
Abstract
Antimicrobial resistance (AMR) is a great threat to animal and public health. Here, we conducted a surveillance of Escherichia coli isolated from healthy chickens during 2009–2014 to identify the characteristics of AMR. A total of 351 (95.64%) E. coli isolates were obtained from 367 healthy chicken fecal samples collected from 6 farms located in Shandong Province, China. The susceptibility to 10 antimicrobials, the prevalence of antibiotic resistance genes (ARGs), phylogenetic clustering, and multilocus sequence typing were evaluated. The isolates exhibited high resistant rates (>95%) to ampicillin, cefotaxime, ciprofloxacin, ceftiofur, and enrofloxacin. The most prevalent ARGs were bla CTX-M (36.36%), aac(6′)-Ib-cr (30.79%), qnrS (29.62%), oqxAB (27%), mcr-1 (15.83%), bla TEM (9.09%), qnrC (3.52%), qnrD (0.88%), and qepA (0.29%). Phylogenetic clustering analysis indicated that the most prevalent group was group D (37.89%), followed by group B1 (34.76%), A (24.22%), and B2 (3.13%). Fifty-seven sequence types (STs) were identified among the 124 bla CTX-M-positive strains, and the dominant STs were ST354 (13.71%), ST117 (5.65%), ST155, ST2309, and ST2505 (4.84% each). There was a significant association between 17 pairs of AMR phenotypes, 14 pairs of ARGs, and 11 pairs of AMR-ARGs. The strongest association was found between ST602 and qnrC (odds ratios: 22.2). This study implied that E. coli isolated from healthy chickens could potentially serve as a reservoir of AMR and ARGs, and significant associations exist among AMR, ARGs, phylogenetic groups, and STs. Our study highlighted the need for routine surveillance of AMR in healthy chickens, and promoting appropriate antibiotic use and implementing regular monitoring of resistance in broilers are crucial for fostering the development of the poultry industry and safeguarding public health.
Introduction
E
Currently, antibiotics used to treat colibacillosis are mostly shared between veterinary and human medicine. This raises two concerns. First, there are significant genetic similarities among strains found in animals and humans, as previously described in broilers, their products, and humans (Dierikx et al., 2013; Huijbers et al., 2014). Second, AMR is a growing issue that may contribute to treatment failures (Poirel et al., 2018).
In the treatment of E. coli infection, β-lactamics and fluoroquinolones are commonly used drugs, while polymyxin E serves as a last-resort antibiotic for multidrug-resistant (MDR) bacterial infections. Since the first extended-spectrum β-lactamase (ESBL) was discovered in Germany in 1983, ESBL-producing E. coli (ESBL-EC) has rapidly spread globally, leading to a significant increase in the prevalence of ESBL-EC strains worldwide (Pitout and Laupland, 2008). The most prevalent ESBLs are CTX-M-type β-lactamases, and bla CTX-M genes are typically found on transferable plasmids that also carry resistance genes for other antimicrobials, such as fluoroquinolones and polymyxin E (Guo et al., 2021; Liu et al., 2020; Wu et al., 2018). This raises concerns regarding the potential dissemination of other resistance genes through coselection, resulting in the emergence of MDR strains and treatment failures (Haenni et al., 2016).
Several studies worldwide have studied the epidemiology and molecular characterization of commensal E. coli from poultry (Abraham et al., 2023; Goudarztalejerdi et al., 2020; Makarov et al., 2020; Much et al., 2019; Mudenda et al., 2023; Shaib et al., 2023), whereas similar reports in China are mainly concentrated in the following provinces: Shaanxi (Liu et al., 2020), Heilongjiang, Jilin, Liaoning (Tong et al., 2015), Henan (Yuan et al., 2009), Guangdong (Zheng et al., 2012), Jiangxi (Tan et al., 2023), Zhejiang (Ma et al., 2022), Shanghai (Lv et al., 2022), and Qinghai (Tang et al., 2022). Therefore, to better understand the characteristics of E. coli from healthy chickens in Shandong province, AMR, ARGs, phylogenetic groups, and sequence types (STs) were determined, and the associations among AMR, ARGs, phylogenetic groups, and STs were analyzed in this study.
This study offers a theoretical foundation for understanding the fundamental characteristics of E. coli in broilers, thereby contributing to future advancements in the prevention and treatment of colibacillosis.
Materials and Methods
Samples and bacterial isolation
From March 2009 to October 2014, broiler chicken fecal samples were randomly collected from 367 healthy chickens in 6 large broiler farms in Shandong Province, China, respectively (Fig. 1). All experiment activities in this study were approved by the Institutional Review Board of Shandong Academy of Agricultural Sciences. These six farms were chosen based on the two criteria: (1) large-scale breeding farms and (2) no disease occurred during the preselection and sampling period. The selected chickens appeared to be in good health; and each sample was obtained from an individual animal. Sampling must follow the principle of cluster random sampling. The samples were streaked onto MacConkey agar and incubated overnight. A single colony exhibiting typical E. coli morphology was selected; and the identification of E. coli was confirmed by polymerase chain reaction (PCR) amplification of phoA gene (Hu et al., 2011).

The sampling locations, the number of collected samples, and the isolated strains, and the prevalence of Escherichia coli in each farm. Sample sources were shaded in green.
Antimicrobial susceptibility testing
A total of 10 antimicrobial drugs were prepared in different concentrations for antimicrobial susceptibility testing, including penicillins (ampicillin [AMP, 1–512 μg/mL]), cephalosporins (ceftiofur [CEF, 0.0625–512 μg/mL], cefotaxime [CTX, 0.0625–512 μg/mL]), quinolones (ciprofloxacin [CIP, 0.25–512 μg/mL], enrofloxacin [ENR, 0.25–512 μg/mL], levofloxacin [LEV, 0.0625–512 μg/mL]), aminoglycosides (gentamicin [GEN, 0.25–512 μg/mL]), chloramphenicol (florfenicol [FFC, 1–512 μg/mL]), tetracycline (doxycycline [DOC, 0.5–512 μg/mL]), and peptide antibiotic (polymyxin B [PB, 0.0625–64 μg/mL]). Antimicrobial susceptibility testing was performed using the agar dilution method (CLSI, 2019). Susceptibility to colistin was interpreted according to EUCAST guidelines (http://www.eucast.org/clinical_breakpoints/). The breakpoints for the other tested drugs were as recommended by the CLSI (2019) and veterinary CLSI (VET01-A4) guidelines (CLSI, 2013). E. coli ATCC 25922 was used as a quality control in this study.
PCR amplification of antibiotic resistance genes (ARGs)
Isolates showing reduced susceptibility to CTX were subjected to PCR amplification to screen for the presence of bla CTX-M (including group-specific PCRs for bla CTX-M groups 1, 2, 8, 9, and 25), bla TEM, and bla SHV genes (Edelstein et al., 2003; Grimm et al., 2004; Grobner et al., 2009; Liu et al., 2007). The presence of plasmid-mediated quinolone resistance (PMQR) genes was determined by PCR amplification. The targeted genes include qnrA, qnrB, qnrC, qnrD, qnrS, aac(6′)-Ib-cr, qepA, and oqxAB (Cavaco et al., 2009; Gay et al., 2006; Park et al., 2006; Wang et al., 2009; Wang et al., 2004; Yamane et al., 2008; Zhao et al., 2010). Two multiplex PCR assays were utilized to detect colistin resistance genes: one for mcr-1–mcr-5 (Rebelo et al., 2018), and the other for mcr-6–mcr-9 (Borowiak et al., 2020). The PCR products were purified and sequenced by Sangon Biotech (Beijing, China).
Phylogenetic analysis
Phylogenetic groups were determined using a triplex PCR assay following a previously described classification method (Clermont et al., 2000).
Multilocus sequence typing
The bla CTX-M-positive isolates were characterized using multilocus sequence typing (MLST). For this purpose, PCR amplification was conducted on seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) using primers provided on the website (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli) (Wirth et al., 2006). MLST minimal spanning trees were constructed online using PHYLOVIZ.
Statistical analysis
Associations among AMR, ARGs, phylogenetic groups, and MLST sequence types (STs) were analyzed using Pearson's chi-square test (SPSS, version 25; IBM Corp., Armonk, NY). Statistical significance was set at p < 0.05. Meanwhile, odds ratios (OR) and 95% confidence intervals (CI) were calculated to determine statistically significant associations. An OR <1 indicates a negative association, while an OR >1 indicates a positive association.
Results
Antimicrobial susceptibility of E. coli isolates
A total of 351 strains of E. coli were isolated and identified from 367 samples, with an isolation rate of 95.64%. The detailed information of collection time, city, minimum inhibitory concentration of each strain were shown in Supplementary Table S1. Among 351 tested strains, the highest rate of resistance was observed for AMP (99.14%, n = 348), followed by CTX (97.15%, n = 341), CIP (95.44%, n = 335), CEF (95.16%, n = 334), ENR (95.16%, n = 334), FFC (94.02%, n = 330), and DOC (82.62%, n = 290). The resistance rates for GEN, LEV, and PB were lower at 73.50%, 63.53%, and 34.47%, respectively. Figure 2A illustrates the variation in the resistance rates of the antimicrobials across different farms, and the rate of AMR in different farms were significantly different. The E. coli strains in Farm 3 and Farm 6 had a higher rate of AMR than in other cities.
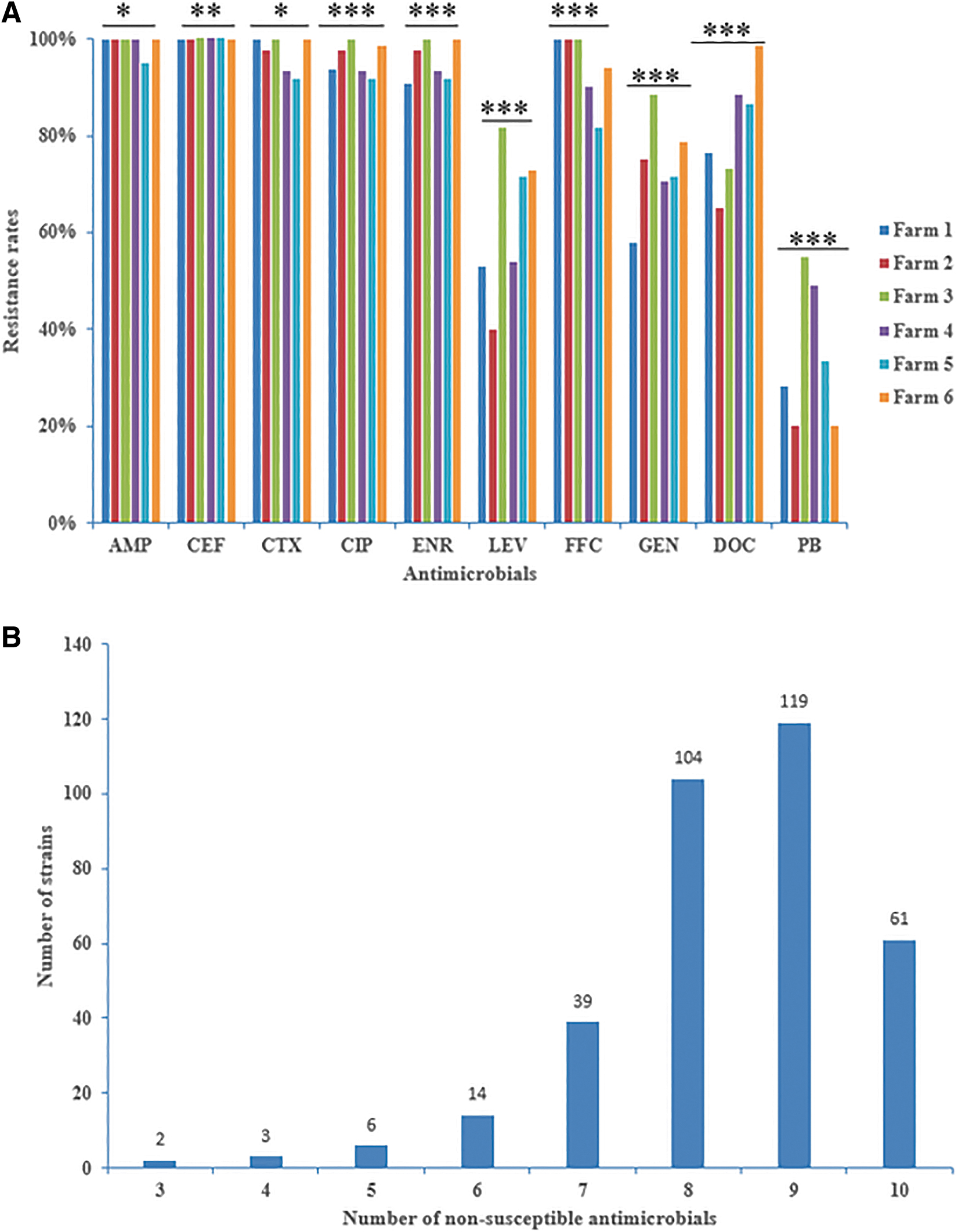
The antimicrobial resistance phenotypes of Escherichia coli isolates.
In addition, 348 (99.15%) were classified as MDR, exhibiting resistance to 3 or more classes of antimicrobial agents. Furthermore, 323 isolates (92.02%) were identified as having high-level MDR, displaying resistance to 7 or more classes of antimicrobials (Fig. 2B). The 350 resistant strains exhibited diverse AMR profiles. Among the 49 AMR phenotypes observed, the most common resistance phenotype was AMP-CEF-CTX-CIP-ENR-LEV-FFC-GEN-DOC (84, 24%), followed by AMP-CEF-CTX-CIP-ENR-LEV-FFC-GEN-DOC-PB (61, 17.43%), and AMP-CEF-CTX-CIP-ENR-FFC-GEN-DOC (38, 10.86%).
Characterization of ARGs
Among the 341 CTX-resistant isolates, 124 (36.36%) carried the bla CTX-M gene, 31 (9.09%) carried the bla TEM gene, and no bla SHV gene was detected. Among the 124 bla CTX-M-positive strains, 52 were positive for bla CTX-M-1G, 97 were positive for bla CTX-M-9G, and 26 strains carried both bla CTX-M-1G and bla CTX-M-9G genes. The sequencing results revealed five subtypes of bla CTX-M-1G and seven subtypes of bla CTX-M-9G, with bla CTX-M-65 (64, 51.61%) being the most prevalent subtype, followed by bla CTX-M-55 (32, 25.81%) and bla CTX-M-14 (24, 19.35%) (Fig. 3A). Notably, 12 isolates carried both bla CTX-M-1G and bla TEM, 23 isolates carried both bla CTX-M-9G and bla TEM, and 4 isolates carried all 3 genes (bla CTX-M-1G, bla CTX-M-9G, and bla TEM).
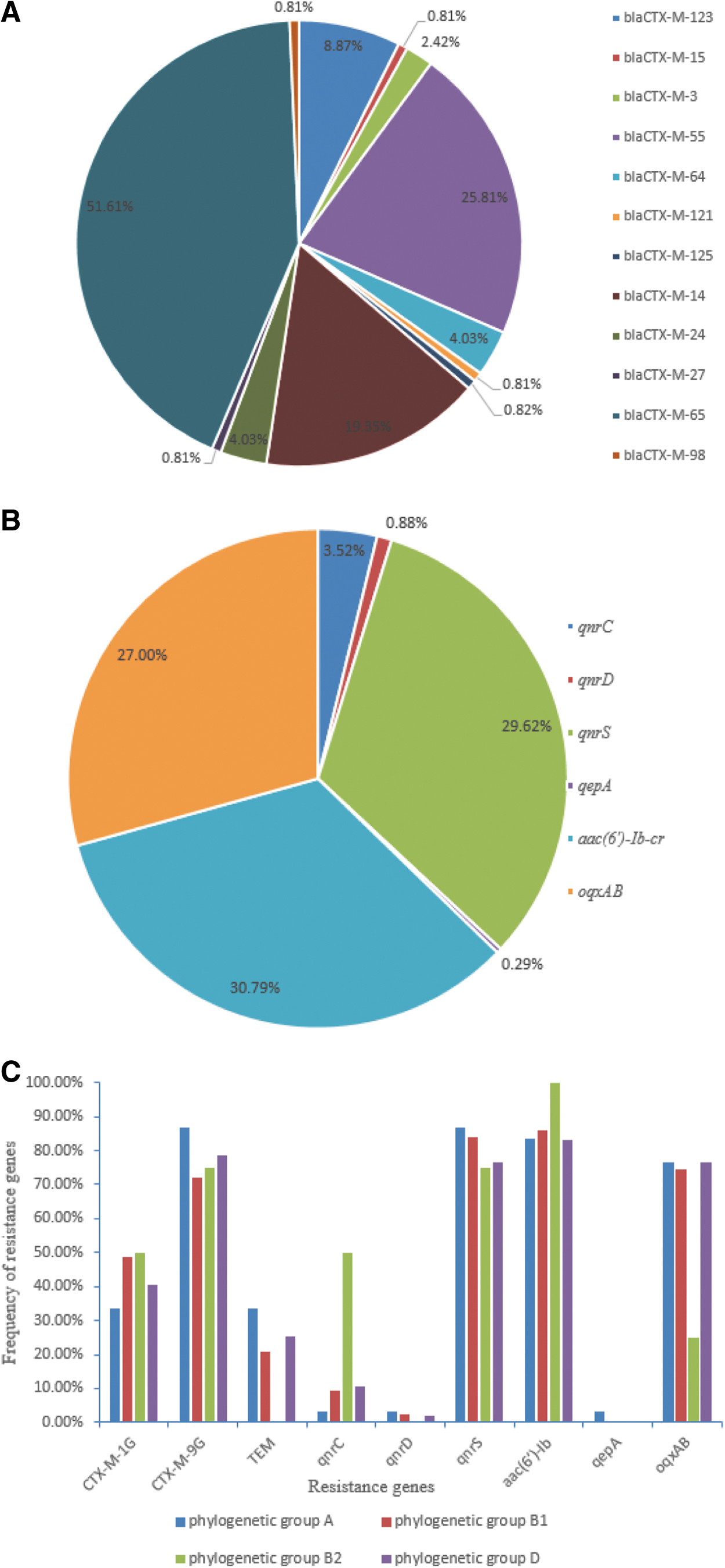
Distribution of ESBLs and PMQRs resistance genes.
Regarding PMQR genes among the 341 CTX-resistant isolates (Fig. 3B), aac(6′)-Ib-cr was the most prevalent (105; 30.79%), followed by qnrS (101; 29.62%), oqxAB (92; 27%), qnrC (12; 3.52%), qnrD (3; 0.88%), and qepA (1; 0.29%). None of the strains harbored qnrA and qnrB. Furthermore, all PMQR genes coexisted with ESBL genes; and 119 bla CTX-M-positive E. coli isolates (95.97%) carried at least one PMQR gene. The distribution of PMQR genes among ESBL-EC isolates is shown in Table 1. The most prevalent gene combination was bla CTX-M-9G+qnrS+aac(6′)-Ib-cr+oqxAB (26, 20.97%), followed by bla CTX-M-9G+bla TEM+qnrS+aac(6′)-Ib-cr+oqxAB (15, 12.10%), and bla CTX-M-1G+bla CTX-M-9G+qnrS+aac(6′)-Ib-cr+oqxAB (11, 8.87%).
Distribution of Plasmid-Mediated Quinolone Resistance Genes/Genotypes Among Extended-Spectrum β-Lactamase-Positive Escherichia coli Isolates
CTX, cefotaxime; ESBL, extended-spectrum β-lactamase; PMQR, plasmid-mediated quinolone resistance.
For PMCR detection, the multiplex PCR assay detected the mcr-1 (54, 15.83%) gene, but did not detect other mcr genes. Among the ESBL-EC isolates, mcr-1 was detected in 44 (35.48%) strains. Of the mcr-1-positive isolates, 22 (40.74%) were resistant to AMP-CEF-CTX-CIP-ENR-LEV-FFC-GEN-DOC-PB. All mcr-1-positive isolates carried at least one ESBL/PMQR gene, and 93.18% (41/44) had a combination of ESBL and PMQR determinants (Table 2).
The Information Regarding mcr-1-Positive Escherichia coli Isolates
AMP, ampicillin; CEF, ceftiofur; CIP, ciprofloxacin; CTX, cefotaxime; DOC, doxycycline; ENR, enrofloxacin; ESBL, extended-spectrum β-lactamase; FFC, florfenicol; GEN, gentamicin; LEV, levofloxacin; PB, polymyxin B; PMQR, plasmid-mediated quinolone resistance; ST, sequence type.
Phylogenetic grouping
Phylogenetic group D was the most frequent (133, 37.89%) among the isolates, followed by groups B1 (122, 34.76%), A (85, 24.22%), and B2 (11, 3.13%). The distributions of ESBL, PMQR, and mcr-1 genes varied among the different phylogenetic groups, as shown in Figure 3C.
Molecular typing of 124 blaCTX-M-harboring strains
A total of 57 STs were identified, with ST354 being the most predominant (17, 13.71%), followed by ST117 (7, 5.65%), ST155, ST2309, and ST2505 (6, 4.84% each). In addition, 36 strains had their own specific ST types, indicating a lack of widespread clonal transmission in this study (Fig. 4).
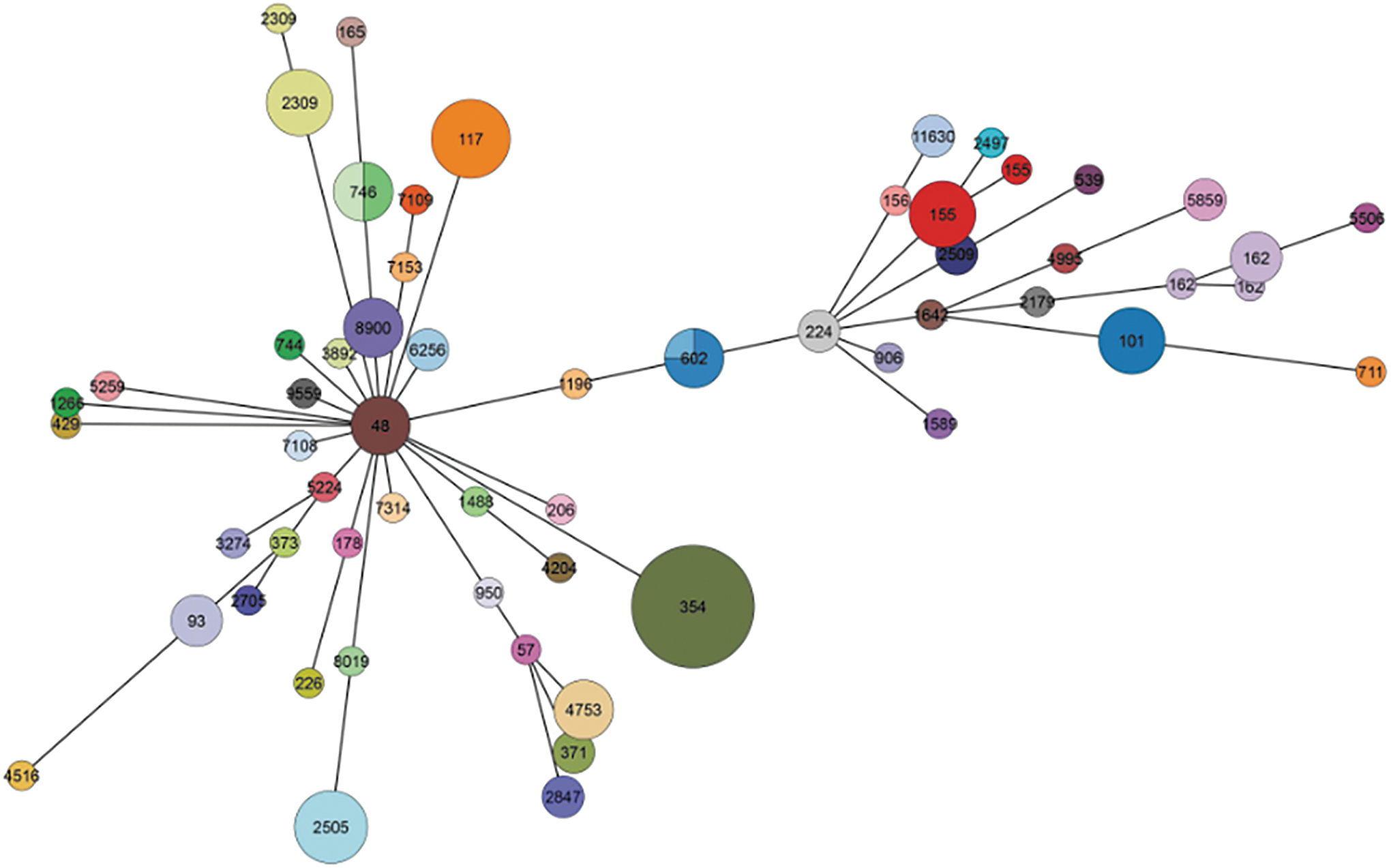
Clustering analysis based on the multilocus sequence genotyping assay results for the bla CTX-M-positive isolates using PHYLOVIZ online.
Association among AMR phenotypes
Resistance to one antibiotic frequently exhibits a certain correlation with resistance to other antibiotics. The statistical analysis showed that resistance to AMP was positively associated with resistance to ENR (OR: 44.4; 95% CI: 3.810–517.396; p = 0.000) (Supplementary Table S2). Positive associations were also found in other 16 pairs of AMR phenotypes, namely: CEF-CTX, CEF-CIP, CEF-ENR, CEF-GEN, CEF-DOC, CTX-CIP, CTX-ENR, CTX-LEV, CTX-GEN, CIP-ENR, CIP-LEV, CIP-GEN, ENR-FFC, ENR-GEN, LEV-FFC, and LEV-GEN.
Association among ARGs
Presence of bla TEM gene was found to be positively associated with the qnrD gene (OR: 22; 95% CI: 1.936–249.88; p = 0.021) (Supplementary Table S3). Positive associations were also observed between gene pairs of bla TEM-qnrS, bla TEM-aac(6′)-Ib-cr, bla TEM-oqxAB, bla TEM-mcr-1, qnrC-aac(6′)-Ib-cr, qnrC-oqxAB, qnrC-qnrS, qnrS-aac(6′)-Ib-cr, qnrS-oqxAB, qnrS-mcr-1, aac(6′)-Ib-cr-oqxAB, aac(6′)-Ib-mcr-1, oqxAB-mcr-1.
Association between AMR and ARGs
Both positive (OR >1) and negative (OR <1) associations between AMR and ARGs were observed in the present study. Resistance of PB was found to be positively associated with the presence of bla CTX-M gene (OR: 8.806; 95% CI: 5.324–14.568; p = 0.000) (Supplementary Table S4). Positive associations were also observed between pairs of PB-bla CTX-M-1G, PB-bla CTX-M-9G, PB-bla TEM, PB-qnrC, PB-qnrS, PB-aac(6′)-Ib-cr, and PB-oqxAB. On the contrary, significant negative associations were also observed between FFC-bla CTX-M-1G, ENR-bla CTX-M-1G, and ENR-mcr-1.
Association of STs with AMR, ARGs, and phylogenetic groups
Correlation analysis demonstrated a negative association between ST101 and resistance to DOC (OR: 0.119; 95% CI: 0.019–0.762; p = 0.009) (Supplementary Table S5). Similar negative associations were also observed between ST2309-FFC, ST2309-ENR, ST2509-CTX, ST162-oqxAB, ST2309-qnrS, and ST117-bla CTX-M-9G. On the contrary, positive significant correlation was found between ST602-qnrC, ST117-group A, and ST155-group B1.
Discussion
In recent years, AMR in veterinary settings has gained global attention owing to its significant impact on human health and increasing reports of animal-to-human AMR transmission. In this study, an MDR phenotype was observed in 99.15% of the isolates, which was higher than the rates reported in other countries, including Korea (98.5%) (Park et al., 2019) and Tanzania (69.3%) (Mgaya et al., 2021). AMR has emerged as a result of selective pressure and exposure to antibiotics. Moreover, this study identified positive significant correlations among 17 pairs of AMR phenotypes. These findings indicate that the use of a particular antimicrobial agent not only leads to resistance against specific drugs but also promotes the development of resistance to other antimicrobial agents, including unrelated drugs.
Previous studies have indicated that bla TEM is the dominant ESBL gene in chicken-derived E. coli isolates (Tohmaz et al., 2022), whereas others indicate that bla CTX-M, particularly bla CTX-M-55, is prevalent, with an increasing trend in recent years (Liu et al., 2021; Shafiq et al., 2021; Wu et al., 2018). In our study, we observed that chicken-derived E. coli primarily produced bla CTX-M-65 enzyme. The occurrence of bla CTX-M-65 is relatively rare in ESBL-EC. Previous reports have noted the dominance of bla CTX-M-65 in E. coli isolated from chickens (Yuan et al., 2009), diseased pets (Chen et al., 2019), and diseased farmed pigeons (Yang et al., 2015) in China, raw retail chickens in South Korea (Park et al., 2019), and healthy children in the Bolivian Chaco region (Riccobono et al., 2015). PMQR genes have been reported worldwide and are frequently found in ESBL-EC (Xu et al., 2015).
In our study, PMQR genes were present in 95.97% of ESBL-EC, with aac(6′)-Ib-cr being the most prevalent, either alone or in combination with other PMQR genes. Moreover, our study revealed that 85.71% of the strains harbored two or more PMQR genes, indicating a higher prevalence than that reported in previous studies (Xu et al., 2015). Interestingly, the detection rates of ARGs did not necessarily align with the corresponding resistance phenotype in our study. As widely recognized, the antibiotic resistance profile of bacteria is determined by the associated ARGs. However, it has been observed that there is no absolute correlation between resistance phenotypes and the detection rates of resistance genes (Fan et al., 2022; Zhang et al., 2021). This discrepancy could be attributed to several factors.
First, the expression of ARGs represents only one of the mechanisms through which bacterial resistance can occur. Other mechanisms, such as target modification or efflux pump activity, might also contribute to resistance without being dependent on gene expression. Second, the abnormal or low-level expression of ARGs may not be sufficient to confer drug resistance in bacteria (James et al., 2021; Munita and Arias, 2016). These observations indicate that the relationship between ARG detection and resistance phenotype is complex and requires further investigation.
Colistin (polymyxin E) has been widely recognized as a last-resort treatment option for specific MDR bacterial infections. In our study, positive correlations were observed between PB resistance and the presence of eight ARGs, and similar positive correlations were found between mcr-1 and the other four ARGs. These ARGs are present in gram-negative bacteria and are prevalent worldwide. It remains uncertain whether these significant correlations promote polymyxin resistance and the spread of mcr-1 gene. Further research is needed to investigate this possibility.
AMR, ARGs, virulence genes, phylogenetic groups, and STs are important characteristics of bacteria that can provide valuable information for understanding epidemiological characteristics. Previous studies have shown significant correlations among AMR, ARGs, virulence genes, and phylogenetic groups (Fan et al., 2022; Rahman et al., 2022; Rybak et al., 2022). However, few studies have investigated the correlation between ST and the other four factors, and only a few ST type have been found to have significant correlations with virulence genes (Kromann et al., 2023) or some phylogenetic groups (Suresh et al., 2021) in E. coli. In our study, statistical analysis showed that there were significant correlations between STs and the other three factors (AMR, ARGs, and phylogenetic group), with seven negative significant correlations and three positive significant correlations.
ST101 has previously been linked to antimicrobial-resistant genes, such as ESBL and AmpC (Lee et al., 2021; Truong et al., 2021). In addition, when compared to other ST groups, ST101 showed a significant overrepresentation of 12 virulence genes (Kromann et al., 2023). Statistical analysis in our study demonstrated a significant association between ST101 and susceptibility to DOC. However, since ST101 has been reported in both humans and animals, it is likely that they would encounter tetracyclines. If tetracycline resistance does not result in a loss of fitness in E. coli, one would expect its frequency to increase over time. This is supported by the high prevalence of ST101 within Danish broiler production, further highlighting the potential rise of tetracycline resistance. Although this conclusion is not necessarily general due to the small number of experimental data, it can at least prove that there are significant correlations between these factors.
In future, studies can be given to investigating the correlation between ST and other factors to increase knowledge about the transmission of bacteria of the same ST type between different species.
In conclusion, E. coli isolates from chickens exhibited a very high rate of MDR strains. Among the bla CTX-M-positive isolates, 95.97% harbored at least one PMQR gene, and all mcr-1-positive isolates carried at least one ESBL/PMQR gene. In addition, significant associations were identified among AMR, ARGs, STs, and phylogenetic groups. Antibiotic-resistant E. coli strains derived from chickens pose a potential threat, emphasizing the crucial need for the ongoing monitoring of E. coli resistance patterns in chicken populations.
Footnotes
Disclosure Statement
No competing financial interests exist.
Authors' Contributions
L.L.L., X.N.Z., and Y.Q.L. designed the experiments; Q.Z. and Y.B.L. completed the sample collection and identification; L.L.L. carried out the experiments and wrote the article; G.L.G. contributed the figure drawing; M.H. and Y.B.C. revised the article.
Funding Information
This work was supported by the National Key Research and Development Program of China (2019YFA0904002), and Jinan City “new university 20” funding project (202228001).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.