
Abstract
Desiccation stress could induce crossprotection and even affect virulence of Salmonella enterica. However, the influence of food matrices with low-water activity on desiccation adaptation of Salmonella still remains unclear. This study investigated the survival and adaptation of Salmonella Enteritidis in skim milk powder, ginger powder, and chocolate powder under desiccation storage conditions for a total of 12 weeks. High survival rates of Salmonella Enteritidis in all food matrices maintained over the long-term desiccation storage. Desiccation-adapted Salmonella Enteritidis enhanced heat resistance (p < 0.05) with the increase of storage time. Food composition plays an important role in the induction of crossresistance of desiccation-adapted Salmonella. After desiccation storage, Salmonella Enteritidis in ginger powder was most tolerant to heat treatment. Salmonella Enteritidis in skim milk powder was most resistant to the gastrointestinal simulation environment, and had strongest adhesion to Caco-2 cells. The effects of food composition on gene expression (rpoS, proV, otsA, otsB, grpE, dnaK, rpoH, and sigDE) in desiccation-adapted Salmonella Enteritidis were not significant (p > 0.05). At initial desiccation storage, osmotic protection-related genes (fadA, proV, otsA, and otsB), stress response regulator (rpoS), and heat-resistance–related genes (grpE, dnaK, and rpoH) were all significantly upregulated (p < 0.05). However, after 4-week storage, the expression level of desiccation-related genes, proV, otsA, otsB, grpE, dnaK, and rpoH, significantly decreased (p < 0.05). This study enables a better understanding of Salmonella's responses to long-term desiccation stress in different kinds of low-water activity foods.
Introduction
Low-water activity foods (LWFs) are generally considered unsuitable for the growth of pathogenic bacteria, including Salmonella. However, Salmonella enterica has been found to be highly adaptable to drying conditions encountered in LWFs (Finn et al., 2013a). Salmonellosis outbreaks have been linked to consumption of a variety of LWFs, such as infant milk products (Jourdan-da Silva et al., 2018), chocolate (Werber et al., 2005), and spices (Jernberg et al., 2015). Multiple serotypes such as Salmonella Enteritidis, Tennessee, and Agona have been associated with Salmonella contamination in LWFs (Abdelhamid and Yousef, 2021). It is reported that desiccated Salmonella Enteritidis acquired more tolerance to multiple stressors than other serotypes (Gruzdev et al., 2011).
The exposure of Salmonella to low-water activity induces crosstolerance to other environmental stresses, such as heat, acid, and antibiotics (Morasi et al., 2022). A recent study (Dhaliwal et al., 2021) shows that water activity and food composition may significantly influence the thermal resistance of Salmonella, in which Salmonella Enteritidis displayed the highest desiccation and thermal resistance in LWF. Food matrix is highly relevant to the survival of desiccated Salmonella. The uptake of extracellular compatible solutes from food ingredients plays an essential role in the protection of biological macromolecules in Salmonella cells during desiccation stress (Dhaliwal et al., 2021).
Salmonella can survive well in LWFs such as almond and peanut, with enhanced heat tolerance after long-term storage (Pereira et al., 2020; Zhu et al., 2021). With the protection of food matrices, Salmonella can even tolerate extremely acidic conditions (Waterman and Small, 1998; Ye et al., 2019). Moreover, stress adaptation of pathogens in food matrix may facilitate their survival through the gastrointestinal system (NicAogáin and O'Byrne, 2016).
Desiccation could induce regulatory variations in expression of genes involved in stress response, such as alternative sigma factors, heat shock response genes, and osmotic regulators (Abdelhamid and Yousef, 2020; Finn et al., 2013a). These changes may help elucidate the mechanism of desiccation adaptation of Salmonella. Studies have been conducted in food matrices to identify deferentially expressed genes induced after exposure of Salmonella to desiccation conditions (Crucello et al., 2019; Deng et al., 2012). Nevertheless, few studies have examined the expression of adaptation-related genes during long-term storage in different LWF products.
In this study, skim milk powder, chocolate powder, and ginger powder were selected for investigation as these LWF products have been previously involved in Salmonella contamination and product recall. The responses of Salmonella Enteritidis were assessed after inoculation in food products for a total of 12-week desiccation storage.
Materials and Methods
Bacterial strains and inoculum preparation
Salmonella enterica serotype Enteritidis CICC 21482 used in this study was obtained from China Center of Industrial Culture Collection. Salmonella Enteritidis strain was inoculated into Tryptic Soy Broth (TSB; Huankai Biotechnology Co., Guangzhou, China) and incubated at 37°C for 18 h. Then, 1 mL of cell suspensions were spread on Tryptic Soy Agar (TSA) and incubated at 37°C for 24 h. Colonies on the plate were harvested with 5 mL of sterile phosphate-buffered solution (PBS, pH = 7.4). The concentration of the obtained stationary-phase Salmonella inoculum (∼11 Log colony-forming unit [CFU]/mL) was determined by plating serial dilutions on TSA.
Dehydration conditions and inoculation
Skim milk powder (Anchor; Fonterra Ltd., Auckland, New Zealand), ginger powder (Xinran Foods Co., Ltd., Hangzhou, China), and chocolate powder (AIK CHEONG SDN. BHD., Melaka, Malaysia) were purchased from grocery stores. The protein, fat, and carbohydrate contents of the three foods are provided in Supplementary Table S1 in the Supplementary Data. These food products were spread in uncovered culture dishes, respectively, followed by irradiation with an ultraviolet (UV) lamp for 30 min to reduce the background bacteria. The three powders were then mixed with buffered peptone water (BPW) and plated on xylose lysine deoxycholate agar (XLD) to ensure that the samples were not contaminated with Salmonella.
The water activity of food substrates was maintained at 0.30 ± 0.05 (26.0°C ± 1.0°C) in a desiccator. A modified sample inoculation process was adapted from the method of Liu et al. (2019). In brief, 1 mL of 11 Log CFU/mL bacterial inoculum was mixed with 10 g of food substrates, respectively, before thorough mixing in sterile homogeneous bags. Inoculated food matrices were transferred to sterile petri dishes and dried by silica gel beads in a desiccator chamber at room temperature for 24–48 h to stabilize the water activity at 0.30 ± 0.05.
Ten grams portions of the redried inoculated food samples were crushed with a mortar and pestle to eliminate clumping, and then evenly mixed into 90 g of uninoculated food matrix, respectively. To evaluate the survival rate of Salmonella Enteritidis, 1.00 ± 0.05 g of each sample was transferred into 15 mL sterile centrifuge tube, sealed, and stored in the desiccator. At each of the time points (0, 1, 2, 3, 4, 6, 8, 10, and 12 weeks), the centrifuge tubes were removed from the dryer, followed by adding 9 mL of BPW and vortexing for 60 s. The obtained Salmonella population was quantified by plate counting on XLD.
Thermal treatment
Inoculated foods were collected for heat resistance test at 0-, 4-, and 8-week storage, respectively. At each sampling time point, the test sample was heated in a water bath at 80°C with a holding time of 1 min. One gram of test sample was drawn at certain time intervals after heating for each food matrix. After treatment, the samples were immediately cooled down in an ice water bath for 1 min to stop heat stress. The population of surviving Salmonella in heated samples was determined by serial dilution in BPW and enumeration on XLD plates. The time required to kill 90% of the Salmonella population (D-value) under heat treatment was calculated to demonstrate the thermal resistance of Salmonella cells.
Survival of Salmonella Enteritidis through a simulated gastrointestinal system
Human gastrointestinal environment was simulated using a standardized in vitro digestion model (Brodkorb et al., 2019). Detailed experimental procedures can be found in the Supplementary Data. In brief, 1.0 g of each sample stored at 0-, 4-, and 8-week time points was added to a gastric simulation system, which was adjusted to pH 3.0, and then incubated in a shaking incubator (100 RPM) at 37°C for 40 min. After the gastric phase, the digestive juice was adjusted to pH 7.0, transferred to an intestinal simulation system, and then incubated at 37°C for 120 min at 100 RPM. Enumerations of Salmonella Enteritidis were performed at the beginning and at the end of each digestion stage by plate counting.
Bacterial adhesion and invasion assay
Human colon adenocarcinomas Caco-2 cells were used for the bacterial adhesion and invasion assay. Caco-2 cells (105 cells/mL) were seeded into 24-well plates and cultured in a cell culture box with 5% (v/v) CO2 at 37°C until the wells were fully covered with cells. Caco-2 cells were washed with PBS three times and inoculated in the culture medium without penicillin–streptomycin solution.
Gentamicin protection assay was performed on a 24-well plate affixed with monolayer Caco-2 cells to determine the adhesion and invasion ability of Salmonella Enteritidis as described previously (Gagnon et al., 2013) with details provided in Supplementary Data. The adhered and invasive bacterial populations were determined by plate counting. The adhesion and invasion abilities were calculated, respectively, as follows:

RNA extraction and reverse transcription quantitative PCR
Bacterial suspensions obtained from inoculated foods at 0- and 4-week storage were collected for RNA extraction, respectively. RNA extraction was performed using PowerMicrobiome™ RNA Isolation Kit (MO BIO Laboratories, Inc., San Diego, CA) according to the manufacturer's instructions. The quality of RNA was determined using ultramicro spectrophotometer (Hangzhou Allsheng Instrument Co., Ltd., Hangzhou, China). Qualified RNA was converted to complementary DNA (cDNA) using PrimeScript™ reverse transcription reagent Kit with genomic DNA (gDNA) Eraser (Takara, Kusatsu, Japan). Gene expression assays were executed through a real-time polymerase chain reaction (PCR) system (CFX Connect™; Bio-Rad).
Each 20 μL of reverse transcription quantitative PCR contained 10.0 μL TB Green Premix Ex Taq II (Tli RNaseH Plus; 2X; Takara), 0.8 μL of 10 μM of each forward and reverse prime, 2.0 μL cDNA template, and 6.4 μL sterilized distilled water. Primers and PCR procedures are shown in Supplementary Table S2 in the Supplementary Data. The relative expression levels of the target genes were calculated using the Livak method (2−ΔΔCT) (Livak and Schmittgen, 2001). Salmonella Enteritidis cultured in TSB was used as the calibration sample, and 16S ribosomal RNA (rRNA) was used as the internal reference gene.
Data analysis
D-values were calculated using the equation D = −1/slope for the regression plots of surviving Salmonella populations (Log CFU/g) versus treatment times (min). The results were expressed as means ± standard deviations of three biological replicates. Analysis of variance was performed with Fisher's Least Significant Difference test in SPSS 22.0 to determine significant differences between groups (p < 0.05).
Results and Discussion
Survival of Salmonella Enteritidis in desiccated foods
The inoculation amount of Salmonella Enteritidis in the three foods was ∼9 Log CFU/g. When aw of the food matrix dropped to 0.30 (∼24 h), the counts of Salmonella Enteritidis decreased by 0.72 ± 0.19 Log CFU/g in chocolate powder, 2.2 ± 0.22 Log CFU/g in skim milk powder, and 0.84 ± 0.20 Log CFU/g in ginger powder, respectively (Fig. 1).
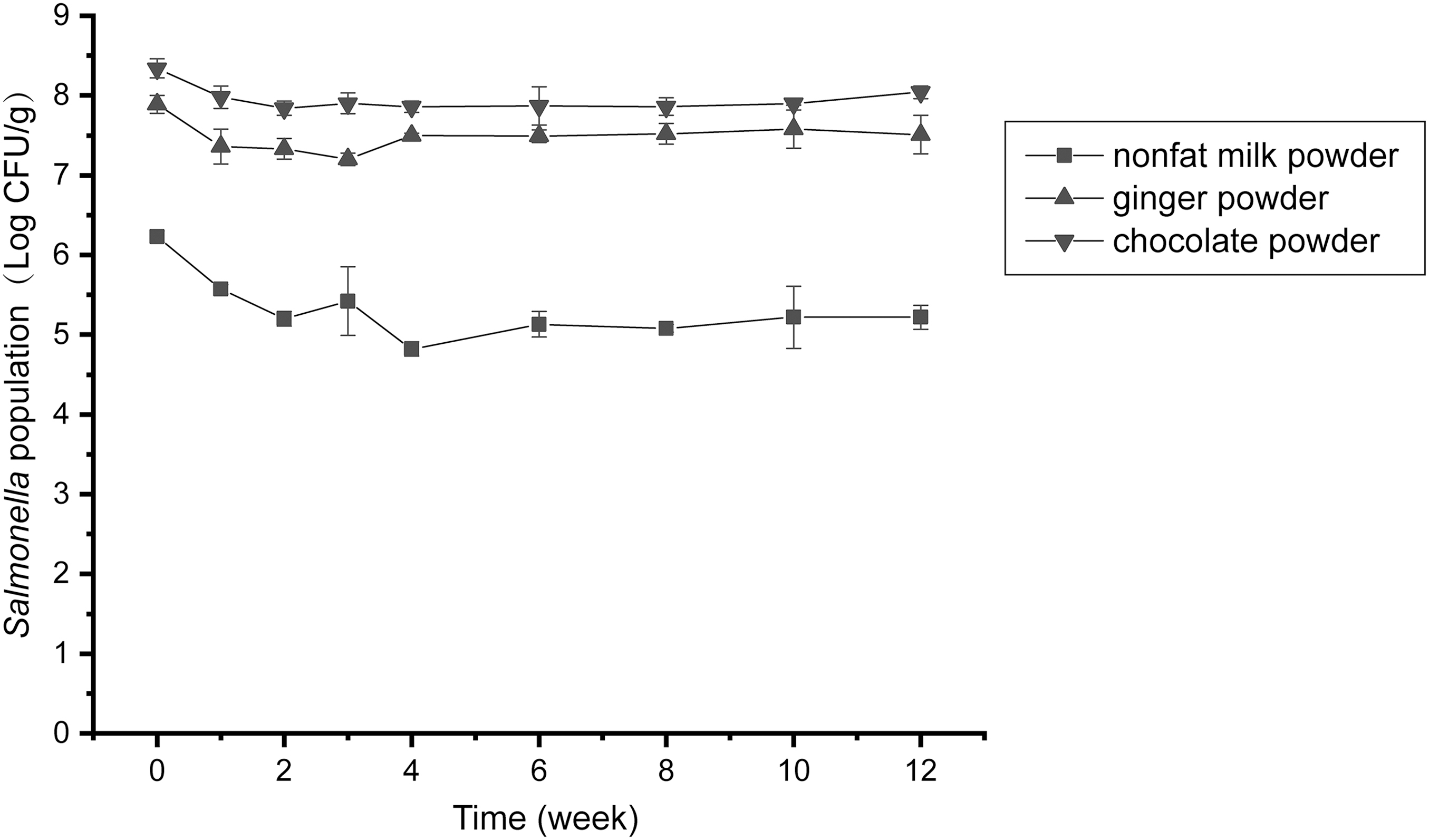
Survival of Salmonella Enteritidis in different food matrices at aw 0.30 during 12-week storage at 26°C. Skim milk powder, square; ginger powder, upper triangle; chocolate powder, lower triangle. Data shown are expressed as means ± standard deviations of three parallel experiments.
Similarly, it has been reported that Salmonella populations remained relatively constant throughout long-term storage on dried ginger slices when aw stabilized to 0.31 (Gradl et al., 2015). The viable population of Salmonella Enteritidis in skim milk powder (6.23 ± 0.07 Log CFU/g) declined significantly (p < 0.05) when compared with chocolate powder (8.34 ± 0.12 Log CFU/g) and ginger powder (7.89 ± 0.11 Log CFU/g). These results indicate that food matrices play an important role in the survival of Salmonella Enteritidis during the drying process.
The population of viable Salmonella Enteritidis in three food substrates all showed a slight decrease at early stage of storage, with a reduction of 0.44 ± 0.22, 0.81 ± 0.50, and 0.69 ± 0.19 Log CFU/g in chocolate powder, skimmed milk powder, and ginger powder at the fourth week, respectively (Fig. 1). High survival rates of Salmonella were observed, with no significant differences among food matrices (p > 0.05). Previous studies have also reported that Salmonella can maintain a high survival rate during storage process in various dry foods such as ground black pepper (Keller et al., 2013), wheat powder (Forghani et al., 2019), and milk powder (Sekhon et al., 2021).
Thermal resistance of Salmonella Enteritidis differed in LWF during long-term desiccation storage
Heat tolerance of Salmonella Enteritidis was investigated at 80°C (Fig. 2). The survival of Salmonella Enteritidis in all food matrices decreased log-linearly with the increase of heat treatment time. Similar trends of thermal resistance have been reported on Salmonella survival in low-moisture high-sugar products (Alshammari et al., 2020) and in skim milk powder products (Liu et al., 2019). As D-values shown in Table 1, heat resistance of Salmonella Enteritidis increased significantly with storage time in all three food matrices (p < 0.01). The D-value of Salmonella inoculated in chocolate powder at 8-week storage was significantly higher than that in skim milk (p < 0.05).
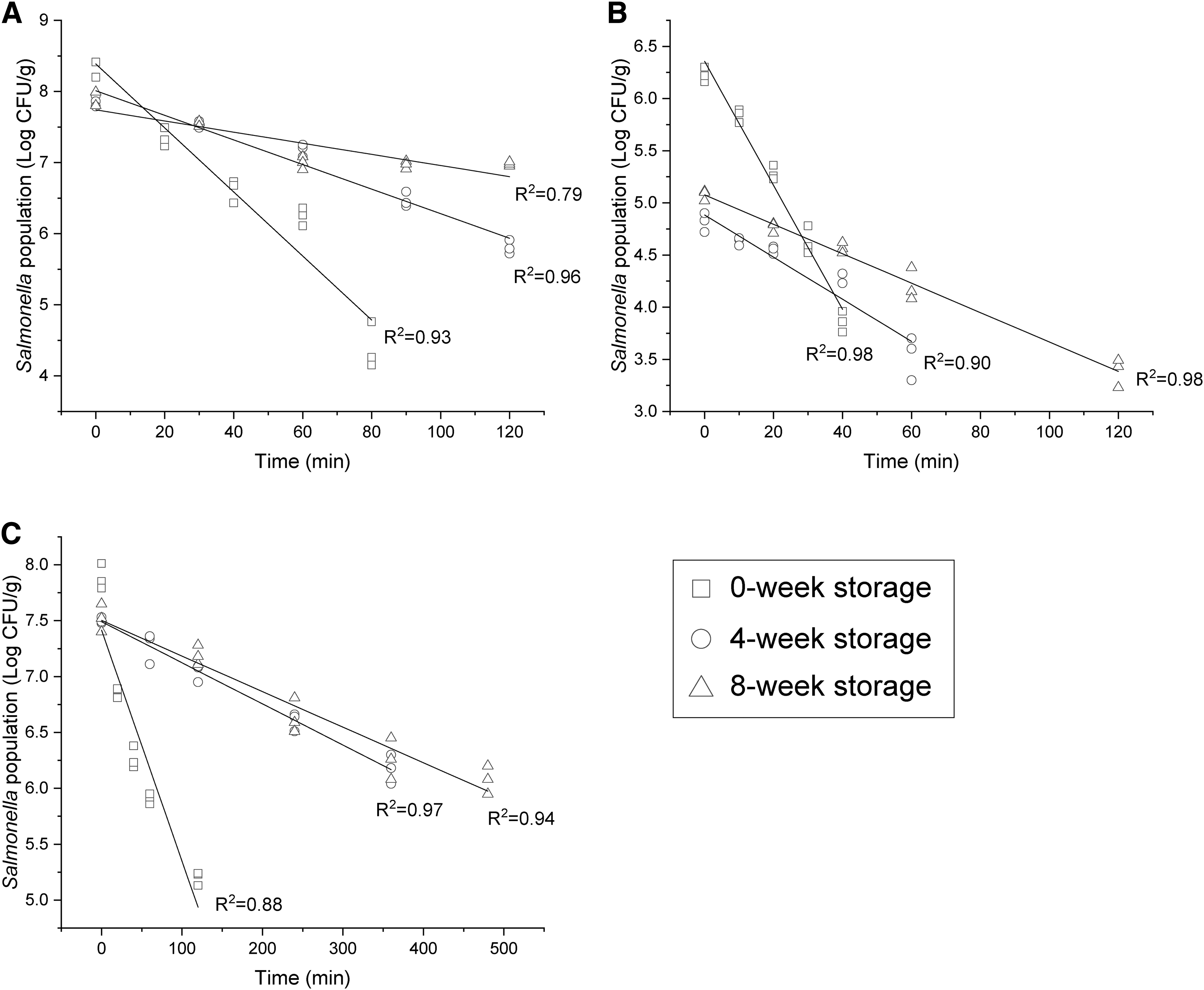
Survival curves of Salmonella Enteritidis in chocolate powder
D-Values of Salmonella Enteritidis at 80°C with Different Storage Time and Food Matrices
Different letters indicate significant differences in means ± standard deviations of three biological trials (p < 0.05). D-values were calculated as the negative reciprocal of the slope of the thermal inactive curves.
p < 0.01.
SEM, standard error of the mean.
The thermal tolerance of Salmonella Enteritidis in ginger powder was the highest, with its D-value being significantly higher than that in the other two at the same storage stage (p < 0.05), which may be owing to the crossresistance conferred by Salmonella's adaptation to the natural antibacterial substances in ginger powder. Natural compounds may lead to the development of crossresistance to other stresses in bacteria. For instance, resveratrol treatment could lead to a significant increase in tolerance to heat stress in Staphylococcus aureus and Listeria monocytogenes (Oliveira et al., 2017). Such crossresistance development may also occur in ginger powder that presents mild stress due to antibacterial substances such as gingerol and ginger essential oils (Beristain-Bauza et al., 2019).
Food matrices influenced the survival of Salmonella Enteritidis in the simulated gastrointestinal system
At the gastric digestion phase, the tolerance of Salmonella Enteritidis inoculated in skim milk powder and ginger powder at 8-week storage was lower than that at 0- or 4-week storage (p < 0.05) (Fig. 3A). Salmonella Enteritidis in ginger powder was least tolerant to simulated gastric digestion, whereas Salmonella Enteritidis in skim milk powder was most tolerant. Salmonella Enteritidis in skim milk powder with 0- and 4-week storage only reduced by ∼0.5 Log CFU/g, possibly due to the protection of the protein content in skim milk powder under acidic conditions (Brodkorb et al., 2019).

Reduction of Salmonella Enteritidis during gastric digestion
Simulated intestinal digestion assays showed that the number of viable Salmonella Enteritidis in chocolate powder was significantly higher at 8-week storage than at 0-week storage (p < 0.05) (Fig. 3B), indicating that desiccation-adapted Salmonella Enteritidis may have better tolerance to intestinal environment with the protection of fat contents. Fat may have a protective effect on Salmonella, as cells encased in the hydrophobic lipid portion were more likely to survive under gastrointestinal environment (Aviles et al., 2013). In contrast, ginger powder with low fat content had no protective effect as the population of Salmonella Enteritidis decreased significantly after simulated intestinal digestion. It is worth noting that Salmonella became more tolerant to intestinal stress with the increase of storage time.
Cell adhesion ability of Salmonella Enteritidis increased after desiccation storage
Salmonella Enteritidis passed through intestinal digestion were further used for intestinal cell adhesion and invasion assays. Because of the weak tolerance of Salmonella Enteritidis in the ginger powder to the simulated gastrointestinal digestion, the number of Salmonella cells adhered and invaded to Caco-2 cells was below the detection limit (1.69 CFU/mL). Compared with the control samples (0-week storage), the adhesion of Salmonella Enteritidis to Caco-2 cells significantly increased in both skim milk powder and chocolate powder at 4-week storage (p < 0.05) (Table 2).
Adhesion and Invasion of Salmonella Enteritidis to Caco-2 Cells in Different Food Matrices After In Vitro Digestion
Different letters indicate significant differences in means ± standard deviations obtained from three independent tests (p < 0.05). “−” Represents that the invasion efficiency is lower than detection limitation.
Weak invasiveness of Salmonella Enteritidis in skim milk powder was observed, while it was not detected in chocolate powder (Table 2). Salmonella Enteritidis adapted in skim milk powder had stronger adhesion and invasion abilities to Caco-2 cells, which supports the opinion that protein may serve as a better protective factor than fat under acidic conditions (Waterman and Small, 1998).
Expression of adaptation-related genes altered during desiccation storage
Compared with nondesiccated cells, the expression levels of fadA, rpoS, proV, otsA, otsB, grpE, dnaK, and rpoH genes were significantly upregulated with at least twofold changes in all samples after 4-week desiccation storage (Fig. 4). Interestingly, two expression patterns were observed over the desiccation storage. Compared with samples at 0-week storage, the expression levels of fadA, rpoS, and sigDE increased or remained steady at 4-week storage, while the expression levels of six genes, including otsB, otsA, dnaK, rpoH, grpE, and proV, significantly decreased.

The relative expression levels of fadA
As bacterial metabolism is severely compromised under low-water activity conditions, the induced protective metabolic regulation is important for bacterial survival at the beginning of dehydration process (Lebre et al., 2017). It should be noted that this study was designed to examine Salmonella's responses throughout a certain storage period (from 0 to 4 weeks) under a consistent desiccation condition with water activity stabilized at 0.3. To fully understand the influence of desiccation storage on the induction of crossresistance, the rate of aw reduction and the effect of physiological injury on bacterial cells should be further considered important factors influencing gene expression.
The expression of fadA was found upregulated under desiccation conditions at either 0- or 4-week storage. 3-ketoacyl-coA thiolase encoded by the fadA gene is involved in catabolism of long-chain fatty acids into acetyl-CoA, which is essential for the production of ATP (Li et al., 2012). Moreover, the expression of fadA in ginger powder at 4-week storage was significantly higher than that in the other two (p < 0.05) (Fig. 4A). The expression of fadA was found significantly influenced by food composition (p < 0.05), possibly due to the active ingredients in ginger powder that have affected the fatty acid metabolism level of Salmonella (Anggraini et al., 2021).
The rpoS gene encodes an alternative sigma factor of RNA polymerase for promoter recognition and transcription initiation, which enables bacteria's adaptation to various stress conditions (Dong and Schellhorn, 2010). Existing studies have shown that rpoS gene is upregulated in desiccation-adapted Salmonella (Chen and Jiang, 2017; Chen et al., 2021). Consistently, rpoS was found significantly upregulated in all food matrices (Fig. 4B), which demonstrates the importance of rpoS gene during desiccation adaptation of Salmonella.
The highest upregulation level was observed in proV, an osmotic protection transporter gene (Fig. 4C). The generation of osmotic protective substances is one of the main mechanisms of Salmonella desiccation resistance (Finn et al., 2013a). Under the desiccation stress, bacteria can accumulate osmoprotective solutes, which can help survive by maintaining a balance of osmolarity inside and outside the cells. Upregulated expression of proV gene has been previously reported in desiccation-adapted Salmonella Typhimurium (Chen and Jiang, 2017) and Salmonella Tennessee (Li et al., 2012).
Genes related to trehalose synthesis, that is, otsA and otsB, were significantly upregulated (greater than twofold) in all food matrices (Fig. 4D, E). Previous studies also reported that trehalose biosynthesis-related genes were upregulated when Salmonella was dehydrated on paper or stainless steel for 24 h (Finn et al., 2013b; Li et al., 2012). Trehalose can stabilize dry membranes by preventing the decrease of spacing between membrane lipids under drying conditions (Strom and Kaasen, 1993).
Furthermore, we observed upregulation of heat shock protein gene grpE (Fig. 4F), chaperone protein gene dnaK (Fig. 4G), and heat-shock–associated gene rpoH (Fig. 4H) in desiccation-adapted cells. Significant upregulated expression of these genes has been reported in Salmonella Typhimurium during desiccation adaptation (Chen and Jiang, 2017). The upregulation of heat-resistance genes under desiccation stress might confer Salmonella's tolerance to high temperature.
The expression of sigDE remained steady when compared with nondesiccated cells (Fig. 4I). This indicates that the invasion ability of Salmonella may not be essential for desiccation adaptation in LWF.
Conclusions
Food composition plays an important role in Salmonella's adaptation to desiccation conditions, which significantly influence its thermal resistance, gastrointestinal tolerance, and adhesion ability. In addition, Salmonella Enteritidis may adapt to desiccation environment by regulating the expression of adaptation-related genes. This study enables a better understanding of the mechanism of Salmonella's survival and adaptation in LWF products.
Footnotes
Authors' Contributions
S.L. designed conceptualization, formal analysis, methodology, writing and editing. W.X. contributed to conceptualization, formal analysis, investigation, and writing—original draft and review. M.L. carried out data curation, formal analysis and investigation, and writing—original draft. Z.L. and Z.M. performed formal analysis and investigation. S.C. and Y.Y. carried out formal analysis. H.Z. designed conceptualization, methodology, and writing-review and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (grant Nos. 31972044, 32202186).
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.