
Abstract
Ultraviolet-C light-emitting diode (UVC-LED) and ultrasound (US) are two nonthermal technologies with the potential to destroy pathogens. However, little is known about their effectiveness in strains with a history of heat resistance. Thus, this study aimed to evaluate the phenotype and genotype of heat-resistant extraintestinal pathogenic Escherichia coli (ExPEC) with heat resistance genes after the application of US, UVC-LED, and UVC-LED+US. For this, two central composite rotatable designs were used to optimize the UVC-LED and US conditions in four ExPEC isolated from beef. From the genome of these isolates obtained in a previous study, possible genes for UVC resistance were analyzed. Results showed that US was ineffective in reducing >0.30 log colony-forming unit/mL, and that when used after UVC-LED, it showed a nonsynergic or antagonistic effect. Also, UVC-LED had the greatest effect at the maximum dose (4950 mJ/cm2 from 1.65 mW/cm2 for 50 min). However, the strains showed some recovery after that, which could be implicated in the expression of genes included in SOS system genes, some others present in the transmissible Locus of Stress Tolerance (trxBC and degP), and others (terC). Thus, ExPEC can overcome the conditions used in this study for US, UVC-LED, and UVC-LED+US, probably due to the history of resistance to other cellular damage. The result of this study will contribute to future studies that aim to find better treatment conditions for each food product.
Introduction
Extraintestinal pathogenic Escherichia coli (ExPEC) comprise a group of E. coli strains responsible for numerous infectious diseases in humans (Tetzschner et al., 2020). Some pathogenic E. coli can express stress resistance genes included in the transmissible Locus of Stress Tolerance (tLST) and can resist temperatures up to 80°C (Guragain et al., 2020; Kamal et al., 2021). In this regard, conventional heat treatments are ineffective in these populations (Dlusskaya et al., 2011; Guragain et al., 2020; Machado et al., 2023a).
In contrast, nonthermal technologies (NTTs), such as ultrasound (US) and ultraviolet-C light-emitting diode (UVC-LED), have been evaluated due to reduced impact on food's characteristics, especially sensory and nutrition aspects (Bernardo et al., 2023; Chakka et al., 2021). Also, they are being classified as environmental friendly technologies (Chakka et al., 2021). Furthermore, the combination of NTT is suggested to increase the efficiency of microbial inactivation in food (Char et al., 2010).
Concerning this theme, our previous study was the first to evaluate the combination of UVC-LED+US on E. coli isolated from pasteurized milk and harboring the tLST (Machado et al., 2023b). In contrast, the results of this study revealed that UVC-LED alone showed more promise than UVC-LED+US. Once E. coli is subjected to stressful environments such as oxidative stress caused by UV radiation, damage repair systems can be activated (Hamamoto et al., 2010).
These systems include genes present in the E. coli SOS system, some in the tLST, and other genes within the bacteria genome, such as terC (Hamamoto et al., 2010). However, there is limited information regarding the efficacy of UVC-LED, US, and their combination in heat-resistant populations, as well as their role in this resistance. In this sense, in our previous study, potential genes related to resistance to UVC-LED were not evaluated, which encouraged us to investigate the presence of potential genetic markers with this function.
With this purpose, our study aimed to (1) establish an optimized condition for UVC-LED and US through an inactivation model on heat and moderately resistant ExPEC, carrying and not carrying tLST; (2) evaluate the effect of UVC-LED and US (alone and in combination) on ExPEC strains; (3) verify the presence of main gene markers reported in response to oxidative stress (UVC) between strains from Brazillian beef.
Materials and Methods
Heat-resistant ExPEC strains
Four strains of E. coli (C9, C31, C97, and C1145) previously described (Machado et al., 2023a) and isolated from beef and characterized as ExPEC, since they harbor adherence genes and virulence factors (bfpA, cadA, papA, iss, and ompT) commonly associated with nonintestinal human infections (Tetzschner et al., 2020), were used in this study. These strains were previously evaluated for heat resistance (60°C/6 min) and the presence of tLST through whole genome sequencing (Machado et al., 2023a; Machado et al., 2022). The data are available on the National Center for Biotechnology Information (NCBI) in BioProject PRJNA806981.
UVC-LED and US treatments optimization
To optimize an exposure condition for UVC-LED and US, we used the same model we performed in our previous study (Machado et al., 2023b). For that, two central composite rotatable designs (CCRDs) were performed. Among the four E. coli strains, the isolate described as C31 was the only one previously studied regarding UVC application (Castro et al., 2019), and, therefore, that was selected for the CCRD optimization trials. Thus, two CCRDs were performed to optimize the conditions for UVC-LED and US (Table 1).
Factors and Levels of the Variables Used in the Central Composite Rotatable Design for Power Intensities and Time for Ultraviolet-C Light-Emitting Diodes and Ultrasound
Points of factorial design (−1; +1), axial points (−1.414; +1.414), and central point (0).
US, ultrasound; UVC-LED, ultraviolet-C light-emitting diode.
UVC-LED treatment and screening for UVC genes
The treatments alone or combined with US (UVC-LED+US) were performed after parameters optimization. The assays for UVC-LED were carried out using modules with single pick emissions at 275 nm (Black Box Smart®, BioLambda, Brazil). For this, 1.5 mL of each bacterial suspension in broth heart infusion (BHI) broth (109 log colony-forming unit (CFU)/mL of bacteria) was transferred to 2 mL microtubes. Then, closed microtubes were treated with UVC-LEDs at different intensities and times (Table 1) and at 15 cm from the LED. The optimized treatment was calculated according to the exposure at a given intensity and time (E = P × t, wherein E is dose in mJ/mL, P is irradiance in mW/cm2, and t is time in seconds) (Ghate et al., 2013).
To characterize the presence of some main genes for UVC resistance, including markers for SOS, tLST, and others (Jánošíková et al., 2021; Kamal et al., 2021; Sargentini et al., 2016), we used the gene bank of these isolates provided in our previous study (Machado et al., 2023a).
US treatment
The US treatment alone and in combination (UVC-LED+US) after parameter optimizations were performed using an ultrasonic liquid processor (VCX 750; Sonics Vibra cell™) with a frequency of 20 kHz. Likewise, 1.5 mL of each bacterial suspension in BHI broth (109 log CFU/mL of bacteria) was transferred to 2 mL microtubes. Microtubes were sonicated with a 64 mm-diameter probe. The ultrasonic power corresponded to 299 W and the acoustic energy density was calculated according to Equations 2 and 3 of our previous study (Machado et al., 2023b).
Statistical analysis
The CCRD experimental points obtained were analyzed in Statistica software version 12.0 (StatSoft Power Solutionsâ). Furthermore, two-way analysis of variance (ANOVA) with Tukey test at a significance of 0.05 was used to compare the effect of UVC-LED, US, and their combination between strains. These assays were performed using the software Minitab® version 19.1 (United States).
Results
The time and intensity conditions for UVC-LED performed to isolate C31 corresponded to a linear model (Fig. 1), relating the combination of time (50 min) with intensity (1650 mW/cm2), yielding a UVC-LED dose of 4950 mJ/cm2 (Supplementary Table S1). The CCRD exhibited a nonsignificant lack-of-fit (p < 0.010), R 2 of 0.90, adjusted determination coefficient (R 2 adj.) of 0.86, and model mean square error value of 0.010.
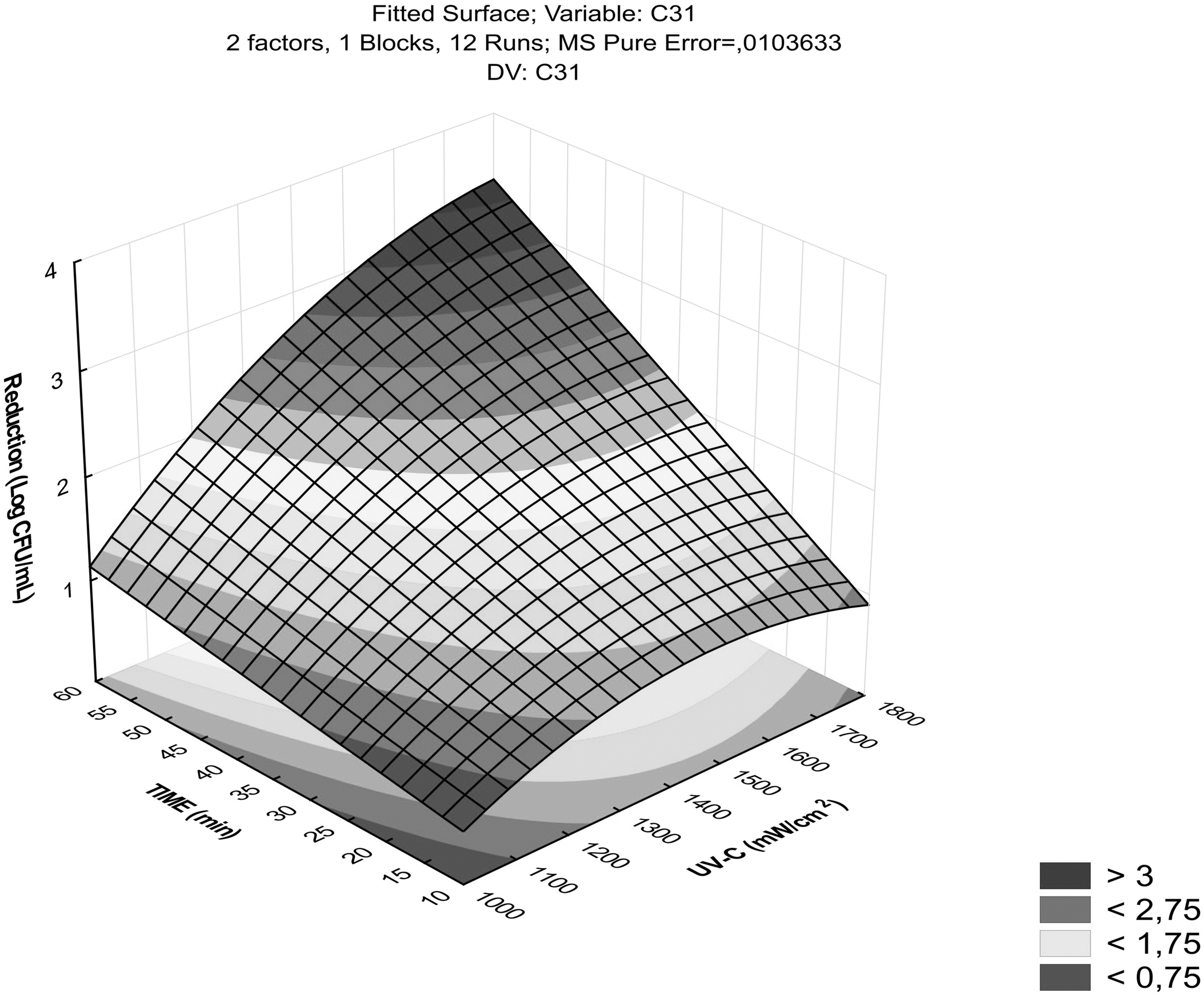
Logarithmic reduction of heat-resistant ExPEC from a CCRD for UVC-LED by combining intensities (mW/cm2) and times (min). The darker points indicate the greatest bacterial reduction, while the lighter points indicate the lowest reduction. CCRD, central composite rotatable designs; ExPEC, extraintestinal pathogenic Escherichia coli; UVC-LED, ultraviolet-C light-emitting diode.
In contrast, the CCRD model's performance for the US was inadequate since it showed a significant lack of fit. The inactivation effect for US was the same for all isolates (Table 2; p < 0.05), with a reduction of no more than 0.30 log CFU/mL. Even when this technology was combined with UVC-LED, the logarithmic reduction for two strains (C9 and C97) was lower (nonsynergic or antagonistic effect) than when only UVC-LED was used (p < 0.05). For strains C31 and C1145, UVC-LED and UVC-LED+US treatments showed the same result, which theoretically means that the combination of technologies was null.
Log Reduction of Heat-Resistant Extraintestinal Pathogenic Escherichia coli Strains After Treatments with Ultraviolet-C Light-Emitting Diodes, Ultrasound, and Their Combination
Mean log reduction (log10 CFU/mL) ± standard deviation. Capital letters in the same row indicate significant differences for the same strain in different treatments, and lowercase letters in the same column indicate significant differences between strains in relation to each treatment (p < 0.05).
CFU, colony-forming unit; US, ultrasound; UVC-LED, ultraviolet-C light-emitting diode.
Concerning genes related to UVC, we identified some in the genome of the isolates that could be involved in the survival of ExPEC in UVC environments (Fig. 2). Some of these genes, such as complex trxBC (which encodes for the thioredoxin [Trx] and Trx reductase system), and degP, are present in the tLST and are part of the module responsible for oxidative stress.
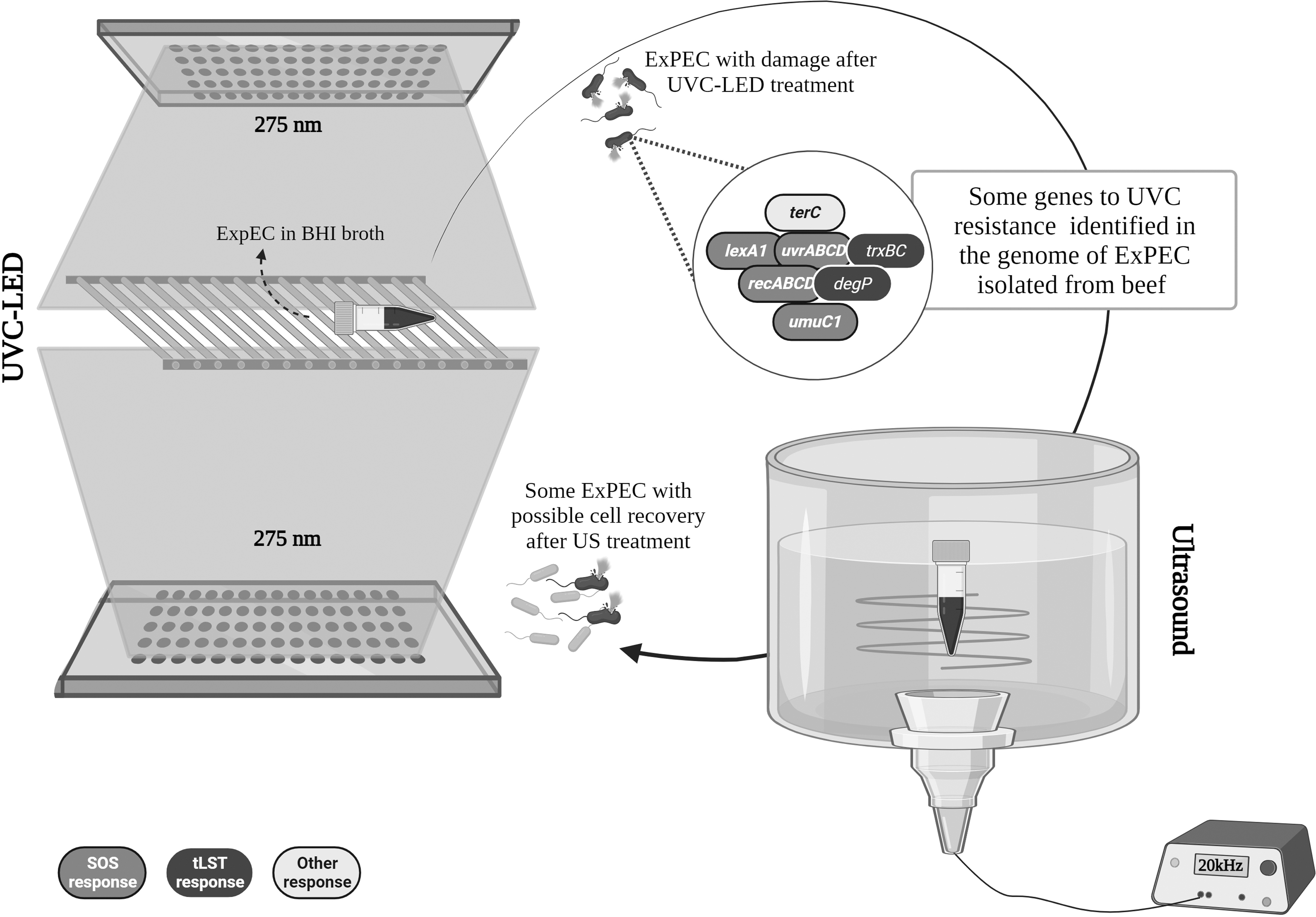
UVC-LED, US, and their combination on heat-resistant ExPEC isolated from beef, and the possible genes related to UVC resistance. ExPEC, extraintestinal pathogenic Escherichia coli; US, ultrasound; UVC-LED, ultraviolet-C light-emitting diode.
Discussion
Some studies have shown that US with frequencies between 20 and 70 kHz with intensities between 2 and 128 W/cm2 applied to lactic acid and pathogenic bacteria, including E. coli, can favor their viability due to the formation of temporary pores that favor the transport of nutrients and oxygen into the cells (Alves et al., 2018; Pitt and Ross, 2003). Again, when we evaluated the effect of US alone or combined with UVC-LED on heat-resistant ExPEC, the bacteria reduction rates were as low as those found in our previous study (Machado et al., 2023b). Therefore, under the conditions of time and intensity used, US does not prove to be an adequate strategy for reducing pathogens.
Bacteria that can overcome hostile environments (e.g., heat, acid, osmotic, and starvation shocks) can subsequently lead to cross-protection in the face of new survival challenges (Gayán et al., 2014; Mutz et al., 2020; van der Veen and Abee, 2011). In this sense, the acquisition of heat resistance genes in the isolates of this study due to selective pressure may also have corroborated with gene expression in the face of new challenges such as UVC. During the treatment with this technology, the CCRD showed that only the highest dose (4950 mJ/cm2 from 1.65 mW/cm2 for 50 min; Fig. 2) could reduce the highest bacteria load, although, after use, the strains showed some recovery.
Contrary to our findings, some other studies using UVC dosages between 1 and 4000 mJ/cm2 have proved to be efficient in reducing five or more logs of E. coli O157:H7 in a culture medium (Cheigh et al., 2012; Fan et al., 2021; Kim et al., 2017). Likewise, doses <4000 mJ/cm2 may eliminate some E. coli and other pathogens in meat products (Fan et al., 2021; Wang et al., 2023). Thus, the response to DNA damage can be more expressive when the strain has a history of resistance.
The damage caused by UVC is one of the main factors responsible for triggering an SOS response in E. coli that induces the expression of genes responsible for repairing DNA damage (Hamamoto et al., 2010). In total, we identified seven possible genes that could be expressed during or after UVC treatment, since a nonsynergistic effect (isolates C97 and C9) and null effect (C31 and C1145) were found after the combination of UVC-LED+US.
In E. coli, the SOS response has been globally studied since the 1970s, and, therefore, it is known that RecA and LexA proteins have some of the main functions of DNA repair in the SOS response pathway (D'Ari, 1985; Hamamoto et al., 2010). The genes involved in these proteins' expression include recA and lexA1 (mutant of the lexA gene) (Courcelle et al., 2001). Other genes responsible for repair, mutagenesis, excision, or recombination in the SOS response include the uvrABCD gene complex and the umuC and umuD genes (mutants umuC1 and umuD1) (D'Ari, 1985; Serment-Guerrero et al., 2020; Whitney and Weir, 2015). While the uvrABCD genes are involved in DNA strand damage correction, the umuCD genes form Pol V (Whitney and Weir, 2015).
Therefore, the genes involved in the SOS response are essential, especially those controlled by RecA. According to Whitney and Weir (2015), E. coli cells with RecA deficiency tend to have a lower lethal dose for mutagenic agents such as methyl methane sulfonate. Also, some plasmids encoded genes and prophages may be involved in SOS response (D'Ari, 1985). In this regard, we identified the terC gene in our strains (Machado et al., 2023a), which is part of ter operon that determines tellurite resistance. This operon was highlighted also showing low sensitivity to UVC (Jánošíková et al., 2021). In addition, the complex of genes trxBC responsible for expressing Trx protein for the tLST and degP expressing periplasmic protease were identified. Both of these proteins are part of the oxidative stress module in tLST (Kamal et al., 2021).
According to Mercer et al. (2017), trx genes have a low contribution to heat resistance, and degP does not have its contribution confirmed. However, these genes constitute important routes for expressing essential proteins during other oxidative stress (Krojer et al., 2008; Ritz et al., 2000). At this point, we highlight that the C97 strain presented these genes in its genome even with the absence of tLST (Machado et al., 2023a) and was the one that showed the lowest resistance to UVC-LED (p < 0.05). We believe that the absence of tLST in this strain did not influence the UVC-LED resistance phenotype since all other genes for oxidative stress were present.
The level of expression of these genes in each strain may have contributed to the different phenotypes observed, although future studies should be conducted in this regard. Thus, the expression of genes for UVC may have contributed to the recovery of damage, and the US contributed to the formation of pores that facilitated the transport of nutrients and cell replication.
Conclusion
In this study, US with intensity up to 299 W for 20 min was not able to reduce >0.30 log of ExPEC, but it could also be corroborating with the growth of this population when it was used after UVC-LED at a dose of 4950 mJ/cm2 (1.65 mW/cm2 for 50 min), showing a nonsynergistic effect. In this sense, by presenting a genotypic (presence of tLST) and phenotypic history of heat resistance, these strains may have responded to UVC stress more pronouncedly.
The probable expression of genes from the SOS system, and others such as trxBC and degP (present in tLST), and the terC gene (related to tellurite resistance), may have culminated in the recovery of the strains after UVC-LED treatment. Thus, the conditions analyzed in this study for US, UVC-LED, and their combination are ineffective in ExPEC populations with a certain history of heat resistance.
Footnotes
Authors' Contributions
Conceptualization of the study was contributed by M.A.M.M., V.S.C., M.L.G.M., Y.A.d.A.B., E.E.d.S.F., and C.A.C-.J. Data curation was done by M.A.M.M. and V.S.C. Formal analysis was carried out by M.A.M.M. and V.S.C. Funding acquisition was taken care by M.A.M.M., E.E.d.S.F., and C.A.C-.J. Investigation was by M.A.M.M. Methodology was done by M.A.M.M., V.S.C., M.L.G.M., Y.A.d.A.B., E.E.d.S.F., and C.A.C-.J.
Project administration was taken care by M.A.M.M., E.E.d.S.F., and C.A.C-.J. Resources were taken care by M.A.M.M., E.E.d.S.F., and C.A.C-.J. Software was carried out by M.A.M.M. and V.S.C. Supervision was done by E.E.d.S.F. and C.A.C-.J. Validation/visualization was carried out by M.A.M.M., V.S.C., M.L.G.M., Y.A.d.A.B., E.E.d.S.F., and C.A.C-.J. Writing—original draft was by M.A.M.M. Writing—review and editing was by M.A.M.M., V.S.C., M.L.G.M., Y.A.d.A.B., E.E.d.S.F., and C.A.C-.J.
Disclosure Statement
No competing financial interests exist.
Funding Information
We are grateful for the financial support provided by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)/Brazil Process codes 444465/2020-0, 310462/2018-5, 151336/2020-2, 313119/2020-1, 310181/2021-6, and 152241/2022-1, and the Fundação Carlos Chagas Filho de Amparo a Pesquisa do Estado do Rio de Janeiro—Brasil (FAPERJ), grant nos. E-26/200.891/2021 and E-26/204.145/2022.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.