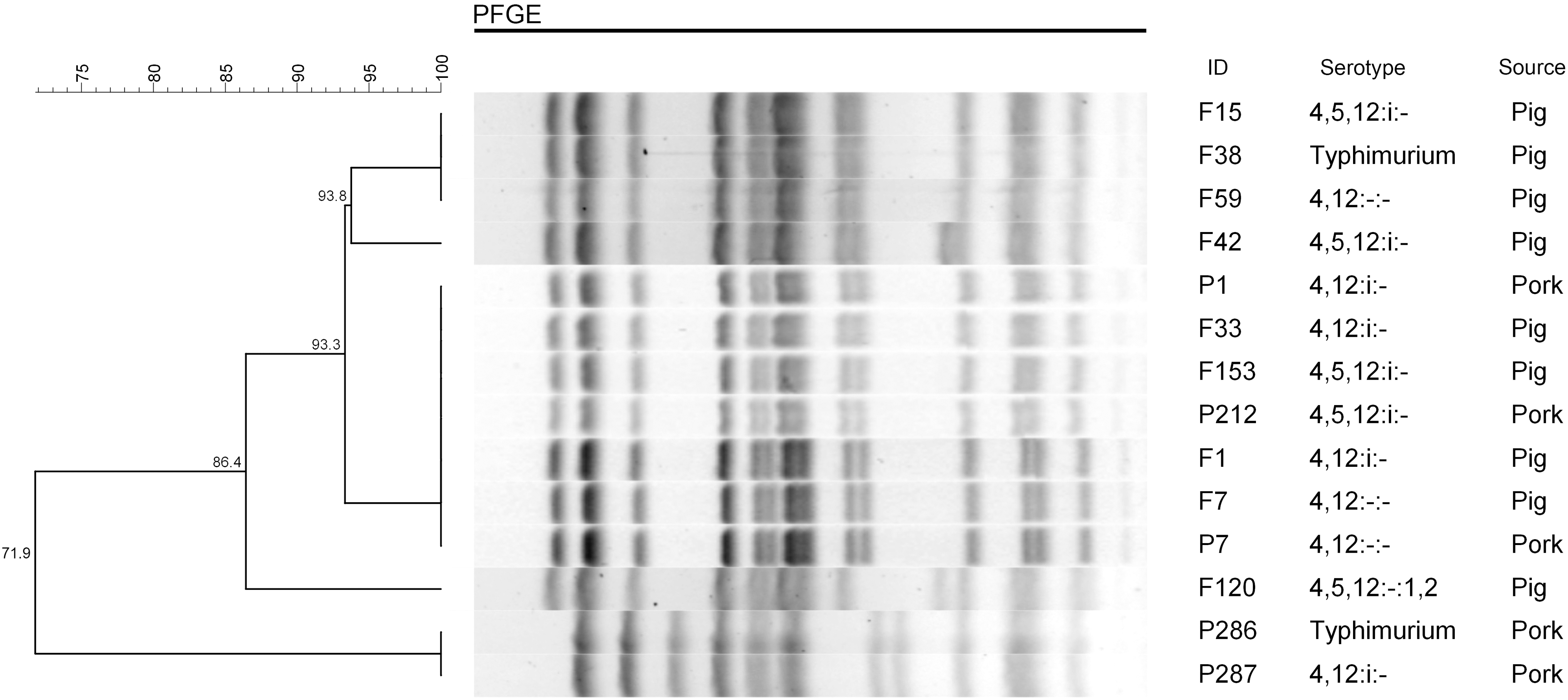
Some different antigenic formula strains exhibited the 100% PFGE similarity among 14 Salmonella Typhimurium and variant strains. PFGE, pulsed-field gel electrophoresis.
Further phylogenetic analyses
To improve the discrimination power of the phylogenetic relationship analyses, the FTIR and wgMLST assays were used. The FTIR technique holds high sensitivity in detecting phenotype changes in the functional groups according to the “fingerprint region” of bacteria. The fingerprint regions are varied based on the spectra information on cellular components such as nucleic acid (antisymmetric and symmetric PO2 − stretching mode from 1200 to 1000 cm−1), ester lipid (CO stretching of 1740 cm−1), protein (stretching of CN and NH from 1700 to 1500 cm−1), and lipid (asymmetric and symmetric vibrations of CH2 and CH3 from 3000 to 2800 cm−1).
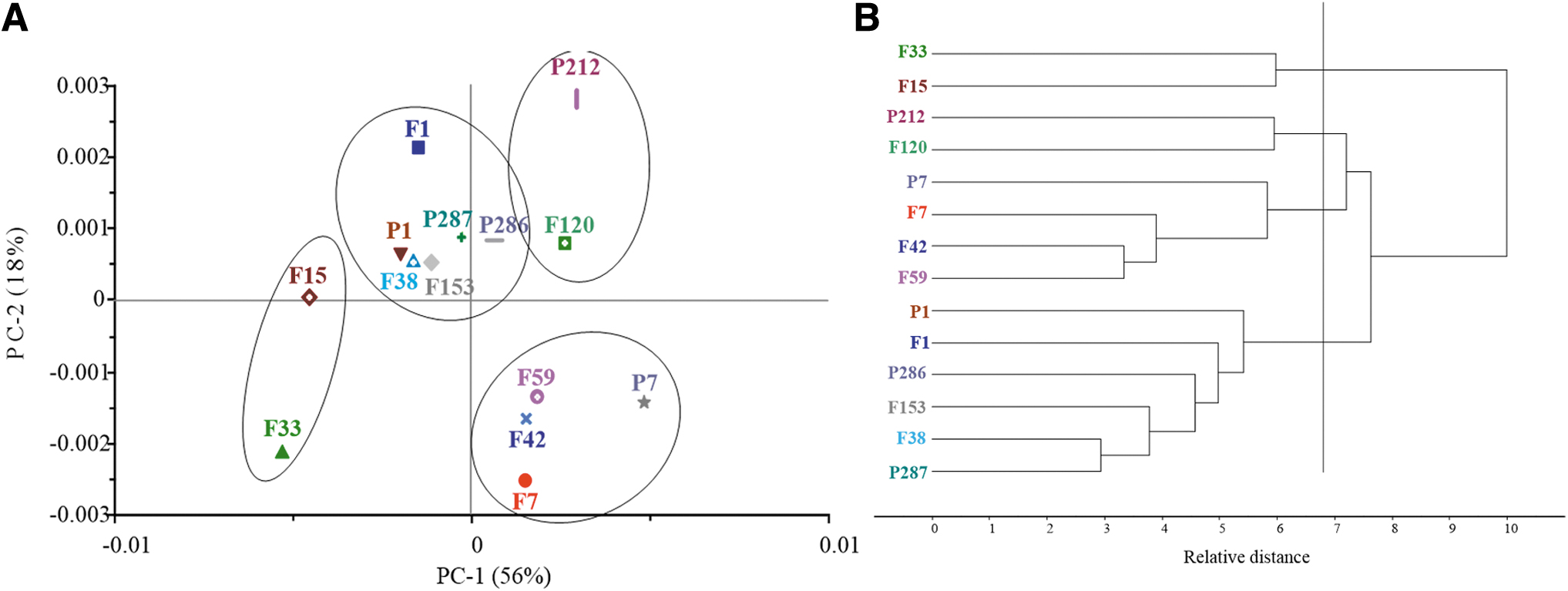
The spectral data of these strains were analyzed by PCA and HCA. The PCA result has been performed to explain the difference of principal components among samples. The HCA result was applied to identify the relationships among the different samples. PCA of FTIR results showed, PCA factors 1 and 2 explained 74% of the total variance (56% plus 18%). The PCA score plot revealed that the 14 strains can be clustered into four groups. Group 1 included F15 and F33, group 2 involved F120 and P212, group 3 involved F42, F7, P7, and F59; while group 4 included P1, F1, F38, F153, P286, and P287. HCA of FTIR results shown the 14 strains were divided into 4 groups at the 7 relative distance.
The first group contained F33 and F15, the second group were P212 and F120, the third group were composed of four strains (ID F42, F7, P7, and F59), and the last one was composed of six strains (ID P1, F1, F38, F153, P286, and P287). The HCA and PCA results were related to each other, indicating that the members of the group could be matched between HCA and PCA (group 1 ID F7, P7, and F59; group 2 ID P1, F1, F38, F153, P286, and P287). However, the results of the two group isolates also showed an unrelated relationship between antigenic formulas and polyphasic subtypes. The group one contained Salmonella Typhimurium variant 1,4,[5],12:-:-, and 1,4,[5],12:i:-. The second group contained Salmonella Typhimurium and variant 1,4,[5],12:i:-. It was demonstrated that these Salmonella Typhimurium isolates and variants hold a high similarity. Details were shown in Figure 2.
For the wgMLST results, the whole genome of 14 Salmonella Typhimurium and variant strains was typed as ST34 according to the multilocus (aroC, dnaN, hemD, hisD, purE, sucA, and thrA). Then, the other 50 strain public ST34 data were selected according to the sample information; results are shown in Figure 3. The total 65 strains (50 strain public data, 14 strains of this study, and one reference strain) were sorted into three major groups (a, b, c) as shown in Figure 3. The most strains were found in group b, of which the antigenic formulas were not matched with the genetic relationship. The distance matrix is shown in Supplementary Data S1.
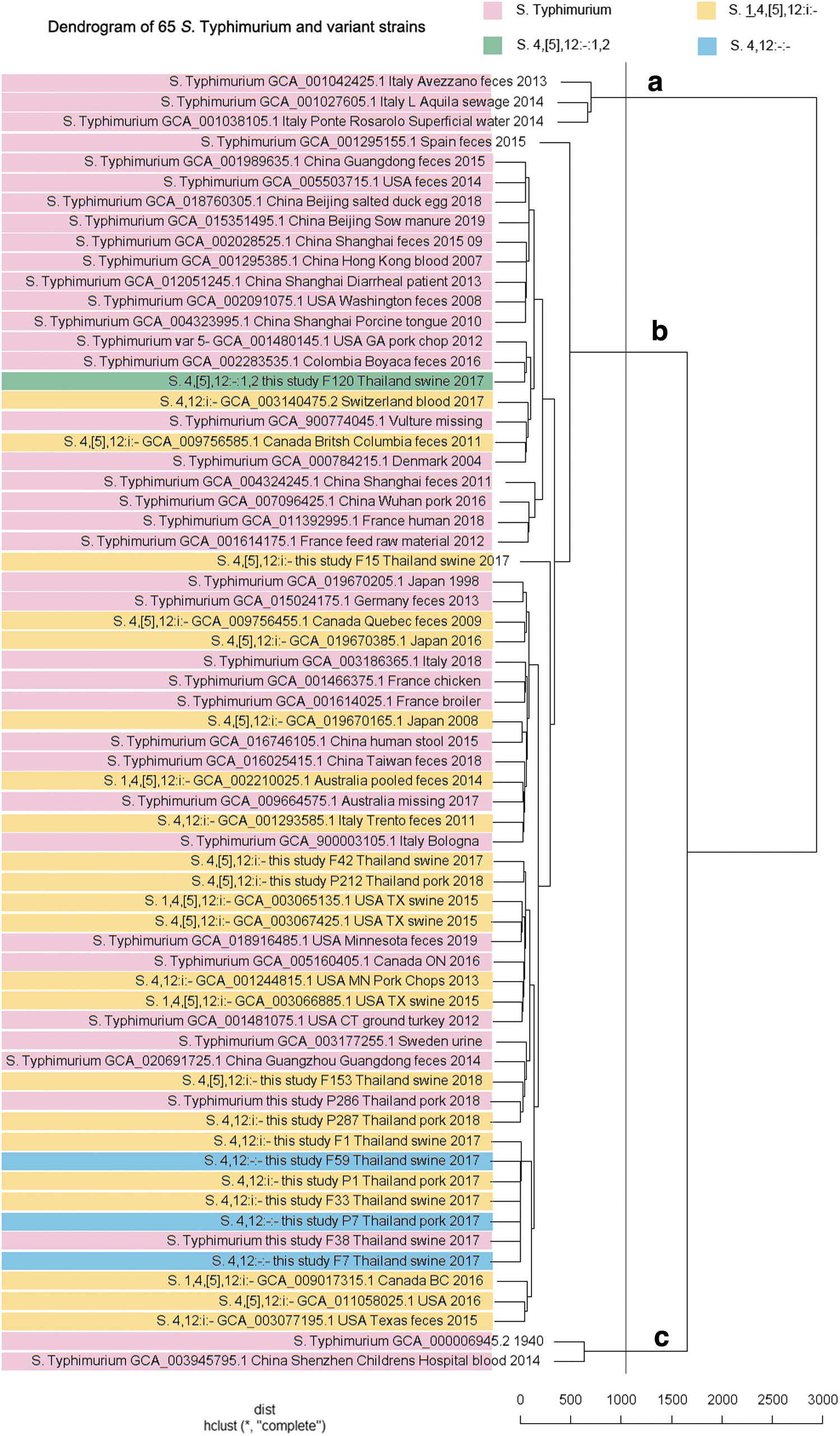
Dendrogram of 65 Salmonella Typhimurium and variant strains (50 strains public data, 14 strains of this study, and 1 reference strain), clustered by wgMLST. wgMLST, whole-genome multilocus sequence typing.
Analyses of flagella antigen-related genes
Fifty-eight flagella encoding genes of the 307 ST34 strains were assembled and annotated by eggNOG-mapper database (shown in Supplementary Data S1). The 307 ST34 Salmonella Typhimurium and variant strains include 236 strains of Salmonella Typhimurium, 66 strains of Salmonella 1,4,[5],12:i:-, three 1,4,[5],12:-:-, and one 1,4,[5],12:-:1,2. Fifty-eight flagella encoding genes were as follows: cheY, epaP, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fljB, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, fljA, flk, hin, mdh, motA, motB, rtsB, rtsA (envY), spaO, spaP, spaQ, spaR, spaS, ssaQ, ssaS, and ycgR.
The flagella encoding gene patterns among the Salmonella Typhimurium and variant strains are listed in Table 1; total 13 patterns were discovered. The Salmonella Typhimurium strains covered the whole 13 patterns; the 1,4,[5],12:i:- were pattern 5 (hin negative), pattern 6 (hin, fljB negative), pattern 9 (fljA, hin, fljB negative), pattern 10 (ssaS, fljA, hin, fljB negative); while the 1,4,[5],12:-:1,2, and 1,4,[5],12:-:- strains were found as pattern 9 (fljA, hin, fljB negative). No regularity was found between the flagellar antigenic formula and flagella encoding genes, according to the 307 whole-genome data. The same flagella encoding gene patterns could express the different antigenic formulas. For example, the pattern 9 (fljA, hin, fljB negative), it is expressed in all the Salmonella Typhimurium and variant serotypes.
Combination of Flagella Antigen-Related Gene Patterns
Pattern 1 (P1): (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fljB, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, fljA, mdh, hin.
Pattern 2 (P2): (−): fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, fljA, mdh, hin.
Pattern 3 (P3): (−): ssaS; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fljB, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ycgR, fljA, mdh, hin.
Pattern 4 (P4): (−): ssaS, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ycgR, fljA, mdh, hin.
Pattern 5 (P5): (−): hin; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fljB, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, fljA, mdh.
Pattern 6 (P6): (−): hin, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, fljA, mdh.
Pattern 7 (P7): (−): ssaS, hin; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fljB, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ycgR, fljA, mdh.
Pattern 8 (P8): (−): ssaS, hin, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ycgR, fljA, mdh.
Pattern 9 (P9): (−): fljA, hin, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, mdh.
Pattern 10 (P10): (−): ssaS, fljA, hin, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ycgR, mdh.
Pattern 11 (P11): (−): envY, fljA, hin, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, mdh.
Pattern 12 (P12): (−): flgM, fljA, hin, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliQ, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, mdh.
Pattern 13 (P13): (−): fliQ, flgM, fljA, hin, fljB; (+): cheY, flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgN, flhA, flhB, flhC, flhD, flhE, fliA, fliB, fliC, fliD, fliE, fliF, fliG, fliH, fliJ, fliK, fliL, fliM, fliN, fliO, fliP, fliR, fliS, fliT, fliZ, flk, motA, motB, envY, rtsB, spaO, spaP, spaQ, spaR, spaS, ssaQ, epaP, ssaS, ycgR, mdh.
Discussion
The serotyping of Salmonella Typhimurium and its variants has always been a challenge. In our previous work, we found that in 14 ST34 Salmonella enterica Typhimurium and variants, antigenic formulas do not match their PFGE evolutionary relationships, which means that the H antigens of phase one “i” and of phase two “1,2” could be lost or both lost, but they had 100% PFGE genetic similarity. To further explore the discrepancy between the antigenic formulas and the PFGE subtypes, we improved the discrimination for the phylogenetic relationship (using FTIR and wgMLST) among the 14 Salmonella Typhimurium and variant strains. Then, we compared the whole genomes of 14 strains (this study) and the other 293 ST34 strains of Salmonella Typhimurium and variant strains for their flagella encoding genes.
The PFGE results suggested that the 14 strains originated from several different sources, but the same ancestor produced different antigenic formulas, respectively, indicating that different antigenic formulas (Salmonella Typhimurium and variant strains) could have 100% similarity. To improve the discrimination power on analyzing the phylogenetic relationship, the FTIR and wgMLST assays were used. However, The FTIR results showed no regularity between the antigenic formulas and genotypes (or phenotypes).
For the wgMLST assay, these 14 strains were first sorted into ST34 according to the MLST results. Then, these 14 strains were compared with public data. Similarly, the higher discriminatory power did not reveal any consistency between the antigenic formulas and genotypes. The different antigenic formulas could be more genetically close than the same antigenic formula among Salmonella Typhimurium and variant strains. The phenotype results also showed the same phenomenon: there was no regularity between the antigenic formulas and relationships among Salmonella Typhimurium and variant strains. That is to say, even the antigenic formulas of Salmonella Typhimurium and variant strains were different; this was possible from the same ancestor.
To uncover the secret of this phenomenon, we compared the flagella (H antigen) encoding genes between the different antigenic formulas. We compared 58 flagella encoding genes and sorted them into 13 flagella encoding gene patterns. However, we found a similar phenomenon: the same flagella encoding gene patterns could express different antigenic formulas. Based on these results, the Salmonella Typhimurium and variant strains revealed a high phylogenetic relationship. This work was consistent with Bugarel's work: they introduced seven gene markers to identify the different antigenic formulas, but they found no consistency either (Bugarel et al., 2012). It was previously suggested that the gene fliC encodes the first flagellar phase and the gene fljB encodes the second flagellar phase.
The hin gene regulates both of these phase encoding genes (Switt et al., 2009). However, some strains in this study, such as Salmonella 1,4,[5],12:-:1,2 and S. 1,4,[5],12:-:-, expressed fliC but not the first flagellar phase. This was a paradoxical example of fliC-positive but first-phase-negative strains. Similarly, some strains, such as Salmonella Typhimurium and Salmonella 1,4,[5],12:-:1,2, did not express fljB but showed the second flagellar phase. This was another paradoxical example of fljB-negative but second-phase-positive strains.
Flagella response for bacterial motility and antigenicity. Generally, the absence of the flagella is thought to help Salmonella evade the host immune system. The role of flagella in triggering an immune response has been argued as important in reducing the immune response during early invasion and systemic spread (Barrow and Methner, 2012). It was reported that the Salmonella serotypes Gallinarum and Pullorum, which cause typhoidal infections in birds, lacked flagellin expression (Barrow and Methner, 2012).
Salmonella can also downregulate flagellin to prevent T-cell activation and antigen processing by dendritic cells (Bernal-Bayard and Ramos-Morales, 2018). Thus, it is speculated that the change of flagella antigen of Salmonella Typhimurium and variants might be related to evading host immune response. Based on the PFGE dendrogram and the PCA and HCA of FTIR results of 14 strains in this study, and the wgMLST results of 65 strains (14 strains genome of this study, 51 strains of public genome data), we can conclude that the serotyping assay of Salmonella Typhimurium and its variants did not match their evolutionary relationship.
In our opinion, we concur with Chattaway et al.'s (2021) suggestion that reporting Salmonella enterica subtypes using the ST followed by the serotype might be more appropriate. Finally, this work was limited to the transcriptional-level studies, because of the limited samples. The transcriptional changes affecting H antigen will be explored in further studies.
Conclusion
We found no consistency between the serotypes and phylogenetic relationships among ST34 Salmonella Typhimurium and variant strains. Even when we screened 58 flagella antigen-related genes, we found no consistency between the flagella antigenic formula and flagella antigen-related genes. Based on these findings, it is speculated that similar phenomena may exist in other ST subtypes. Further exploration of different ST subtypes of these strains is needed to derive this inference. If this phenomenon is widely present in Salmonella Typhimurium and variant strains, it can be inferred that the serotype differentiation among Salmonella Typhimurium and variant strains has insufficient serotyping classification significance.
Footnotes
Acknowledgments
The authors thank the Knorigene Technologies Ltd., the WHO National Salmonella and Shigella Center (NSSC), Thailand, and the Synchrotron Light Research Institute (Public Organization), Thailand, for their kind support.
Authors' Contributions
X.W. contributed to funding acquisition, writing—original draft, and investigation (lead); F.S. performed conceptualization, writing—review and editing, and resources (lead); P.P. carried out formal analysis and visualization (lead); S.S. performed data curation and methodology (lead); H.L. rendered methodology (equal); W.L. provided software (lead); S.A. assisted with project administration, supervision, and validation (lead).
Disclosure Statement
This work was supported by the grants from Yunnan Province Science and technology department, the Key Research and Development Program [grant number 202203AK140017; Major Science and Technology Project in Yunnan Province “Research and Application of Large scale Healthy Pig Breeding Technology” [grant number 202102AE090026]; the Yunnan Joint International R&D Center of Veterinary Public Health [grant number 2023BI60619]; and Open Fund Project from Engineering Research Center for the Prevention and Control of Animal Original Zoonosis of Fujian Province University, College of Life Science, Longyan University [grant number 2022K001].
Funding Information
This work was supported by two grants from Yunnan Province Science and technology department, the Key Research and Development Program [Grant No. 202203AK140017] and the Yunnan Joint International R&D Center of Veterinary Public Health [Grant No. 2023BI60619].
Supplementary Material
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.