
Abstract
Salmonella spp. is among the most central etiological agents in foodborne bacterial disorders. To identify Salmonella spp., numerous new molecular techniques have been developed conversely to the traditional culture-based methods. In this work, a new peptide nucleic acid fluorescence in situ hybridization (PNA-FISH) method was developed for the specific detection of Salmonella species, allowing a faster analysis compared with the traditional methods (ISO 6579-1: 2017). The method was optimized based on a novel PNA probe (SalPNA1692) combined with a blocker probe to detect Salmonella in food samples through an assessment of diverse-rich and selective enrichment broths. Our findings indicated that the best outcome was obtained using a 24-h pre-enrichment step in buffered peptone water, followed by RambaQuick broth selective enrichment for 16 h. For the enrichment step performance validation, fresh ground beef was artificially contaminated with two ranges of concentration of inoculum: a low level (0.2–2 colony-forming units [CFUs]/25 g) and a high level (2–10 CFUs/25 g). The new PNA-FISH method presented a specificity of 100% and a detection limit of 0.5 CFU/25 g of food sample, which confirms the great potential of applying PNA probes in food analysis.
Introduction
S
The FISH technology is a molecular technique for detecting and quantifying microorganisms (Emerson et al., 2017), and is widely applied for bacterial identification (Amann and Fuchs, 2008; Machado et al., 2013; Rocha et al., 2018). Through this technique, specific nucleic acid probes will bind to particular RNAs, due to their high number of copies (Amann and Fuchs, 2008; Rocha et al., 2018). Conventionally, the DNA or RNA probe is used in FISH procedures. However, its use may be unapproachable due to the lack of stability and specificity (Nácher-Vázquez et al., 2022). In food analysis, natural DNA/RNA probes may be hindered by the susceptibility to the food interferents (Rohde et al., 2015). The synthesis of nucleic acid mimics, as in the case of peptide nucleic acid (PNA) oligonucleotides, has been revealed to offer an improved hybridization performance compared with DNA probes (Cerqueira et al., 2008).
A few probes, including PNA probes, have already been developed for Salmonella detection and their theoretical specificity and sensitivity have been evaluated in a previous study (Almeida et al., 2010). Nonetheless, the rising of genomic data/databases now provides a more comprehensive evaluation of probes' sensitivity and specificity (Nácher-Vázquez et al., 2022), enabling the design of more accurate probes.
In this study, we have designed a new PNA probe for the specific detection of Salmonella spp., and its theoretical performance was compared with other probes previously described (Almeida et al., 2010). The inclusion of a blocker probe (BP) was evaluated to increase the specificity of the method.
Materials and Methods
Strains and culture maintenance
A total of 103 Salmonella spp. and 31 non-Salmonella spp. were included in this study (Supplementary Tables S1 and S2, respectively). The various bacterial strains were subcultured on tryptic soy agar (VWR, Carnaxide, Portugal) at 37°C for 24 h.
In silico specificity and sensitivity of the probes described in the literature
To evaluate the theoretical specificity and sensitivity of the probes described for Salmonella spp., we used the TestProbe analysis software at SILVA database (https://www.arb-silva.de/search) (Quast et al., 2013). First, the probes were aligned with sequences present in the large subunit (23S/28S) and small subunit (16S/18S) in the SILVA database. Specificity was calculated as nS/(TnS) × 100, where nS stands for the number of non-Salmonella strains that did not react with the probe and TnS for the total of non-Salmonella strains examined. Sensitivity was calculated as S/(TS) × 100, where S stands for the number of Salmonella strains detected by the probe and TS for the total number of Salmonella strains existent in the databases (Almeida et al., 2010).
In silico design of the PNA probe
To recognize potential oligonucleotides for the Salmonella spp. probe, it was selected a set of sequences from 16S (with lengths >1200 bp) and 23S (>1600 bp) collections available at the SILVA database. This selection comprises a wide number of Salmonella sequences, including typical strains of each subspecies, and other bacterial species from related species belonging to the Enterobacteriaceae family. The sequences were then aligned using the Clustal Omega tool (https://www.ebi.ac.uk/Tools/msa/clustalo). For all possible PNA probes, theoretical sensitivity and specificity were achieved and the best probe was selected taking into account specific features, as described in our previous studies (Almeida et al., 2011; Almeida et al., 2010). Subsequently, the selected sequence, designated here as SalPNA1692, was synthesized as previously reported (Almeida et al., 2010).
BP design
To block the nonspecific binding of SalPNA1692 probe with other Enterobacteriaceae sequences/strains, a BP was designed. The designed probe herein (SalPNA1692) was aligned with the 23S rRNA sequences from Salmonella spp. and other Enterobacteriaceae. For that, sequences from SILVA database were aligned using the Clustal Omega program accessible on the EBI website. The melting temperature and free energy were evaluated for both blocker and detection probes (Yilmaz and Noguera, 2004) to guarantee a comparable affinity to the corresponding target sequences. The BP was also synthesized as previously reported (Almeida et al., 2010), but without a fluorochrome.
Application of the PNA-FISH technique
The PNA-FISH method was carried out as previously described (Almeida et al., 2010) with some modifications. Smears of each strain were prepared by standard procedures, covered with 20 μL of hybridization solution containing 200 nM of PNA probe and 200 nM of BP. The smears were mounted with one drop of nonfluorescent immersion oil (Merck, Darmstadt, Germany) and covered with coverslips. The slides were stored in the dark for a maximum of 24 h before microscopy. During protocol optimization, the PNA-FISH was performed at different hybridization and washing temperatures (59–61°C) and formamide concentrations (5.5% and 30% [vol/vol]). For better discrimination between Salmonella spp. and non-Salmonella spp., it was selected the temperature of hybridization of 61°C and 30% (vol/vol) of formamide that was used subsequently throughout this study.
In vitro sensitivity and specificity of the PNA SalPNA1692+BP probe
The analytical sensitivity of Salmonella spp. probe was established using 103 different strains of Salmonella spp. Then, according to the obtained hybridization signal, the results were qualitatively classified into two categories: positive and negative. For the determination of analytical specificity, a set of 31 strains containing several genera from the Enterobacteriaceae family and closely related to the Salmonella species, including Citrobacter freundii, were included. These experiments were performed in triplicate.
Enrichment step optimization in food samples
To determine the effect of the enrichment step in the detection of Salmonella spp. by PNA-FISH, fresh sausage was used as a food matrix from a local retailer (Pingo Doce, Portugal). This matrix was chosen to assess the suitability of the method since it is frequently related to the prevalence of Salmonella, being also commonly associated with salmonellosis outbreaks (Mattick et al., 2002). As such, herein, to get a positive result for artificially contaminated samples with as low as 1 colony-forming unit (CFU)/25 g of food, we evaluated diverse enrichment broths. For this assay, we used the Salmonella Derby SGSC 2467. It is also important to highlight that both universal and selective enrichment broths were tested, namely, buffered peptone water (BPW; Liofilchem, Roseto degli Abruzz, Italy), tetrathionate broth (TT, Sigma-Aldrich), Muller Kauffmann broth (MK; Liofilchem), Muller-Kauffmann tetrathionate-novobiocin broth (MKTTn; Liofilchem), Rappaport Vassiliadis broth (RV; Liofilchem), and Ramba Quick broth (RQ; Chromagar, Paris, France).
In addition, two-step enrichment protocols were carried out using the following combinations: BPW-TT, BPW-MK, BPW-MKTTn, BPW-RV, and BPW-RQ. Of note that, in the first enrichment (BPW), the samples were incubated for 24 h, and in the second-step enrichment they were incubated for 16 h. The first medium (225 mL of BPW) was used to dilute the matrix composed of 25 g of food sample in a stomacher bag. Afterward, each tube containing 9 mL of each second-step enrichment medium was inoculated with 1 mL of the previous enrichment. At the end of the two-step enrichment, 20 μL of the sample was analyzed as reported before.
Salmonella detection in an artificially contaminated food matrix
To validate the effect of the enrichment step in the detection of Salmonella spp. by PNA-FISH, a different matrix was used, fresh raw ground beef from a local retailer (Pingo Doce). To achieve it, three bulk batches for the matrix were prepared according to what was described by Rocha et al. (2019). The detailed protocol for this section is described in Supplementary Data. Briefly, for the inoculation of fresh raw ground beef, the inoculum was prepared in brain–heart infusion broth by using fresh colonies of Salmonella Senftenberg SGSC 2516. Subsequently, the culture was incubated at 37°C for 18–24 h and then spread and mixed into the matrix bulk batches to obtain the two ranges of concentration of inoculum: a low level (LL) (0.2–2 CFU/25 g) and a high level (HL) (2–10 CFU/25 g) (Rocha et al., 2019).
A noninoculated batch is also kept and analyzed simultaneously with the LL and HL samples. From each batch, samples were taken (5 for the noninoculated; 20 for the LL- and 5 for the HL-batch) and pre-enrichment was carried out in the optimal conditions described in the previous section, as also described in Supplementary Table S3. Then PNA-FISH was performed as described above. Simultaneously, to confirm the presence of Salmonella, the ISO 6579-1: 2017 reference method (ISO 2017) was used, in this study, as a comparative method to PNA-FISH.
Afterward, for estimating/confirming the inoculation level in the food matrix sample, a most probable number (MPN) evaluation was carried out on the same meat batches (noninoculated, LL and HL) as described in Supplementary Data. As such, the number of positives from the different tested levels was used to calculate the MPN using the LCF MPN calculator (“Least Cost Formulations, Ltd., MPN Calculator Version 1.6,”).
Microscopy visualization
The Salmonella spp. population in each sample was discriminated by using the PNA-FISH method, as previously reported (Almeida et al., 2010). Briefly, this experiment was carried out using an OLYMPUS BX51 epifluorescence microscope following the protocol described by Rocha et al. (2019). It is also important to note that all the images were acquired with equal exposure time, using the Olympus Cell B software.
Results and Discussion
Salmonella detection is traditionally performed using culture methods, such as ISO6579-1:2017. However, it is a time-consuming and laborious process. Therefore, there is a need to develop new fast and reliable methods that can assist in the control of bacteria to reduce salmonellosis in humans and animals. To this end, an alternative approach based on PNA-FISH for the detection of Salmonella spp. from food samples was developed.
Salmonella probe design and evaluation
Before designing a new PNA probe, we carried out an in silico analysis to verify the specificity and sensitivity of the previously available probes in the literature for the detection of Salmonella. The determination of these parameters was carried out using equations previously reported (Almeida et al., 2010) and the calculations were performed in the same period of time, to assure the same number of sequences within the database. Although all the probes have high values of specificity, the sensitivity values presented a higher range of variation (Table 1). Both DNA probes' (Sal3 and Salm63) sensitivity values were less satisfactory, as a large number of Salmonella sequences on the database were not detected, 64 and 142, respectively.
Theoretical Specificities and Sensitivities of the DNA or PNA Probes for Detection of Salmonella spp. to Perform These Calculations, the SILVA Database Was Used in the Same Period
The number of Salmonella sequences on the database was 873.
PNA, peptide nucleic acid.
Regarding the two PNA probes, Sal23S10 and SalPNA1873, they already present higher sensitivity values. However, Sal23S10 has hybridized with a considerable number of non-Salmonella strains (621). The SalPNA1873, previously designed by Almeida and colleagues, had only 19 non-Salmonella strains hits, however, the sequences were mostly from Proteus mirabilis, a species that can be easily found in food matrices (Gong et al., 2019; Wang et al., 2010).
To overcome this potential cross-hybridization problem, a new PNA probe was designed by our group. As such, the identification of valuable oligonucleotides was achieved by aligning 16S and 23S RNA sequences of each subspecies of Salmonella and other bacterial species belonging to the Enterobacteriaceae family. Of the conceivable probe regions, one appeared to be the best option since it presents the highest specificity for Salmonella. According to this, the PNA oligomer sequence selected was as follows: 5′CACCAAGTACGGGA3′. The probe was named SalPNA1692, considering the starting position in 23S rRNA.
In silico evaluation of Salmonella spp. probe performance was performed as for the previous ones, using the TestProbe tool of SILVA database. The analysis of the SalPNA1692 probe resulted in a theoretical value of sensitivity of 96.449% and a specificity of 99.997%, as shown in Table 1. Although this probe presents a slightly lower sensitivity value (96.449%), it only matched one P. mirabilis (while SalPNA1873 matched six P. mirabilis sequences).
Further evaluation of the probes also showed a few one-mismatch differences to some closely related Enterobacteriaceae species (especially P. mirabilis and Escherichia coli sequence); for that reason and to avoid any possible cross-hybridization, the inclusion of a BP was considered. The value of melting temperature for SalPNA1692 probe was also calculated in silico, resulting in 58.3°C, while the Gibbs free energy resulted in −16.9.
Optimization of the FISH procedure
The hybridization protocol was based on a previous study (Almeida et al., 2010), however, hybridization conditions had to be optimized, namely the hybridization temperature. As such, we tested a range between 59°C and 61°C on some number of Salmonella and non-Salmonella strains, as shown in Supplementary Table S4. In addition, samples were visualized in the various available filters at the microscope, to make sure that no autofluorescence was observed. As it was possible to see in Supplementary Table S4, the probe presented higher specificity at the hybridization temperature of 61°C, and thus, this temperature was selected to be used in the next assays.
Improving the method specificity by including a BP
To evaluate the specificity of the SalPNA1692 probe together with the BP, a larger number of Salmonella and non-Salmonella spp. strains were tested, using 61°C as the optimal temperature (Supplementary Table S5). Given the fact that previous data support that, depending on the thickness of the peptidoglycan layer, formamide might damage bacteria cell envelope (Santos et al., 2014), here, we tested two different concentrations of the formamide in the hybridization solution (5.5% and 30% vol/vol). According to Supplementary Table S5, 30% (vol/vol) of formamide provides better results, showing no cross-hybridization with non-Salmonella spp. A previous study carried out by Almeida and coworkers, who developed another PNA probe (SalPNA1873) for the detection of Salmonella enterica serovar Enteritidis, also used a hybridization solution containing 30% (vol/vol) of formamide in their PNA-FISH protocol (Almeida et al., 2013).
The BP included to avoid possible cross-hybridization has the following sequence: 5′-ACACCAAGTACAGGA-3′. Similar to what was described for the PNA SalPNA1692 probe, herein, melting temperature (56.1°C) and Gibbs free energy (−16 ΔG) values were also calculated in silico.
To evaluate the effect of using a BP, a larger number of Proteus species were tested (Supplementary Table S6), considering the optimal hybridization conditions (30% vol/vol formamide and 61°C). Taking into account the results of Supplementary Table S6, no hybridization was observed for all strains of P. mirabilis tested, confirming the specificity of the SalPNA1692 + BP, as also verified in Figure 1.

Fluorescence microscopy images using the two probes (SalPNA1692 and BP) and DAPI staining.
SalPNA1692+BP sensitivity and specificity testing
The sensitivity and specificity of the SalPNA1692+BP were tested considering the hybridization conditions optimized above. As such, the protocol was applied to 103 representative Salmonella strains and 30 other non-Salmonella strains. The inclusivity study yielded 102 positive results in 103 strains tested, meaning a 99% sensitivity rate (Supplementary Table S1). The negative outcome observed for Salmonella Arizonae SGSC 2426 was already expected due to the sequence of the PNA probe used. The target region of ribosomal RNA of Salmonella spp. has slight differences in some strains of Salmonella Arizonae, and thus, not all strains of this particular serovar should be detected. The exclusivity study produced only negative outcomes (30 strains of different genera), which corresponds to 100% specificity (Supplementary Table S2).
Optimization of an enrichment step for Salmonella detection in artificially contaminated food matrices
After assessing the experimental sensibility and specificity, this technique was adapted to the detection of artificially contaminated food matrices. For that, a food matrix consisting of fresh sausage was selected and artificially contaminated with Salmonella Derby SGSC 2467 at an LL of concentration. After pre-enrichment in BPW for 24 h, followed by a selective enrichment with each of six different media for 16 h, pathogen detection was performed by PNA-FISH and confirmed by ISO 6579-1:2017, as shown in Table 2. According to the results, it was verified that in a second enrichment with RQ, 8 out of 10 confirmed positive samples (according to ISO) were also positive by using PNA-FISH. Using the most promising enrichment conditions, the next assays were carried out aiming for the method validation at different concentrations of artificial contamination.
Optimization of an Enrichment Step for Salmonella Detection Using Fresh Sausage as a Matrix (Low Level of Contamination)
According to ISO 6579-1: 2017.
BPW, buffered peptone water; FISH, fluorescence in situ hybridization; MKTTn, Muller-Kauffmann tetrathionate-novobiocin broth; PNA, peptide nucleic acid; RQ, Ramba Quick broth; RV, Rappaport Vassiliadis broth; TT, tetrathionate broth.
Salmonella detection in artificially contaminated food matrices
Based on the conditions optimized in the previous experiments, the validation was performed by using fresh raw ground beef (73% lean) as a matrix and inoculating Salmonella Senftenberg at LL and HL concentrations. Table 3 summarizes the results, by comparing the outcomes from the PNA-FISH assay, and the reference method ISO 6579-1:2017. Of note that the probability of detection (POD) was calculated as the number of positive outcomes divided by the total number of trials (Wehling et al., 2011). The POD was calculated for the candidate method results, PODC, the reference method results, PODR, and the difference in the candidate method results and reference method results, dPODC. The POD analysis between SalPNA1692+BP results and the reference method results showed that there was no statistically significant difference at the 5% level.
Performance of SalPNA1692+BP, Candidate vs. Reference Method ISO 6579-1:2017
MPN = most probable number is calculated using the LCF MPN calculator version 1.6 provided by AOAC RI, with a 95% confidence interval.
N = number of test portions.
x = number of positive test portions
PODC = candidate method confirmed positive outcomes divided by the total number of trials.
PODR = reference method confirmed positive outcomes divided by the total number of trials.
dPODC = difference between the confirmed candidate method result and reference method confirmed result POD values.
95% CI = if the confidence interval of a dPOD does not contain zero, then the difference is statistically significant at the 5% level.
BP, blocker probe; POD, probability of detection.
For the low inoculation level of Salmonella Senftenberg SGSC 2516, there were 11 positives for the candidate method and 11 confirmed positives for the reference method. For the high inoculation level, there were five positives for the candidate method and five confirmed positives for the reference method. According to these findings, for the low inoculation level, a dPODC value of 0.00 was obtained with a 95% confidence interval (−0.13 to 0.13), indicating no significant difference between the candidate and the reference methods. Similarly, for the high inoculation level, a dPODC value of 0.00 was obtained with a 95% confidence interval (−0.47 to 0.47), revealing, also, no significant difference between the candidate and the reference methods.
Figure 2 is an experimental example of the detection of Salmonella in fresh ground beef, which demonstrated the typical morphology of Salmonella spp. (Fig. 1A). Despite all the advantages of the PNA-FISH approach, it is also commonly found autofluorescence in tissues (Weaver et al., 2019) and food matrices (Rohde et al., 2015). However, it is also important to note that, in our study, it did not interfere or was otherwise minimalized through sample preparation and analysis.
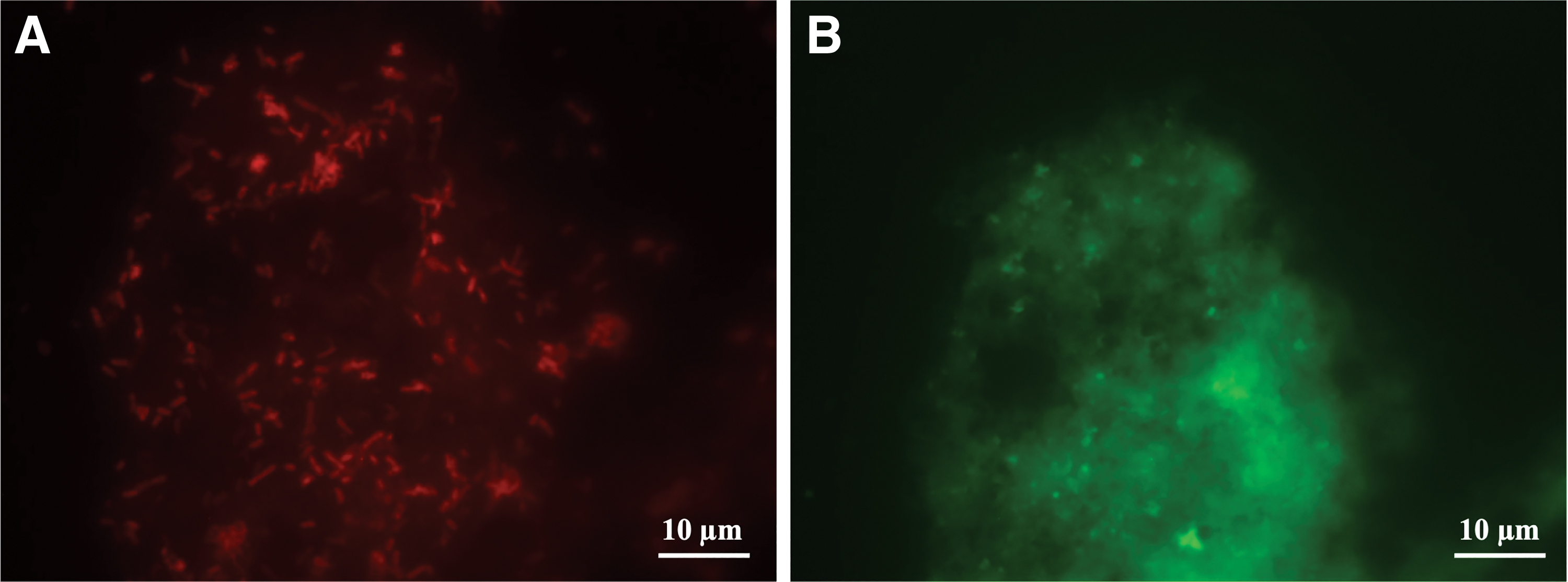
Detection of Salmonella spp. using the SalPNA1692+BP for fresh, raw ground beef.
Concluding Remarks
This study relates the development of a new detection method for Salmonella spp. in food matrices. The approach is based on a PNA-FISH technique that copulated the use of a new PNA probe for Salmonella, SalPNA1692, with a BP resulting in 96.449% of sensitivity and 99.997% of specificity. To be able to detect 1 CFU of Salmonella spp. in 25 g of sample, various selective and nonselective enrichment broths were assessed. Globally, the second step of the enrichment procedure in RQ medium provided the most reliable outcomes at the desired limit of detection. A validation assay in fresh ground beef (73% lean) demonstrated that the method presents a high accuracy performance.
The SalPNA1692 reveals a detection limit of 0.5 CFU/25 g of food sample, which is slightly better than those previously reported for the probes described in the literature, demonstrating, thus, the potential of the SalPNA1692 probe to be used in food analysis. While the obtained detection limit is very promising, it is also important to take into consideration that this matrix validation was only performed for meat, which means that different food matrices can present slightly different results. To conclude, the optimized Salmonella PNA-FISH method presents high specificity and sensitivity, and it is a faster time-to-result method compared with the conventional culture methods, making it a good candidate to be used for routine application in food safety laboratories.
Footnotes
Acknowledgments
We thank Prof. Javier Garaizar, Prof. Mónica Oleastro, Dr. Hugo Oliveira, Prof. Séamus Fanning, Dr. Nuno Cerca, and Dr. Sara Cleto for providing some of the strains used in this study.
Authors' Contributions
N.F.A., L.C., and C.A. conceived and designed the experiments; M.S. and R.R. performed most of the experiments. A.B. helped to perform experiments. M.S., R.R., N.F.A, L.C., and C.A. analyzed the data; D.A. and J.C. wrote the article. The overall editing was performed by N.F.A, L.C., and C.A. All the authors have read and agreed to the published version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financially supported by: FCT/MCTES (PIDDAC): LEPABE, UIDB/00511/2020 (DOI: 10.54499/UIDB/00511/2020) and UIDP/00511/2020 (DOI: 10.54499/UIDP/00511/2020) and ALiCE, LA/P/0045/2020 (DOI: 10.54499/LA/P/0045/2020). This article is a result of project FLUDS—Desenvolvimento de sistemas baseados em espectroscopia de fluorescência para detecção microbiana, with reference NORTE-01-0247-FEDER-046970, cofunded by the European Regional Development Fund (ERDF), through the North Portugal Regional Operational Programme (NORTE2020), under the PORTUGAL 2020 Partnership Agreement. PhD fellowship ![]() and CEEC Individual (2022.06886.CEECIND) were supported by national funds through FCT—Fundação para a Ciência e a Tecnologia.
and CEEC Individual (2022.06886.CEECIND) were supported by national funds through FCT—Fundação para a Ciência e a Tecnologia.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.