
Abstract
Members of the Bacillus cereus group are well-known opportunistic foodborne pathogens. In this study, the prevalence, hemolytic activity, antimicrobial resistance profile, virulence factor genes, genetic diversity by enterobacterial repetitive intergenic consensus (ERIC)-polymerase chain reaction (PCR) genotyping, and adhesion potential were investigated in isolates from a Tunisian dairy farm environment and raw milk. A total of 200 samples, including bedding, feces, feed, liquid manure, and raw bovine milk, were examined. Based on PCR test targeting sspE gene, 59 isolates were detected. The prevalence of B. cereus group isolates in bedding, feces, liquid manure, feed, and raw milk was 48%, 37.8%, 20%, 17.1%, and 12.5%, respectively. Out of the tested strains, 81.4% showed β-hemolytic on blood agar plates. An antimicrobial resistance test against 11 antibiotics showed that more than 50% of the isolates were resistant to ampicillin and novobiocin, while a high sensitivity to other antibiotics tested was observed in most isolates. The distribution of enterotoxigenic genes showed that 8.5% and 67.8% of isolates carried hblABCD and nheABC, respectively. In addition, the detection rate of cytotoxin K (cytk), enterotoxin T (bceT), and ces genes was 72.9%, 64.4%, and 5.1%, respectively. ERIC-PCR fingerprinting genotype analysis allowed discriminating 40 different profiles. The adhesion potential of B. cereus group on stainless steel showed that all isolates were able to adhere at various levels, from 1.5 ± 0.3 to 5.1 ± 0.1 log colony-forming unit (CFU)/cm2 for vegetative cells and from 2.6 ± 0.4 to 5.7 ± 0.3 log CFU/cm2 for spores. An important finding of the study is useful for updating the knowledge of the contamination status of B. cereus group in Tunisia, at the dairy farm level.
Introduction
The Bacillus cereus group, also known as Bacillus cereus sensu lato, comprises a growing list of genetically closely related Gram-positive, spore-forming bacterial species (Carroll et al., 2022). The most prominent members are as follows: B. anthracis, B. cereus sensu stricto, B. thuringiensis, B. weihenstephanensis, B. mycoides, B. pseudomycoides, B. cytotoxicus, and B. toyonensis.
Members of the B. cereus group are widely distributed in the environment and commonly isolated from soil, water, air, and plants. Due to the formation of endospores highly resistant to heat, dehydration, and physical stresses, the B. cereus group has a strong possibility to contaminate various raw, heat-treated, and processed food products such as cereal grains, vegetables, meat, seafood, milk, and dairy products (Berthold-Pluta et al., 2019; Biesta-Peters et al., 2016; Gdoura-Ben Amor et al., 2018; Samapundo et al., 2011).
Moreover, the B. cereus group can cause two different types of food poisoning in humans, namely emetic and diarrheal (Granum, 1994). The emetic syndrome is caused by ingesting of a heat-stable cereulide, a nonribosomal peptide synthetase encoded by the ces gene cluster (Ehling-Schulz et al., 2005; Rajkovic et al., 2008).
The diarrheal syndrome is associated with the production of heat-labile enterotoxins (EntS), including hemolysin BL (Hbl), nonhemolytic enterotoxin (Nhe), and cytotoxin K (CytK) (Beecher and Wong, 2000; Fagerlund et al., 2004; Granum et al., 1999; Lund et al., 2000). In addition, enterotoxin FM (EntFM), enterotoxin T (bceT), and EntS have been described as potential diarrheal toxins; however, their role in the development of the disease is not clear (Agata et al., 1995; Choma and Granum, 2002; Hansen et al., 2003).
The B. cereus group has been highlighted as concerns for the dairy environment (Cruz-Facundo et al., 2023; Fusieger et al., 2023; Neokleous et al., 2023). The monitoring of B. cereus group in milk and dairy products is important since it presence can lead to both spoilage and safety problems.
Spores of B. cereus in milk are a source of contamination for milk-derived products, as the spores are resistant to heat treatment. In fact, milk pasteurization processes are not effective in reducing contamination and can instead act as an activator of spore germination. In this regard, it is essential to consider the isolates in raw milk, as many of the dairy products are still made with this milk (Adame-Gómez et al., 2020; Fei et al., 2019; Owusu-Kwarteng et al., 2017). In addition to spore production that the B. cereus group can produce, this process has been considered a possible way that products are systematically contaminated (Christison et al., 2007; Shemesh and Ostrov, 2020).
B. cereus group has been isolated not only from dairy products but also from dairy farms, mainly from feed, cattle feces, bedding, and milking equipment that come into touch with the cows' udders, which can explain its presence in the final product (Tirloni et al., 2022). Therefore, it is crucial to evaluate B. cereus group from the dairy farm, during production and until the product's sale.
To date, there is currently no reported study on the prevalence of B. cereus group in a dairy farm environment in Tunisia. Therefore, the specific objectives of this study were to assess the prevalence, hemolytic activity, antimicrobial resistance profile, virulence factor genes, genetic diversity by enterobacterial repetitive intergenic consensus (ERIC)-polymerase chain reaction (PCR) genotyping and adhesion potential of B. cereus group isolated from a dairy farm environment in Sfax, Tunisia.
Materials and Methods
Sampling
A total of 200 samples, including bedding (n = 50), feces (n = 45), liquid manure (n = 30), feed (n = 35), and raw milk (n = 40), were collected from a local dairy farm in the governorate of Sfax located in the south of Tunisia, from September 2020 to December 2021.
Isolation of presumptive B. cereus group strains
B. cereus group isolation was performed according to Owusu-Kwarteng et al.'s (2017) method.
In brief, 10 g of each sample was homogenized with 90 mL of buffered peptone water containing 5 g/L of lithium chloride. Serial dilutions were prepared, and 0.1 mL of each diluted sample was streaked in MYP agar medium (Oxoid, Basingstoke, UK) and incubated for 24 h at 30°C.
Suspected B. cereus group colonies with pink-orange color surrounded by a precipitation zone were enumerated and a typical colony from each plate was subcultured on BHI-YE agar (Fisher Bioblock, Illkirch, France) and incubated for 24 h at 30°C. These strains were stored at −80°C in glycerol 25% for further identification.
Detection of hemolytic activity
According to Da et al. (2014), a 24-h culture of each isolate was streaked on blood agar plates (Oxoid) and incubated at 30°C for 24–48 h.
Identification of B. cereus group by PCR
Genomic DNA was extracted using the Chelex extraction method as described by Techer et al. (2014). As reported by Kim et al. (2005), B. cereus group was identified by amplification of 71 bp sequence of sspE gene, using the following primers:
sspE1-F (5′-GAAAAAGATGAGTAAAAAACAACAA-3′) and
sspE1-R (5′-CATTTGTGCTTTGAATGCTAG-3′).
The amplification conditions and the composition of the PCR used were described in Gdoura-Ben Amor et al.'s (2018) study.
ERIC-PCR genotyping
For ERIC-PCR, the primer 5′ -ATGTAAGCTCCTGGGGAT TCAC-3′ (Versalovic et al., 1991) was used. The amplification conditions and the composition of the PCR used were described in Gdoura-Ben Amor et al.'s (2018) study.
The comparison between gel patterns was made using the unweighted-pair group method with arithmetic mean/Dice, optimization 1%, and similarity 80%. The data were analyzed with the BioNumerics 6.5 software (BioMerieux, Belgium).
Detection of virulence genes
Confirmed B. cereus group strains were screened by PCR for the presence/absence of EntS and emetic toxin encoding genes. The primers used, their annealing temperatures, and the size of the amplified fragment are shown in Table 1.
Primers Used in the Simplex Polymerase Chain Reaction for the Detection of Virulence Genes in Bacillus cereus Group
bceT, enterotoxin T; cytK, cytotoxin K; hbl, hemolysin BL; nhe, nonhemolytic enterotoxin.
The amplification conditions, the composition of the PCR, and positive controls used were based on our previous study (Gdoura-Ben Amor et al., 2019).
Antibiotic susceptibility testing
The Kirby–Bauer disk diffusion method (Bauer et al., 1966) was used. A total of 11 antibiotics (Oxoid) were tested: ampicillin (10 μg), vancomycin (30 μg), gentamicin (10 μg), erythromycin (15 μg), tetracycline (30 μg), ciprofloxacin (5 μg), chloramphenicol (30 μg), novobiocin (30 μg), streptomycin (10 μg), kanamycin (30 μg), and rifampicin (5 μg). The diameter of the inhibition zone was determined according to the CLSI disk diffusion breakpoints of Staphylococcus aureus (CLSI, 2010).
Adhesion on stainless steel
Treatment of stainless-steel coupons
The adhesion test was studied on 2-mm-thick and 15-mm-diameter 304 L stainless-steel coupons (Bretagne Laser, Guer, France). The coupons were treated according to the protocol of Jan et al. (2011), which consisted of immersing the coupons in 2% (v/v) RBS 35 solution (Sigma) followed by a series of rinses with distilled water and autoclaving.
Preparation of spore suspensions
As recommended by Leguerinel et al. (2000), sporulation was induced by adding magnesium sulfate (40 mg/L) and calcium chloride (100 mg/L) in nutrient agar at 30°C. After 5 d of incubation, the culture was harvested, heated for 10 min at 80°, and then centrifuged 3 times at 4000 g for 10 min. The sporal suspensions are stored at 4°C in sterile water until use.
Adhesion test
The adhesion test is performed by adding 300 μL of bacterial (vegetative cells or spores) suspension (106 colony-forming unit [CFU]/mL) in each well. After a 2-h incubation at 30°C, suspensions were eliminated and the coupons maintained in the wells were rinsed with 9 g/L NaCl to remove nonadhering cells.
To detach adherent cell, the coupons were scraped for 1 min after addition of 300 μL of saline at 9 g/L of NaCl and 0.02% of Tween 80 for the vegetative cell and spore adhesion assays, respectively.
The number of adhering cells was then measured after growth on BHI-YE agar at 30°C for 24 h, using a plate counting micromethod (Baron et al., 2006). Tests were done in triplicate and results were expressed as average log CFU per cm2.
Statistical analysis
All assays were performed in triplicate, and the results were calculated as the mean ± standard deviation. The statistical significance was carried out using the Excel program. The differences between measurements were considered significant at p < 0.05.
Results
Isolation and identification of bacterial strains
Among the 200 examined samples, 65 were positive for B. cereus group-like strains with an overall prevalence of 32.5%% (Table 1).
PCR test targeting the sspE gene showed that 59 isolates are confirmed to belong to B. cereus group.
As shown in Table 2, the highest number of isolates was found in bedding samples (24/50, 48%), followed by feces (17/45, 37.8%), liquid manure (7/30, 20%), feed (6/35, 17.1%), and raw milk (5/40, 12.5%) samples.
Prevalence of Bacillus cereus Group Bacteria in Local Dairy Farm Samples Collected in Tunisia During the Period from September 2020 to December 2022
PCR, polymerase chain reaction.
The incidence of B. cereus group strains in environmental samples (33.75%) was much higher than for raw milk samples (12.5%).
Hemolytic activity
Out of the 59 confirmed B. cereus group isolates, 48 isolates (81.4%) were found to be β-hemolytic, 7 isolates (11.9%) showed indeterminate hemolytic activity, while the remaining 4 isolates (6.7%) were γ-hemolytic.
ERIC-PCR typing analysis
Considering 80% similarity in band pattern as cutoff criteria, the analysis of the ERIC-PCR profiles allowed to discriminate 40 profiles on a total of 59 tested strains.
As shown in Figure 1, 24 isolates were unclustered, while the remaining 35 isolates were grouped into 16 clusters.
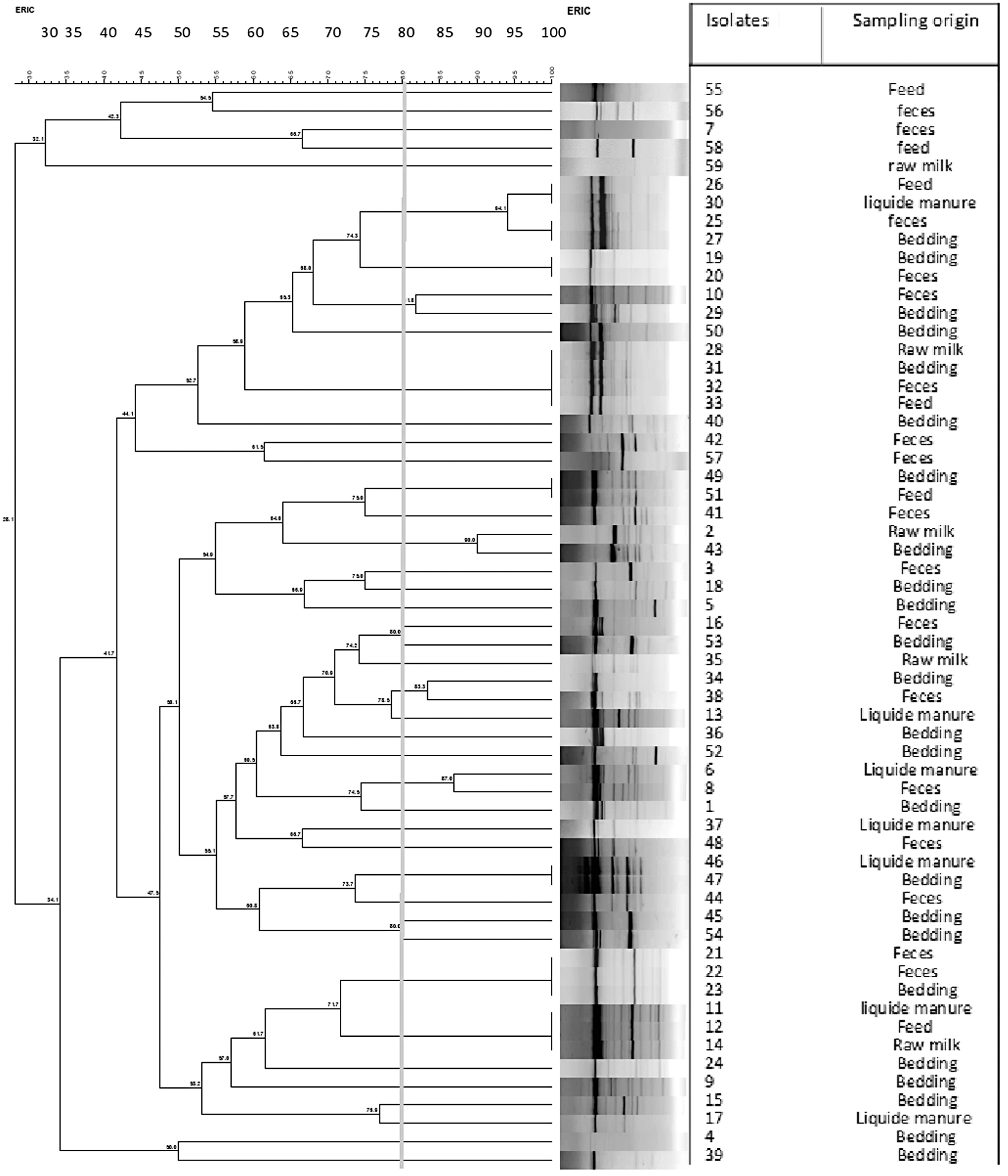
ERIC-PCR dendrogram showing the relationship between Bacillus cereus group isolates. The similarities between strains were evaluated using the Dice coefficient and the UPGMA clustering method. Genetic similarity between samples in duplicate is 80%. ERIC, enterobacterial repetitive intergenic consensus; PCR, polymerase chain reaction; UPGMA, unweighted-pair group method with arithmetic mean.
Distribution of virulence factor genes
The distribution of virulence genes among the 59 isolates is shown in Table 3.
Distribution of Enterotoxin and Emetic Toxin Genes in Bacillus cereus Group Strains Isolated from Dairy Farm Environment and Raw Milk in Tunisia
bceT, enterotoxin T; cytK, cytotoxin K; HBL, hemolysin BL; NHE, nonhemolytic enterotoxin.
At least one gene of each of the NHE and HBL complexes was detected in 100% and 52.5% of strains, respectively.
For NHE encoding genes, 67.8% (40/59) of isolates were found to harbor simultaneously the nheABC genes, 27.1% (16/59) harbored simultaneously two genes, and 5.1% (3/59) harbored only one gene.
For HBL encoding genes, 18.7% (11/59) possessed only one gene, 13.5% (8/59) possessed simultaneously two genes, 11.8% (7/59) possessed simultaneously three genes, 8.5% (5/59) possessed simultaneously all four hblABCD genes, and 47.5% (28/59) possessed no HBL encoding gene at all.
The cytK gene was present in 72.9% of the strains, but further testing revealed that strains harboring the cytK gene belonged to the cytK-2 variant. The cytK-1 gene was not found in any isolate.
The prevalence of bceT among isolates was 64.4%.
The prevalence of the emetic gene ces among isolates was 5.1%. The emetic gene was only detected in strains isolated from raw milk samples but not from environmental samples.
Antibiotic susceptibility
The results of the antimicrobial tests are presented in Table 4.
Antibiotic Susceptibility of 59 Bacillus cereus Group Strains Isolated from Dairy Farm Environment and Raw Milk in Tunisia
More than 50% of the isolates were resistant to ampicillin (54.2%) and novobiocin (50.8%). They were, however, susceptible to other antimicrobials such as rifampicin (100%), chloramphenicol (100%), ciprofloxacin (100%), gentamicin (100%), streptomycin (88.1%), kanamycin (96.6%), erythromycin (86.4%), vancomycin (83%), and tetracycline (72.9%).
Adhesion ability of B. cereus group on stainless steel
All the isolates were able to adhere on stainless steel at various levels. The values recorded are between 1.5 ± 0.3 and 5.1 ± 0.1 log CFU/cm2 for vegetative cells and between 2.6 ± 0.4 and 5.7 ± 0.3 log CFU/cm2 for spores (Fig. 2).

Study of the adhesion capacity of spores and vegetative cells of 59 isolates of the Bacillus cereus group on stainless-steel coupons.
In our study, it appears that 72.9% of the collection had greater attachment to stainless-steel surfaces of spores than vegetative cells of the same strain. For 8.5% of the strains, spores and vegetative cells showed the same adhesion capacity. While for 18.6% of the strains, the vegetative cells showed a higher adhesion than the spores (Fig. 3).

Comparison of the adhesion capacity of spores and vegetative cells of 59 Bacillus cereus group isolates.
Discussion
The B. cereus group is considered an important contaminant in the dairy environment and is related to milk quality and safety (Tirloni et al., 2022).
Hence, minimizing B. cereus group transmission at the farm level is crucial, as spores in used bedding (Magnusson et al., 2007) and soil (Christiansson et al., 1999) are major sources of contamination of raw milk via contaminated teats and udder surfaces.
The prevalence of B. cereus group in dairy farm environment varies across studies.
Compared with the prevalence of B. cereus group in our study (29.5%), a higher prevalence was reported by McAuley et al. (2014) and Meng et al. (2022) with 41% and 43%, respectively. Conversely, Fei et al. (2019) reported a lower prevalence (18%), whereas Cui et al. (2016) reported a similar prevalence (28.7%).
The high prevalence founded in bedding samples (48%) is comparable with that recorded by Fei et al. (2019) (40%), whereas Cui et al. (2016) reported a higher prevalence (93.3%).
The low prevalence of B. cereus group was recorded in raw milk samples (12.5%). Compared with our data, a higher prevalence was reported in raw milk samples analyzed by Hammad et al. (2021), Meng et al. (2022), Owusu-Kwarteng et al. (2017), and McAuley et al. (2014) with 85%, 61.11%, 46.6%, and 33%, respectively. Conversely, a lower B. cereus group prevalence in raw milk was reported by Cui et al. (2016) (9.8%) and Fei et al. (2019) (4%).
The low prevalence in raw milk (12.5%) compared with environmental samples (33.75%) is due to the inhibitory action of the natural microflora (lactic acid bacteria) in raw milk (Tirloni et al., 2017) and/or the effective hygienic practices during the milking process.
Hemolytic activity is crucial for assessing bacteria safety (Nwagu et al., 2020), as β-hemolysis indicates the presence of cytotoxic phospholipases (Sorokulova et al., 2008), a hemolytic factor reducing iron availability for the host (Seker et al., 2010).
Similar to our finding, high frequencies (>80%) of hemolytic activity in B. cereus group strains have been reported (Arslan et al., 2014; Chaves et al., 2011; Chica et al., 2020).
In our study, ERIC-PCR genotyping allowed a better assessment of the biodiversity of strains. Our finding is in agreement with Chen et al.'s (2022) study demonstrating the high discriminatory capability of ERIC-PCR.
ERIC-PCR was easier and faster than other molecular typing methods such as pulsed-field gel electrophoresis and ribotyping (Dorneles et al., 2014; Magyar et al., 2019). However, due to its limited repeat capabilities, more accurate methods such as multilocus sequence typing and genomic sequencing may be more informative (Nguyen and Tallent, 2019; Shen et al., 2021; Zhang et al., 2020).
In this study, ERIC-PCR genotyping highlighted that some genetic relatedness is not correlated with sample origins. This diversity could suggest various origins of contamination (Tirloni et al., 2022).
The screening for the presence of virulence factors showed that HBL complex genes were less common than that of the NHE complex with a detection rate of 67.8% and 8.5%, respectively. Similar results of high prevalence of nheABC genes have previously been reported in strains from raw milk, dairy plants, and environmental samples (Lin et al., 2017; Zhao et al., 2020).
Conversely, various studies have reported higher prevalence rates, usually between 40% and 70.6% of the HBL complex in strains from milk and dairy products (Gao et al., 2018; Owusu-Kwarteng et al., 2017; Porcellato et al., 2021).
The cytK gene was present in 72.9% of the strains. A similar occurrence was observed in strains from raw milk, farms, and dairy products (Gao et al., 2018; Owusu-Kwarteng et al., 2017; Zhao et al., 2020). However, our detection rate of cytK is higher than previously reported by Meng et al. (2022).
The prevalence of bceT gene among strains was 64.4%. Although bceT encoded by bceT gene is categorized as a diarrheal EntS, its role in the development of the disease is not clear (Agata et al., 1995; Choma and Granum, 2002; Hansen et al., 2003).
The emetic gene ces was detected in 5.1% of B. cereus group strains isolated from raw milk samples. Previous studies have found a low incidence of ces gene in B. cereus group strains from dairy products (Andersen Borge et al., 2001; Cui et al., 2016; Owusu-Kwarteng et al., 2017; Svensson et al., 2006). However, several studies showed a high prevalence of ces gene among strains isolated from milk, dairy farm environments, and ultra-high-temperature milk processing lines (Cui et al., 2016; Lin et al., 2017; Meng et al., 2022; Owusu-Kwarteng et al., 2017).
Briefly, virulence factors vary across studies due to geographical locations, strain sources, and PCR assay primers, potentially affecting the results.
Antibiotic therapy remains the predominant treatment for B. cereus group infections. However, the emergence of antibiotic-resistant strains due to misuse of antibiotics or the acquisition of resistance genes leads to the failure of antibiotic treatment (Banerjee and Sarkar, 2004). Therefore, in this study, the resistance to antibiotics was determined, and we found that most of the isolates were sensitive to rifampicin, chloramphenicol, ciprofloxacin, gentamicin, streptomycin, kanamycin, erythromycin, vancomycin, and tetracycline, while more than 50% of the isolates were resistant to ampicillin and novobiocin.
Resistance to beta-lactams as well as sensitivity to different groups of antibiotics, including glycopeptides, aminoglycosides, tetracyclines, quinolones, and phenicols, has been reported previously in strains from farm environment, milk, and dairy products (Chang et al., 2021; Gao et al., 2018; Owusu-Kwarteng et al., 2017; Radmehr et al., 2020; Zhao et al., 2020).
The adhesion test showed that 72.9% of the collection had greater attachment to stainless-steel surfaces of spores than vegetative cells of the same strain. This finding is in agreement with the study of Andersson et al. (1998) showing that B. cereus group spores are 33–48% more adherent than vegetative cells.
The high hydrophobicity of spores enables them to adhere to the surfaces of food processing equipment and this can raise hygiene issues in the food industry due to their resistance to cleaning procedures (Andersson et al., 1998; Faille et al., 2010; Peng et al., 2001; Shaheen et al., 2010).
Our study shows that in 18.6% of strains, vegetative cells are more adherent than spores.
The high adhesion of vegetative cells could be attributed to their increased hydrophobicity during the stationary phase, the stage when they are placed in contact with coupons (Peng et al., 2001), as well as to their mobility, as flagella plays a crucial role in adhesion stages (Houry et al., 2010).
Conclusions
To conclude, this study forms the starting point for the control of B. cereus group in dairy products and production plants. The results of our work were useful for updating the knowledge of the contamination status of B. cereus group in Tunisia, especially at the dairy farm level. There is therefore the need to observe good hygienic and manufacturing practice by milk producers and traditional dairy processors to prevent contamination and the subsequent potential disease outbreak by B. cereus group.
Footnotes
Acknowledgments
This work is part of a doctoral thesis by R.B.A. The authors are grateful to all the collaborators in this study. They would like to thank the Ministry of Higher Education and Scientific Research of Tunisia and the European Commission of Erasmus+ program for their financial support.
Authors' Contributions
R.B.A.: conceptualization, methodology, and original draft preparation. M.G.-B.A.: writing—original draft preparation. H.S.: writing—reviewing and editing. N.G.: conceptualization and methodology. S.J.: visualization and investigation. M.G.: supervision and validation. R.G.: writing—reviewing and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
The 1st author's mobilities in France to carry out this work have been financed by the Ministry of Higher Education and Scientific Research of Tunisia and the European Commission of Erasmus+program.