
Abstract
Staphylococcus aureus causes various toxigenic and invasive diseases in humans worldwide. This study examined the prevalence, virulence genes, and antibiotic resistance of S. aureus isolates collected from 894 retail food samples in Ardabil, Iran. Staphylococcal cassette chromosome mec (SCCmec), spa, and multilocus sequence typing methods were employed to further investigate the molecular characteristics of methicillin-resistant S. aureus (MRSA) isolates. The results revealed that 11.18% (n = 100) of food samples exhibited contamination with S. aureus (10.50% methicillin-sensitive S. aureus [MSSA] and 0.67% MRSA). Notably, raw minced meat (29.41%), Faloodeh (25%), and Olivier salad (21.42%) emerged as the most frequently contaminated food items. Among the 100 isolates of S. aureus, 94% were characterized as MSSA, with the remaining 6% identified as MRSA. The highest resistance was observed for penicillin (12%). MRSA isolates exhibited significantly higher resistance rates. Seventy-nine percent of the isolates were positive for sea, 14% for seb, 8% for a sec, and 0% for sed enterotoxin-encoding genes. Sixteen percent of isolates harbored two or more staphylococcal enterotoxin genes, simultaneously. Moreover, 97%, 94%, 24%, and 22% of isolates were positive for hla, hld, tst, and pvl virulence-encoding genes, respectively. No isolate was positive for the exfoliative toxins encoding eta and etb genes. MRSA isolates belonged to CC8 (n = 4) and CC22 (n = 2). Isolates in CC8 belonged to lineage ST239-MRSA-III and spa type t030; the isolates in CC22 belonged to ST22-MRSA-IV and spa types t310 and t223. In conclusion, a relatively high proportion of our retail food samples were contaminated with S. aureus. The high incidence of isolates with toxigenic genes raises serious health concerns. Furthermore, the presence of MRSA lineages linked to humans suggests that retail foods may be contaminated with human origin.
Introduction
S
Moreover, S. aureus easily contaminates various foods during preparation and handling, making it a common foodborne pathogen (Kadariya et al., 2014). This organism is the third cause of food poisoning in Europe, and each year, 241,000 cases of staphylococcal food poisoning (SFP) are reported in the United States (Merda et al., 2020; Scallan et al., 2011). However, it is thought that the actual incidence of SFP is underestimated because most cases are often not recorded.
It has been documented that only 10% of SPF patients seek medical care (Holmberg and Blake, 1984). S. aureus produces about 27 types of enterotoxins (SEs) and enterotoxin-like (SEls) proteins in contaminated foods. Food poisoning occurs following the ingestion of contaminated foods (Lefebvre et al., 2022). SEA, SEB, SEC, SED, and SEE are the common toxins responsible for 95% of SFP cases worldwide (Abril et al., 2020). Generally, the symptoms include severe vomiting, nausea, diarrhea, and abdominal pains. The occurrence of symptoms depends on the individual's vulnerability to toxins and the amount of intoxication (Hennekinne et al., 2012). SFP is usually a self-limiting disease, with a mortality rate of 77 in 1 million (Bokaie et al., 2020).
S. aureus produces two other toxins, exfoliative toxins and toxic shock syndrome toxin-1 (TSST-1), which are responsible for staphylococcal scalded skin syndrome and toxic shock syndrome, respectively (Lowy, 1998). In addition, S. aureus produces several other enzymes and cytolysins that contribute to the development of invasive staphylococcal infections in humans and animals (Ahmad Mansour et al., 2021).
S. aureus isolates commonly exhibit antibiotic resistance, and methicillin-resistant S. aureus (MRSA) is considered a significant threat to human and animal health (Aires de Sousa, 2017). Methicillin resistance is mediated by the production of a modified penicillin-binding protein (PBP2a) that is encoded by mecA or mecC genes. These genes are carried by the staphylococcal cassette chromosome mec (SCCmec). To date, 15 SCCmec elements (SCCmec I to SCCmec XV) have been identified (Nikolic and Mudgil, 2023). From an epidemiological perspective, MRSA is classified into three distinct types: Health care-associated MRSA (HA-MRSA), community-associated MRSA (CA-MRSA), and livestock-associated MRSA (LA-MRSA) (Stefani et al., 2012).
HA-MRSA isolates mainly carry SCCmec types I, II, and III; CA-MRSA isolates are generally associated with SCCmec types IV, V, and VI; and LA-MRSA can contain any of the SCCmec types associated with CA-MRSA or HA-MRSA (Boswishi et al., 2020). MRSA types belong to distinguished clonal complexes (CCs), determined by the multilocus sequence typing (MLST) approach (Turner et al., 2019). To date, MRSA isolates are frequently isolated from foods intended for human consumption. Enterotoxigenic MRSA isolates are usually associated with SPF, with similar clinical significance compared to methicillin-sensitive S. aureus (MSSA) counterparts (Sergelidis and Angelidis, 2017). However, some studies have reported MRSA-initiated invasive infections by ingesting contaminated foods (Kluytmans, 2010). In Iran, detailed studies of MRSA isolates in foods are scarce.
In this study, we examined the prevalence of MSSA and MRSA isolates in large-scale retail food samples received by the food control laboratory of Ardabil University of Medical Sciences in northwest Iran. We characterized the isolates' antimicrobial resistance patterns and genes that encode toxins. To better understand the epidemiological characteristics of MRSA isolates associated with food, we also identified molecular features such as CCs, sequence types (STs), SCCmec types, and spa types.
Materials and Methods
Bacterial isolation and identification
Between December 2019 and April 2021, a total of 894 retail food samples were admitted to the food control laboratory of Ardabil University of Medical Sciences. The samples were collected from 12 cities in Ardabil province, northwest Iran (Supplementary Fig. S1), and comprised raw meat (n = 232), dairy products (n = 180), pastries (n = 128), beverages and frozen desserts (n = 123), cooked protein foods (n = 159), salads (n = 42), and other foods (n = 30). S. aureus strains were isolated according to Iran's national standard number 6806 (ISIRI, 2006).
Briefly, 1 g of solid food sample was dissolved in 10 mL sterile Ringer's solution, then 1 mL of that or 1 mL of liquid foods was cultured onto plates of Baird-Parker agar (Biolife, Milan, Italy) and incubated at 37°C for 24 to 48 h. Coagulase-positive Staphylococcus colonies were identified by their ability to reduce tellurite to tellurium and lecithinase activity (ISIRI, 2006). From each food sample processed, one presumptive S. aureus colony was transferred onto Brain Heart Infusion agar (BioMaxima, Lublin, Poland) to provide a nurturing environment for growth.
The colony was then initially characterized by conventional microbiological tests such as catalase, gram stain, DNase (Merck, Darmstadt, Germany), and tube coagulase tests (Mahon and Lehman, 2023). The isolates were further characterized using polymerase chain reaction (PCR) by targeting the partial sequence of the nuc gene (a gene encoding the thermostable DNase of S. aureus) as described previously (Omid et al., 2023; Park et al., 2007; Ranjbar Omid et al., 2021). The PCR cycling and thermal conditions are described in Supplementary Table S1.
The isolates were stored in 1.5 mL cryotubes containing Tryptic Soy Broth (Merck) and 20% glycerol (Kimia Tehran acid, Tehran, Iran) at a temperature of −80°C.
Antibiotic susceptibility testing
Antimicrobial susceptibility analysis was performed using the disk diffusion approach on Muller-Hinton agar (BioMaxima) according to the Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2023). Nine antibiotic disks (Padtan Teb, Ghods, Iran) were tested, including penicillin (10 U), amoxicillin (20 μg), chloramphenicol (30 μg), tetracycline (30 μg), imipenem (10 μg), ciprofloxacin (5 μg), ceftriaxone (30 μg), cefazolin (30 μg), and gentamicin (10 μg). S. aureus ATCC 25923 was used for quality control. The inhibition zone diameters were measured and scored as resistant (R), intermediate-resistant (I), and susceptible (S) according to CLSI guidelines (CLSI, 2023). S. aureus isolates resistant to ≥3 antibiotics of different classes were reported as multidrug resistant (MDR).
MRSA detection
Methicillin resistance was detected using a cefoxitin disk (Fox, 30 μg) according to the protocols described by CLSI guidelines (CLSI, 2023). Isolates with an inhibition zone diameter of ≤21 mm were considered MRSA (CLSI, 2023). The mecA gene, which is responsible for methicillin resistance, was detected by PCR testing using primer sequences and conditions demonstrated in Supplementary Table S1 (Lee, 2003).
Screening of toxin- and cytolysin-encoding genes
PCR assay with specific primers (Supplementary Table S1) was used to detect enterotoxin (sea, seb, sec, and sed), exfoliative toxin (eta and etb), and toxic shock syndrome toxin-1 (tst)-encoding genes, as well as the cytolysins Panton–Valentine leukocidin (pvl)- and alpha hemolysin (hla and hld)-encoding genes, according to previous reports (Mkhize et al., 2021; Ranjbar Omid et al., 2021; Sharma et al., 2017). The PCR cycling and thermal conditions are described in Supplementary Table S1. DNA from previously identified S. aureus isolates with known virulence genes was used as positive controls.
Molecular typing
The MRSA isolates were subjected to SCCmec, S. aureus protein A (spa), and MLST.
SCCmec typing
SCCmec types were identified in MRSA isolates, according to previous reports (McClure-Warnier et al., 2013; Ranjbar Omid et al., 2021). The primer sequences and PCR conditions are listed in Supplementary Table S1. DNA from previously identified S. aureus isolates with known SCCmec types was used as a positive control (Omid et al., 2023).
spa Typing
The spa gene X region of MRSA isolates was amplified by PCR described previously (Omid et al., 2023). The primer sequences and PCR conditions are listed in Supplementary Table S1. Both strands of the amplicons were sequenced (Microsynth, Switzerland), and the spa types were identified using the Ridom StaphType software available online at www.spaserver.ridom.de
Multilocus sequence typing
MLST was performed on MRSA isolates using PCR amplification and sequencing of seven housekeeping loci: arcC (carbamate kinase), aroE (shikimate dehydrogenase), glpF (glycerol kinase), gmk (guanylate kinase), pta (phosphate acetyltransferase), tpi (triosephosphate isomerase), and yqil (acetyl coenzyme A acetyltransferase) according to Enright's protocol (Enright et al., 2000). STs were assigned using the online database available on the PubMLST website (www.Pubmlst.org).
Statistical analyses
Statistical analyses were performed using SPSS software v.11.5 (SPSS, Inc., Chicago), and the Chi-square test was applied to measure statistical significance. A p-value <0.05 was considered statistically significant.
Results
Of the 894 samples analyzed, 11.18% (n = 100) were positive for S. aureus (Table 1). Raw minced meat, Faloodeh, and Olivier salad were contaminated by S. aureus in a percentage of 29.41, 25, and 21.42, respectively, which were the most contaminated. Overall, 100 strains of S. aureus were collected from food samples, with 94 (94%) being MSSA and 6 (6%) being MRSA isolates. The MRSA isolates were from ice cream, Faloodeh, creamy pastry, lettuce salad, cooked protein food, and traditional cheese samples (Table 1).
Distribution of Staphylococcus aureus Isolates in Retail Food samples in Ardabil, Iran
MRSA, methicillin-resistant S. aureus; MSSA, methicillin-sensitive S. aureus.
Collectively, 79% of the isolates were positive for the sea enterotoxin-encoding gene, while 14% and 8% were positive for the seb and sec enterotoxin-encoding genes, respectively. The gene encoding for SED was not detected in any of the isolates (Fig. 1). In our study, 82% of isolates were positive for at least one SE-encoding gene. Six profiles of enterotoxin-encoding genes were identified overall, and 16% of isolates harbored two or more genes, simultaneously (Fig. 2).
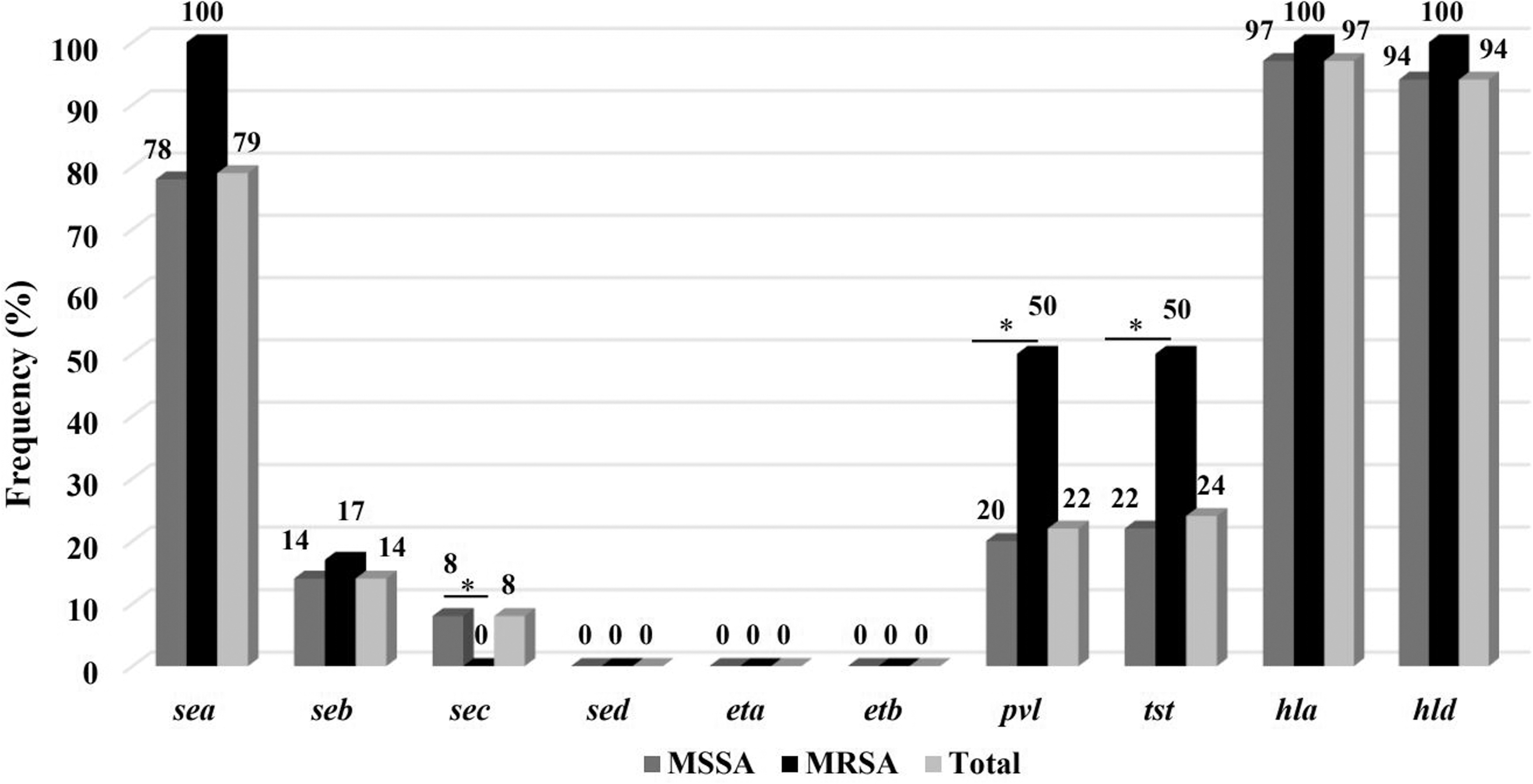
Frequency of enterotoxin and other toxin-encoding genes in Staphylococcus aureus isolates collected from retail food samples in Ardabil, Iran. *p ≤ 0.05.
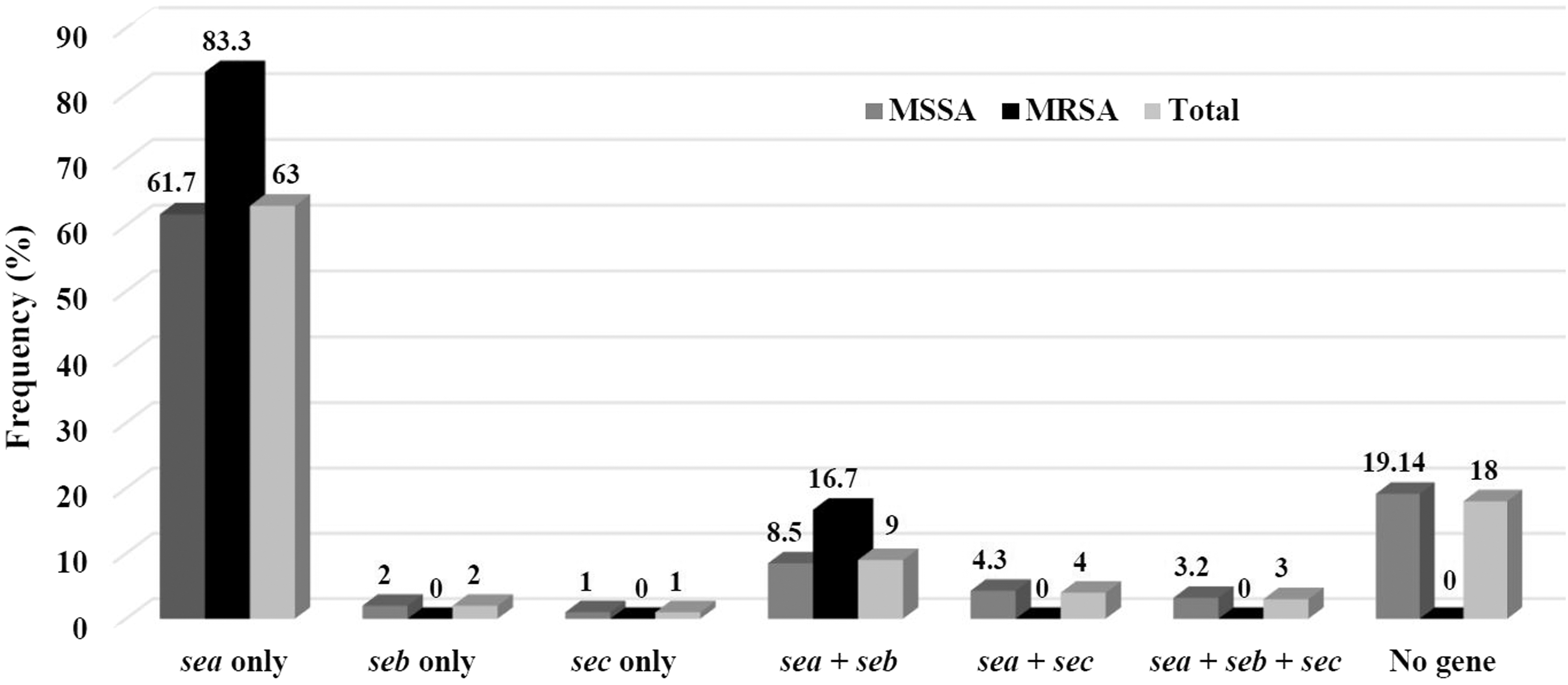
Combination pattern of enterotoxin-encoding genes among Staphylococcus aureus isolates collected from retail food samples in Ardabil, Iran.
Regarding other toxins, 24% and 22% of isolates were positive for tst and pvl genes, respectively. Moreover, 97% and 94% of isolates were positive for hla and hld, hemolysin-encoding genes, respectively. No isolate contained genes encoding exfoliative toxin eta and etb (Fig. 1). The relative abundance of tst and pvl genes was significantly higher in MRSA isolates (Fig. 1).
The antimicrobial resistance pattern of MSSA and MRSA isolates is shown in Figure 3. The majority of S. aureus isolates was susceptible to tested antibiotics, with the most resistance observed for penicillin (12%). The resistance rates of MRSA isolates were significantly higher compared to MSSA isolates.

Antibiotic resistance pattern of Staphylococcus aureus isolates collected from food samples in Ardabil, Iran. *p ≤ 0.05.
All six MRSA isolates examined were positive for the mecA gene. Four of these isolates belonged to SCCmec type III, while two belonged to SCCmec type IVa. According to MLST analysis, MRSA isolates belonged to two distinct CCs. Four isolates belonged to CC8 ST239, positive for SCCmec type III, and spa type t030. Two isolates belonged to CC22 ST22, positive for SCCmec type IVa, and spa type t310 and t223 (Table 2). The most frequently identified clone was the CC8-ST239-MRSA-III-t030 (n = 4).
Molecular Characteristics of Methicillin-Resistant Staphylococcus aureus Isolates Collected from Retail Food Samples in Ardabil, Iran
MRSA, methicillin-resistant S. aureus; SCCmec, staphylococcal cassette chromosome mec.
Discussion
In this study, 11.18% of samples from various foodstuffs were contaminated with S. aureus. The proportion of food contamination with S. aureus reported in some previous studies varies from 3.6% to 36.8%, depending on the sampling site (Oh et al., 2007; Wang et al., 2019; Wu et al., 2019; Xing et al., 2014). In previous studies performed in Iran, the contamination rate of S. aureus in food samples was between 1.10% and 52%, which is partially consistent with this study (Dehkordi et al., 2019).
The overall proportion of contamination with S. aureus in our study is located in the lower parts of the aforementioned reported ranges. However, S. aureus was observed in all food kinds, and the ratio was high in some food categories. This indicates the need to improve good hygiene practices in the food sector in Iran. The most frequently contaminated foods were raw minced meat (29.41%), faloodeh (25%) (an Iranian frozen dessert), and Olivier salad (21.42%). It is worth noting that the mentioned foods are among the most frequently contaminated with S. aureus (Murray et al., 2016).
The MRSA contamination rate in our food samples was 0.67%, which is similar to the rates reported from China (0.7% and 7.4%) (Wu et al., 2019; Zhao et al., 2021), Egypt (2.6%) (Mahros et al., 2021), Taiwan (3.6%) (Wang et al., 2019), and Malaysia (8%) (Seow et al., 2021). However, some studies conducted in Germany (37.2%) (Feßler et al., 2011), Bangladesh (26%) (Islam et al., 2019), and the Czech Republic (35.4%) (Tegegne et al., 2021) have reported higher prevalence of MRSA contamination in foodstuffs.
Compared to studies conducted in Iran, the prevalence of MRSA isolates in our study remains under the average prevalence of 15.5% reported in contaminated foods (Dehkordi et al., 2019). The differences in the incidence of contamination with MSSA and MRSA strains among different studies may be attributed to factors such as the sample size, sampling technique, sample type, and sensitivity of the isolation and identification methods (Şanlıbaba, 2022; Wu et al., 2019). MRSA isolates have been shown to belong to specific lineages with different microbiological and epidemiological characteristics based on the origin of the isolates (Doulgeraki et al., 2017).
The six MRSA isolates detected in our study belonged to CC8 and CC22. The isolates in the CC8 group composed a single ST239-MRSA-III linage with an identical spa type (t030). It is one of the most common pandemic HA-MRSA clones, which is frequently isolated in Saudi Arabia, Iran, and Turkey (Cîrlan et al., 2005; Fatholahzadeh et al., 2009; Oksuz et al., 2013). The isolates in the CC22 group belonged to linage ST22-MRSA-IV, which is also a pandemic MRSA clone responsible for both hospital- and community-acquired infections, as well as associated with domestic animals (Monecke et al., 2011; Oksuz et al., 2013).
In this study, the ST22-MRSA-IV isolates distributed in spa types t223 and t310, previously described as CA-MRSA and HA-MRSA, respectively (Dai et al., 2019; Gostev et al., 2017; Tabaja et al., 2021). The occurrence of these clonal lineages in food samples suggests that humans contribute largely to food contamination with MRSA, which is likely transferred to food by workers at any processing stage. However, S. aureus food contamination can occur through the transmission of bacteria from infected/colonized humans/animals and cross-contamination through unsanitary environments (Kluytmans, 2010).
This study found that a very small proportion of S. aureus isolates were resistant to antibiotics. This differs from the results reported in Iran and other countries, which showed significant rates of antibiotic resistance in food-related S. aureus isolates (Arefi et al., 2014; Wang et al., 2017). Similar to a study from Turkey, resistance to penicillin was more frequently observed than other antibiotics (Şanlıbaba, 2022). The difference in resistance rates reported in different studies can be attributed to the burden of antibiotic consumption in humans and animals in a particular region (Meyer et al., 2013).
In our study, 82% of isolates were positive for at least one SE-encoding gene, which is higher than the rates reported from Turkey (65.62%) (Şanlıbaba, 2022) but lower than that found in China (99.07%) (Wu et al., 2019). Furthermore, among our isolates, the detection rate for sea, seb, and sec genes was 79%, 14%, and 8%, respectively. These findings are consistent with several studies indicating a higher prevalence of sea, seb, and sec genes in the S. aureus isolates (Dehkordi et al., 2019; Oh et al., 2007; Song et al., 2015; Wang et al., 2017; Wu et al., 2019). In our study, sed was not detected, which is inconsistent with a study from China, in which sed was reported as the most frequent SE-encoding genes in S. aureus isolates in contaminated foods (Xing et al., 2014).
In this study, a notably high prevalence of the tst gene was found in S. aureus isolates, which is in contrast to the reports of Song et al. (2015) from China, and Oh et al. (2007) from Korea detected the tst gene in 0% and 4.6% of S. aureus isolated from food, respectively. tst encodes for TSST-1 responsible for the genesis of staphylococcal toxic shock syndrome (Ahmad Mansour et al., 2021). TSST-1 has not been known medically as a toxin that causes foodborne disease (Ahmad Mansour et al., 2021). Recently, Goudsmit et al. (2021) reported a toxic shock syndrome case due to a TSS1-producing S. aureus acquired by ingesting contaminated food in Belgium. This indicates the importance of detecting the tst gene in isolates from food samples.
PVL-encoding genes were found in 22% of S. aureus isolates. A similar finding was reported in a study conducted in China, with 21% of isolates positive for pvl (Song et al., 2015). In contrast, in research conducted by Tegegne et al. (2021), no pvl-positive S. aureus isolate was detected in meat samples. PVL is associated with severe staphylococcal pneumonia and soft tissue infections (Morgan, 2007).
Staphylococcal exfoliative toxin-encoding genes eta and etb were not detected in our isolates. Other studies also have reported similar results, indicating that S. aureus food isolates have a low prevalence of exfoliative toxin-encoding genes (Song et al., 2015; Tegegne et al., 2021). This toxin is responsible for staphylococcal scalded syndrome (Al-Mebairik et al., 2016). In the current study, predominantly high frequencies of isolates were found to be positive for hla (97%) and hld (94%) genes. Consistently, studies from Egypt and China showed similar results (84–100%) in S. aureus food isolates (Mahros et al., 2021; Rong et al., 2017; Sallam et al., 2015), which indicates an overall high incidence of these genes among S. aureus isolates in food samples. Alpha-toxin encoded by hla and hld plays a significant role in tissue damage in invasive staphylococcal infections (Ahmad Mansour et al., 2021).
Although food poisoning caused by SEs is a common food-associated S. aureus disease, a few cases of deadly food-initiated invasive S. aureus diseases have also been reported (Kluytmans, 2010). In these cases, virulence factors other than SEs are responsible (Al-Mebairik et al., 2016). Therefore, investigating other virulence potentials of food-related S. aureus isolates is also essential.
Conclusions
The data suggest that a relatively high proportion of retail foods in Ardabil Province in Iran were contaminated with S. aureus (MSSA and MRSA). While antibiotic resistance was not common in MSSA isolates, MRSA isolates exhibited MDR characteristics. The study also found that S. aureus isolates carry a high rate of toxigenic genes, which could pose a significant threat to public health. In addition, the presence of MRSA lineages linked to humans suggests that retail foods are tainted with human origin. Therefore, to reduce the risk of MRSA transmission to consumers, it is necessary to implement proper hygiene measures throughout the food chain.
Footnotes
Acknowledgments
The authors wish to thank Dr. Firouz Amani (Ardabil University of Medical Sciences, Iran) for his help in statistical analysis.
Ethics Approval
Disclosure Statement
No competing financial interests exist.
Authors' Contributions
E.G.: Methodology, investigation, formal analysis, and original draft preparation. M.M.: Investigation and formal analysis. R.R.: Methodology, review, and editing. H.P.D.: Conceptualization, review, and editing. M.A.: Review and editing. M.A.: Supervision and project administration.
Funding Information
This study was financially supported by the vice-chancellor of research and technology of Ardabil University of Medical Sciences, Ardabil, Iran (Grant no. 400000676).
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.