
Abstract
Salmonella enterica is a common foodborne pathogen that poses significant safety risks across the world. And benzalkonium bromide (BK) is widely used as a disinfectant to sterilize the food processing equipment. It has been reported that sub-lethal concentration of disinfectants induced not only the homologous resistance but also cross-resistances. This work analyzed the induced resistances of Salmonella Enteritidis by short-term adaptation (STA) and long-term adaptation (LTA) to BK. We have demonstrated that inefficient sterilization exposes Salmonella Enteritidis to sub-lethal concentrations of BK, and adapts bacteria to a higher minimum inhibitory concentration and minimum bactericidal concentration. In addition, STA, but not LTA, to BK induced heterogeneous resistance to sodium hypochlorite, and cross-resistance to freezing, desiccation, and heating, which may be caused by the membrane composition change of Salmonella Enteritidis. This work could be useful to the optimization of cleaning protocol.
Introduction
S
S. enterica is mostly transmitted via the fecal-oral route, and undercooked or contaminated foods are major sources of infection (Mkangara, 2023; Teklemariam et al., 2023). Among the 2500 identified serovars, Salmonella enterica serovar Enteritidis is the major cause of foodborne bacterial salmonellosis in the United States and Europe (Tarabees et al., 2017).
To control the contamination of foodborne microorganisms, quaternary ammonium compounds (QACs) are widely used as disinfectants of the hard surfaces of food processing equipment (Zhou et al., 2023). In a typical sanitation cycle, a cleaning reagent is applied to the food processing equipment and rinsed off, followed by the spraying and rinsing off of a disinfectant (Moretro et al., 2017). QACs contain a positively charged nitrogen atom (N+), covalently bound to four carbon atoms, of which at least one carbon is substituted with a long alkyl chain (Zhou et al., 2023).
Additionally, the anion ionically bound to the N+ is usually Bromide (Br−) or Chloride (Cl−) (Zhou et al., 2023). Because of the unique structure, QACs are endowed with physical and chemical features, such as emulsification, dispersion, solubilization, and sterilization (Zhou et al., 2023). Due to the high efficiency and low toxicity, benzalkonium bromide (BK) is a commonly used QAC in medical and industrial areas (Huang et al., 2019).
A few studies reported that a sub-lethal environmental stressor adapted foodborne pathogens, such as S. enterica, Listeria monocytogenes, and Bacillus cereus, to multiple stress factors (He et al., 2016; Huang et al., 2019; Kim et al., 2021; Meena et al., 2016). Although cross-resistance stimulated by various sub-lethal stressors has been reported, fewer reports discussed the impacts caused by the duration of the same disinfectant.
Residue BK may remain on the surface of the food processing equipment for different time duration due to inefficient rinsing or irregular inner surfaces (Moretro et al., 2017). In addition, most studies focused on cross-resistances induced by heat shock, osmosis, or drought, whereas actual disinfectants applied in the food processing industry, BK for example, have been rarely explored. This work mainly analyzes the homologous, heterogeneous, and cross-resistance of Salmonella Enteritidis induced by short-term and long-term treatment of BK. In addition, the biological characteristic changes and potential molecular mechanisms of resistances caused by BK adaption have been studied.
Materials and Methods
Strain and culture preparation
Salmonella Enteritidis CICC21482 (China Center of Industrial Culture Collection) was stored in 25% glycerol at −80°C. Before the experiment, this bacterium was streaked onto tryptic soy agar (TSA; Hope Bio-Technology Co., Qingdao, China) and incubated overnight at 37°C. A single colony was then inoculated into 3 mL tryptic soy broth (TSB; Hope Bio-Technology Co.) at 37°C for 24 h. The stationary phase culture was used for further studies.
Determination of minimum inhibitory concentration and minimum bactericidal concentration
Assays were described previously with slight modifications (Jordán et al., 2013). Sanitizers were prepared in TSB medium. BK (Shanghai Macklin Biochemical Co.) was prepared to 12.5, 25, 50, 100, 200, and 400 (ppm); ethanol (Tianjin Zhiyuan Chemical Reagent Co., Tianjin) was 0%, 5%, 10%, 20%, 40%, and 80% (v/v); and sodium hypochlorite (Aladdin Reagent Co., Shanghai) was 6%, 3%, 1.5%, 0.75%, 0.375%, and 0.1875% (v/v, available chlorine).
Two hundred microliters of each concentration was pipetted into a 96-well plate, followed by addition of 20 μL of Salmonella Enteritidis cells at 7 log10 colony-forming unit (CFU)/mL. Samples were incubated at 37°C for 24 h. Minimum inhibitory concentration (MIC) was recorded as the lowest concentration that completely inhibited the visible growth of bacteria.
In addition, 100 mL of the liquid culture with no visible growth was transferred to a TSA plate with the same sanitizer concentration. Minimum bactericidal concentration (MBC) was recorded as the lowest concentration at which bacteria failed to grow on the plate.
Short-term adaptation
Three hundred microliters of bacterial suspension was added to 3 mL TSB medium with 25 ppm BK (1/2 MIC). Cells were incubated at 37°C, 180 rpm for 24 h, followed by being pelleted at 25°C, 106 g for 2 min. Samples were washed twice by Buffered Peptone Water (BPW; HuanKai Microbial, China), and they were re-suspended in 3 mL BPW.
Long-term adaptation
This method was described previously with slight modifications (Ribič et al., 2020). Three hundred microliters of bacterial suspension was added to 3 mL TSB medium, and an appropriate amount of BK was added to reach the final concentration of 10 ppm. Cells were incubated at 37°C, 180 rpm for 24 h. If visible growth was observed, an aliquot of 300 μL was inoculated to 3 mL TSB medium, and BK was added to reach 20 ppm.
The cell density was counted by serial dilution and plate spreading. This process was repeated with an increment of 10 ppm for every passage until no visible growth was observed. The highest concentration Salmonella could tolerant without affecting its growth was used for long-term adaptation (LTA). The LTA cells were pelleted at 25°C, 106 g for 2 min. Samples were washed twice by BPW, and they were re-suspended in 3 mL BPW.
Heat resistance measurement
Non-adapted (NA), STA, and LTA Salmonella Enteritidis cells were suspended in TSB, pasteurized milk (Inner Mongolia Yili Industrial Group Co., Ltd.), carrot juice (Zhejiang Wei Quan Co.), spaghetti sauce (Chef Boyardee), and Nutella (Ferrero) to reach 7 log10 CFU/mL. One milliliter of suspension was added to a 1.5 mL sterile centrifuge tube, and it was heated at 55°C (HB-100, Bioer, China) for 0, 1, 2, 4, and 8 min, followed by immediate cooling on ice and serial dilution in BPW. Cells were spread-plated on TSA plates, incubated at 37°C for 24 h, and enumerated. The cross-resistance to heat was determined by the D-values (minutes), defined as the time required to kill 90% of the Salmonella population.
Freezing resistance measurement
NA, STA, and LTA cells were suspended in TSB to reach 7 log10 CFU/mL. One milliliter of suspension was added to a 1.5 mL sterile centrifuge tube, and it was incubated at −20°C for 0, 6, and 12 h. Samples were then thawed at 37°C in a water bath, and they were 10-fold serially diluted for enumeration. The reduction of strain populations was used to characterize the induced resistance.
Desiccation resistance measurement
NA, STA, and LTA cells were suspended in TSB to reach 6 log10 CFU/mL. One hundred milliliters of bacterial suspension was transferred to the center of a filter with a pore size of 0.22 μm. The filter was laid flat on a disposable sterile petri dish, which was incubated for 24 h at 25°C in a desiccator with the relative water activity of 0.30 ± 0.05. Samples were vortexed in 10 mL of BPW for 3–5 min. The rehydrated bacterial suspension was serial diluted with BPW, and it was spread-plated on TSA plates. The reduction of strain populations was recorded.
Determination of cell membrane integrity with selective medium
This method was described previously with slight modifications (Lim et al., 2013). Briefly, NA, STA, and LTA cells were serial diluted in BPW. One hundred μL of each dilution was spread-plated on a TSA plates and xylose lysine deoxycholate (XLD) plates (HuanKai Microbial, China), respectively. Samples were incubated at 37°C for 24 h, and the CFU/mL was counted. The CFU/mL difference between the TSA and the XLD plates was an indicator of cell membrane damage.
Determination of cell membrane integrity with propidium iodide staining
This method was described previously with slight modifications (Kim et al., 2021). Briefly, cells were centrifuged at 10614 g for 10 min. The pellet was washed three times with 0.85% sterile saline. The suspended cells were mixed with the fluorescent dye propidium iodide (PI; Bestbio, Shanghai, China), followed by incubation in the dark for 15–20 min at 25°C.
Samples were then washed with sterile saline to remove excess dye. After washing, cells were resuspended in sterile saline. Fluorescence was measured in a microplate reader (DLJ-100D, DLJ biotechnology Co., Nanjing, China) at an excitation length of 490 nm and an emission length of 630 nm.
Determination of cell membrane fatty acid composition
Extraction of fatty acid methyl esters was described previously with a slight modification (Sasser, 2001). Briefly, 0.5 g of pelleted cells collected by centrifugation (10614 g, 5 min) was used for the analysis of fatty acid methyl esters. Subsequently, two-thirds of the organic phase was pipetted into a Gas Chromatography vial and capped.
The fatty acid methyl esters were analyzed using GCMS-QP2010 PLUS Gas Chromatography (SHIMADZU Inc., Tokyo, Japan) with a polar PEG capillary column (RTx-Wax, 0.25 μm, 30 m × 0.25 mm; Restek, Bellefonte, PA, USA). One microliter of the sample was injected with the injector temperature at 250°C and gas flow at a constant pressure of 1 mL/min. The oven temperature was held at 100°C for 1 min and then increased to 250°C at a rate of 4°C/min; detector temperature was held at 250°C for 5 min. Helium was used as the carrier gas.
Expression of fatty acid synthesis-related genes
All experiments were performed according to the reagent manufacturers' protocols. Total RNA was extracted using RNAiso Plus (Code No.: 9108; TaKaRa). Complementary DNA was synthesized with the PrimeScript™ RT reagent kit (Code No.: RR037A; TaKaRa). The real-time PCR was performed by TB Green® Premix Ex Taq™ II FAST qPCR (Code No.: CN830S; TaKaRa) with primers: cfa (forward [F], TCGTTGGGGTTAGGTGAGAG, reverse [R], ATTTCAGCTTGGCTTGCTGT) and fadA (F, ATCTCTCCGCCCACTTAATGCGTA, R, AGCCTTGCTCCAGCGTT-TGTTGTA). The relative expressions of the target genes were calculated using the 2−ΔΔCt method (Livak and Schmittgen et al., 2001).
Statistical analysis
All assays were carried out in triplicate in at least two separate experiments. The resulting data were evaluated via one-way analysis of variance in IBM SPSS Statics 21. Duncan's method was utilized to assess the difference between two different groups (p < 0.05).
Results
BK induced various cross-resistances in short-term adapted Salmonella Enteritidis
Salmonella Enteritidis treated with a sub-lethal concentration (1/2 MIC; Table 1) for 24 h was chosen as the STA group. As residual BK would be accumulated at the corner, dent, or irregular part of the interior surface of food processing machine, a higher sub-lethal concentration for the LTA group was chosen. A continuous adaptation test with increased BK concentration was performed, and 130 ppm BK was selected as the sub-lethal concentration, which was the highest concentration tolerated by Salmonella Enteritidis without affecting its growth (Supplementary Fig. S1).
Minimum Inhibitory Concentrations and Minimum Bactericidal Concentrations of Benzalkonium Bromide Adapted Salmonella Enteritidis to Different Sanitizers
BK, benzalkonium bromide; LTA, long-term adapted; MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration; NA, non-adapted; STA, short-term adapted.
For homologous BK resistance, both the MIC and MBC of the STA group, and of the LTA group, showed a two-fold increase compared with the NA group, respectively (Table 1). For sodium hydrochloride resistance, short-term BK adaptation increased the MIC and MBC to 0.75 and 1.5 (%, v/v), whereas LTA decreased these values to 0.1875 and 0.375 (%, v/v) (Table 1). Moreover, the MIC and MBC of ethanol were almost unchanged for both the LTA and short-term adaptation (STA), except for the MIC of LTA.
We then tested the resistance of cells to freezing stress by incubating the NA, STA, and LTA cells at −20°C for 6 and 12 h (Fig. 1A). The STA cells exhibited increased freezing resistance, such that the population reduction was ∼1.06 log10 CFU/mL after 12 h comparing with ∼1.34 log10 CFU/mL of the NA cells (p ˂ 0.05; Fig. 1A). On the contrary, LTA cells showed ∼3.90 log10 CFU/mL population reduction, nearly three-fold more cell loss compared with the NA cells (p ˂ 0.05; Fig. 1A).
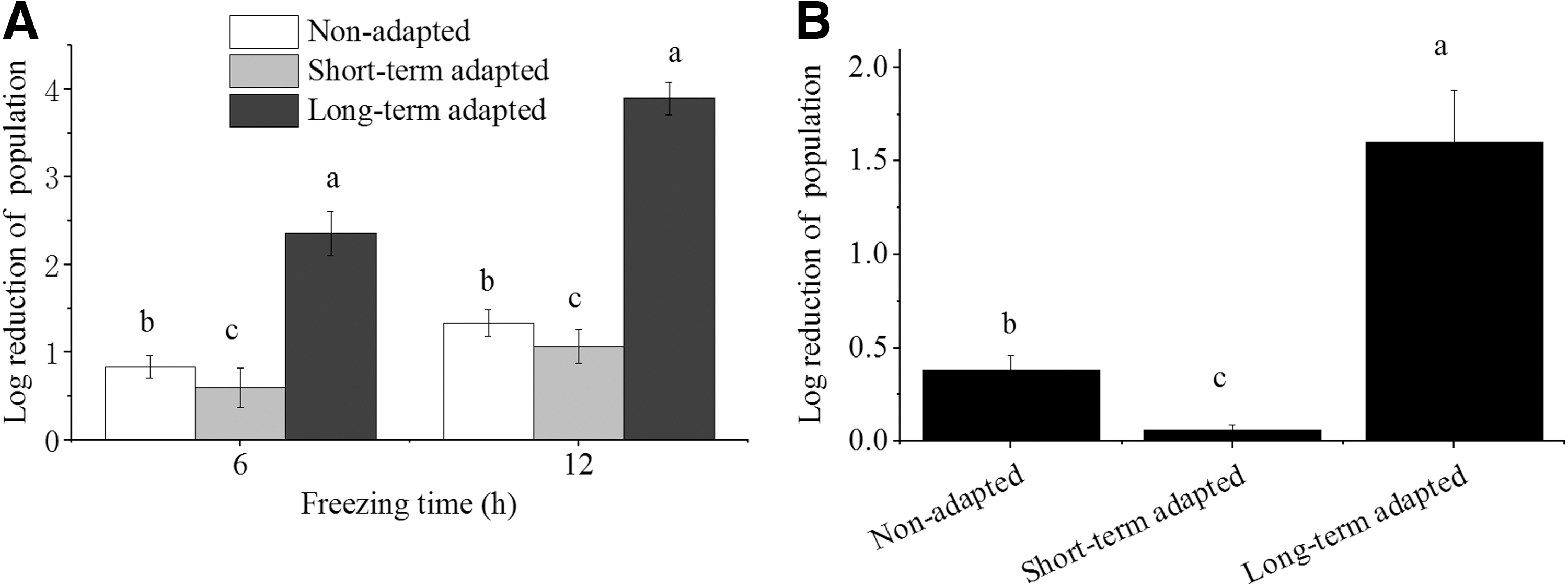
Log reduction of population for benzalkonium bromide adapted Salmonella Enteritidis in freezing stress
Similar to freezing, the Salmonella Enteritidis cells showed the same resistance pattern to drying stress, such that the STA group exhibited a higher resistance than the NA group, whereas the LTA group showed a lower resistance than the NA group (p ˂ 0.05; Fig. 1B).
To mimic the real food processing conditions, the thermal resistance of BK adapted Salmonella Enteritidis was tested in TSB medium, and in a few real-world foods, which represented a different food matrix. The D-value of the STA group in TSB medium was increased, whereas the LTA group was decreased compared with the NA group (p ˂ 0.05; Table 2), consistent with the resistance pattern of sodium hypochlorite, freezing, and desiccation resistances.
D-Value of Benzalkonium Bromide Adapted Salmonella Enteritidis in Different Media
Means with the same letter are not significantly different (p > 0.05).
LTA, long-term adapted; NA, non-adapted; STA, short-term adapted; TSB, tryptic soy broth.
However, this pattern was broken in real-world foods, such that the D-value of the STA group in carrot juice, spaghetti sauce, and Nutella, and of the LTA group in pasteurized milk, were not significantly changed (p ˃ 0.05; Table 2), probably due to the very different solute composition of these media.
Cell membrane integrity was compromised in LTA Salmonella Enteritidis cells
The cell membrane integrity was determined by comparing the CFU on TSA plates (the non-selective agar) and XLD agar plates (the selective agar). When the membrane, as a permeability barrier, was damaged, cells would be more sensitive to the bile acid in the XLD media, and, therefore, cell density would be reduced. The CFU of the NA and STA group in XLD medium, and in TSA medium, showed no significant difference, respectively (p ˃ 0.05; Fig. 2A), whereas the CFU of the LTA group was significantly dropped in XLD medium, compared with the TSA medium (p ˂ 0.05; Fig. 2A).
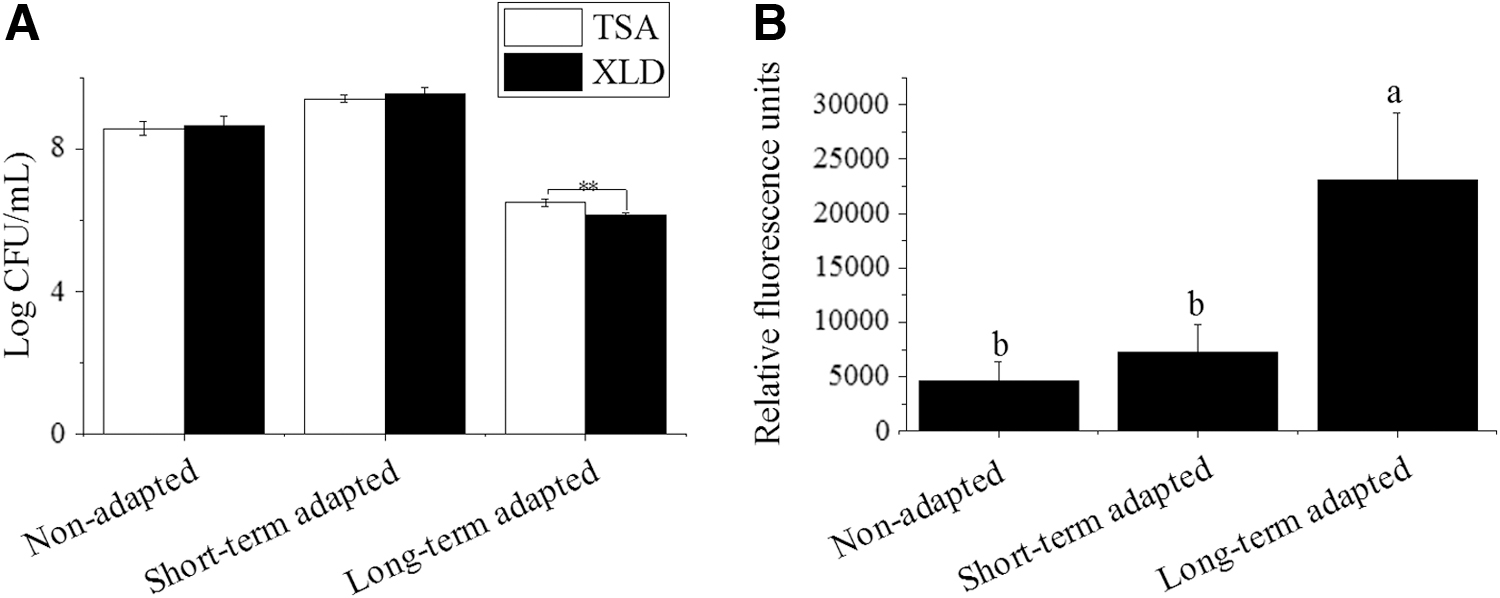
We then compared the fluorescent dye PI uptake of the STA and LTA cells with the NA cells to measure the membrane damage level. In accordance with the cell viability test, there was no significant PI uptake difference between the NA and STA cells (p ˃ 0.05; Fig. 2B). However, the PI uptake in LTA cells was significantly increased, suggesting increased cell membrane damage (p ˂ 0.05; Fig. 2B).
The membrane fatty acid composition of NA and adapted Salmonella is shown in Figure 3 and Supplementary Table S1.
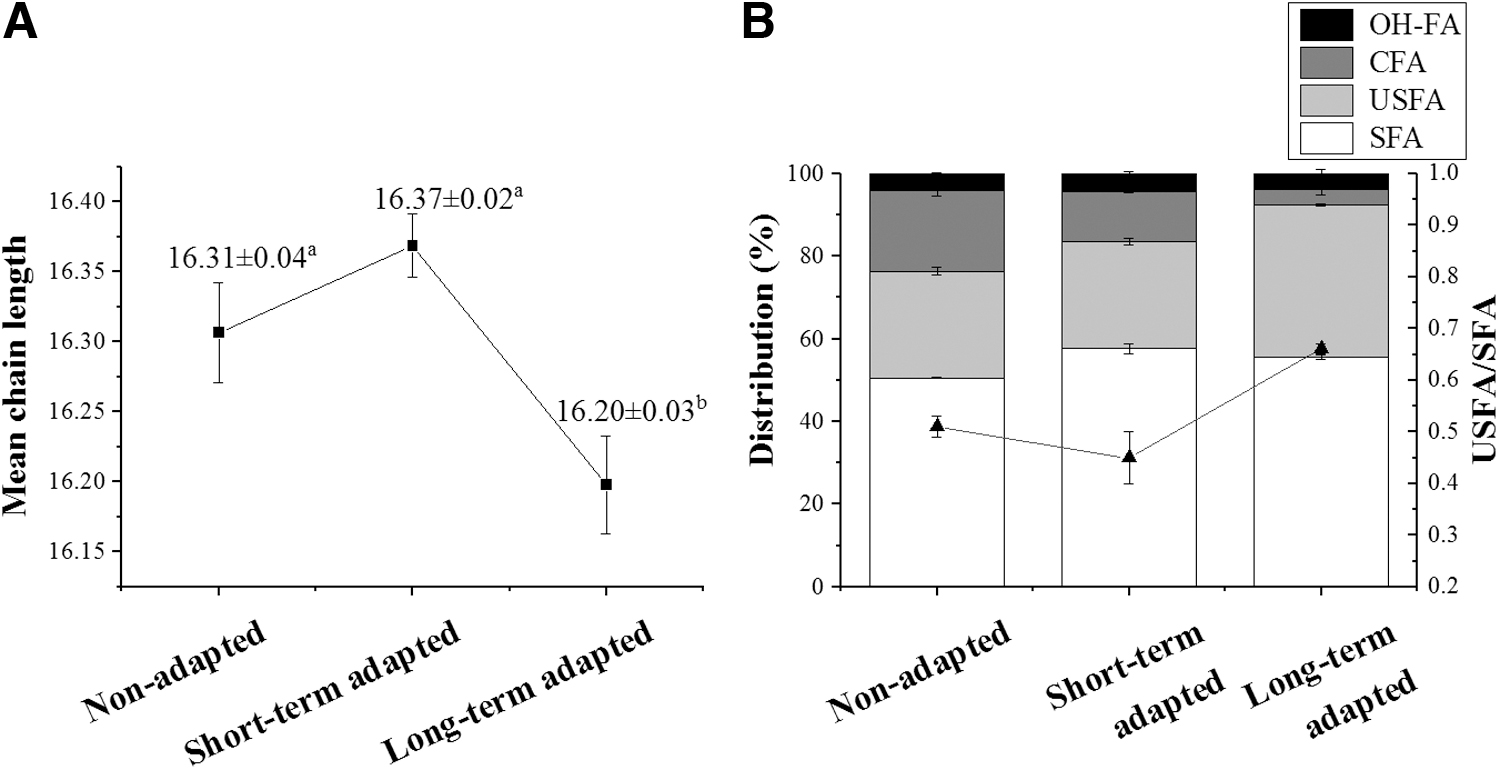
LTA decreased the mean chain length (p ˂ 0.05; Fig. 3A). And the percentage of total cyclic fatty acid (CFA) decreased ∼40% in STA cells, and ∼80% in LTA cells, compared with NA cells (p ˂ 0.05; Supplementary Table S1). The de-saturation degree (unsaturated fatty acid [USFA]/saturated fatty acid [SFA]) was calculated as an indirect indicator of the membrane fluidity. This ratio was decreased to 0.45 ± 0.05 for the STA group, but increased to 0.66 ± 0.01 for the LTA group, compared with the NA group (p ˂ 0.05; Fig. 3B).
Discussion
BK is a widely applied disinfectant in the food industry. However, sub-lethal concentrations of disinfectants are known to induce homologous and cross-resistances (Huang et al., 2019; Kim et al., 2021; Meena et al., 2016). Our data revealed that STA, but not LTA, to BK induced heterogeneous resistance to sodium hydrochloride, but not to ethanol. The anti-microbial activity of BK is through altering the membrane permeability (Huang et al., 2019). The hypochlorite ion is a strong oxidizing agent, reacting with a wide variety of biological molecules, including membrane lipids (Fukuzaki, 2006).
On the contrary, the anti-microbial activity of ethanol is mostly through denaturing the bacterial proteins (Hasan et al., 2021). It is possible that the heterogeneous resistance to sodium hypochlorite is induced because both sodium hypochlorite and BK attack the membrane lipids.
USFA is more difficult to pack densely than SFA, and, therefore, increases membrane fluidity, a key property for maintaining cell functionality (Denich et al., 2003; Fonseca et al., 2019; Loffhagen et al., 2001). As a consequence, bacteria lower the membrane fluidity by increasing the composition of SFAs to cope with various environmental stresses (Broadbent et al., 2014; Chen and Gänzle, 2016; Denich et al., 2003; Fonseca et al., 2019; Fritz et al., 2007; Li et al., 2011; Niu et al., 2022).
Consistently, our data demonstrated the de-saturation degree (USFA/SFA) of the STA group was lower than the NA group, whereas this value in the LTA group was significantly higher than the NA group. This was possibly the reason we observed cross-resistances in the STA group only.
To characterize the molecular mechanism of membrane composition change, we tested the expression of two mostly relevant genes, cfa, encoding an enzyme synthesizing cyclopropane fatty acids from monounsaturated fatty acids, and fadA, introducing a trans double bond to synthesize USFAs (Broadbent et al., 2014; Chen and Gänzle, 2016; Fritz et al., 2007; Li et al., 2011).
However, these two genes seem not to be the cause of membrane composition changes (Supplementary Fig. S2). Interestingly, both the LTA and STA induced the homologous resistance, suggesting more than one metabolic pathway was involved in BK adaptation. Besides membrane fluidity changes, reports have demonstrated that cross-resistances and antimicrobial resistances may be caused by mutations in bacterial genome, the genes coding for AcrAB-TolC efflux pump for example (Pye et al., 2023; Wang et al., 2023; Wu-Chen et al., 2023a; Wu-Chen et al., 2023b). Therefore, further study of whole-genome sequencing and RNA-seq would help to understand the genetic determinants of these differences.
Cleaning-in-Place (CIP) is a significant operation in the modern food industry, which is involved in automated cleaning of the interior surfaces of food processing equipment and pipes by jetting or spraying disinfectants, without disassembly of the equipment and with minimum or no manual operation (Tamine, 2008). Our data suggested the CIP protocol needs to be optimized according to the resistance type.
The cross-resistances induced by residual BK may be overcome by simply increasing the duration of sanitization time. However, extension of the sanitation time cannot overcome resistances induced by heterogenous stressors not functioning on membrane fluidity. A combination of different sanitizers, whose heterogeneous resistances will not be induced by each other, may be required.
Conclusion
We have revealed the STA, but not the LTA, of sub-lethal concentration of BK induced the heterogenous sodium hypochlorite resistance, and cross-resistances of Salmonella Enteritidis. Our data further suggested that the integrity of cell membrane and the change of membrane composition might be the reasons for the differences. Interestingly, both the LTA and STA induced homologous resistances, suggesting the BK induced resistance involved more than one metabolic pathway.
Footnotes
Authors' Contributions
Z.H. designed the project. F.R. carried out the majority of experiments. J.J., X.T., H.D., L.Z., and Z.Z. helped with the experiments. J.Z. and Z.H. wrote the manuscript. J.Z., L.S., and Z.H. reviewed and edited the manuscript.
Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
This study was supported by the National Natural Science Foundation of the people's Republic of China and the Science (Grant Nos. 31972044 and 32370029).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.