
Abstract
Salmonella Typhimurium is a foodborne pathogen threatening livestock and human health. It is highly resistant to commonly used clinical antibiotics, and it is urgently needed to explore new anti-Salmonella treatment schemes. In this study, first, our in vivo mouse experiments showed that Baitouweng decoction (BTW), a classical Traditional Chinese Medicine (TCM) prescription, had good efficacy against Salmonella Typhimurium infection: mitigating weight loss of mice; lowering the bacterial load of liver, spleen, and colon; reducing the production of serum inflammatory factors (interleukin-1β and tumor necrosis factor-α); and decreasing histological index scores than that in the Salmonella Typhimurium infection group. Furthermore, we explored the potential active components and molecular mechanism of BTW in the treatment of Salmonella Typhimurium infection. A total of 465 compounds of BTW were retrieved from herb website and 227 bioactive compounds were identified, 911 potential BTW-related targets and 1,602 disease targets of Salmonella Typhimurium infection were acquired by ten public analytical databases, among them, 188 genes were overlay targets of BTW-Salmonella Typhimurium; String, Metascape, and Cytoscape plug-in Molecular Complex Detection and ClueGo analysis pointed that BTW exerted an anti-Salmonella effect through a multicomponent, multitarget, and multipathway manner, including 10 hub targets (TNF, AKT CASP3, ALB, EGFR, JUN, MAPK, STAT3, VEGFA, and TP53) and 94 pathways such as cell apoptosis, inflammation, and metabolism. Finally, AutoDock Vina showed that the hub target AKT1 with menispermine and quercetin had good binding energy, which was confirmed by the in vitro cellular thermal shift assay and drug affinity responsive target stability assay. This study laid the foundation for further study of BTW mechanism and for further development of BTW anti-Salmonella.

Introduction
Salmonella Typhimurium, a foodborne pathogen that threatens livestock and human health, is one of the most widespread bacteria in the world and has received widespread attention (Hardy, 2004). Salmonella is not a ubiquitous microbe (Ferrari et al., 2019). Salmonella is a pathogen that, either in animals or in humans, normally colonizes the intestinal microbial flora (Linam and Gerber, 2007). More than 2,600 serotypes of Salmonella Typhimurium are known, which is a great public health concern (Cooper and Cheryl, 2020). According to the famous Koch’s postulates on the determination of bacterial pathogenicity, Salmonella Typhimurium is a non-Bacillus species that infects the intestines of humans and animals, causing acute gastroenteritis (Kidgell et al., 2002). Salmonella presence indicates water or food contamination through inadequate hygiene management in some regions or farms, and is secondary to infections in animals and humans (Newell DG et al., 2010). After the host ingests substances contaminated with Salmonella Typhimurium through feces or contaminated food and water sources, the bacteria inhabit the neutral, micro-oxygenated, and relatively sterile small intestine and eventually enter the large intestine, which is rich in bacteria, and flourish in the anaerobic environment (Leclerc et al., 1998). According to World Organization of Animal Health and the European Union, the control of salmonella in animal production should and needs to be based on prevention and not on the use of antibiotics (Anand Kumar P et al., 2023). In modern animal husbandry, it is a common practice for decades to use antibiotics to prevent and treat animals from diseases and to improve production performance. For example, the case fatality risk for calves first treated during the second week was lower after treatment with trimethoprim–sulfadoxine and oxytetracycline with polyvinylpyrrolidone (Harland RJ et al., 1991). However, similar to humans, the use of certain antibiotics can result in life-threatening intestinal overgrowth of opportunistic bacteria (Dethlefsen L et al., 2008). Also, those who survive are at greater risk of becoming chronic carriers of Salmonella. It has been reported that of the 1,387 patients surviving the infections, 2.3% were chronic Salmonella Typhi and Salmonella Paratyphi B carriers (Vogelsang and Bøe et al., 1948). The emergence of drug-resistant strains has also contributed to the great difficulty in controlling Salmonella Typhimurium infection (Mølbak, 2005). However, using new and effective prevention or treatment options for antibiotic resistance in humans and animals judiciously would be an important step (Stanton, 2013). Thus, it is important to identify and develop safe, efficient, and nontoxic drugs.
Baitouweng decoction (BTW) had suitable therapeutic efficacy against Salmonella Typhimurium infection.
• Ten hub targets and 94 pathways were involved in the BTW treatment of Salmonella Typhimurium.
• The hub targets interacted with their corresponding active compounds both in silico and in vitro.
Radix pulsatilla decoction (BTW) is a classical Traditional Chinese Medicine (TCM) formula. The great efficacy of TCM without obvious adverse reactions has been valued (Zhao et al., 2021). TCM works through a “multicomponent target network” means, compared with the “one target, one drug” methods of Western medicine (Lam, 2001). BTW was first reported in “Treatise and Febrile Diseases,” which was written by Zhang Zhongjing, a famous Chinese physician. It has been used for many years to treat intestinal inflammation and fight bacterial infections. BTW includes four components, namely, Pulsatilla chinensis (Bunge) Regel (Baitouweng), Coptis chinensis Franch (Huang Lian, HL), Phellodendron chinense C. K. Schneid (Huang Bo, HB), and Fraxinus chinensis Roxb (Qin Pi, QP) (Linet al., 2019). Modern pharmacodynamic research has shown that BTW has antibacterial and anti-inflammatory properties, promotes immune regulation, and treats diarrhea and endotoxin damage to the body (Wu et al., 2023). For example, BTW induced AMPK phosphorylation and inhibited mTOR expression to repair DSS-induced barrier damage in mouse colitis intestinal epithelium (Pan et al., 2022). It also has a suitable therapeutic efficacy in the treatment of bacterial dysentery (Wang et al., 2019) and ulcerative colitis (Yu et al., 2021). Modern pharmacological studies have also shown that BTW has a multiangle treatment of inflammation. Clinically, a BTW has shown positive effects on inflammation and diarrhea (Muluye et al., 2014). BTW has been used to treat Salmonella-infected rats by regulating the metabolism of glycerol and phospholipids (Hua et al., 2020). In addition, the active constituents of the four main herbal plants have anti-inflammatory, antibacterial, and antitumor effects (Wang et al., 2016). However, the specific efficacy of BTW against Salmonella remains unknown and needs to be further explored.
Network pharmacology is a technique that provides insights for the design of effective clinical drugs with minimal side effects and toxicity (Wang et al., 2019). Network pharmacology has been widely used in many fields by establishing a network “drug-gene-target-disease” system. It reveals the complex relationship between many components of TCM and target proteins and the prevention mechanism of toxic and side effects (Pan et al., 2022). The complex interactions between TCM ingredients and disease-related targets can be investigated with integrated network analysis (Zhang et al., 2013). Network pharmacology also has been used to screen the active ingredients of TCM and further explore the relationships between drugs and organisms as well as the pathogenesis and diseases (Huo et al., 2020; Lu et al., 2021). This study revealed that the regulation of complicated signaling pathways by BTW was related to its therapeutic effect, providing novel insights for drug research and development.
Materials and Methods
BTW preparation
Pulsatilla chinensis (Bunge) Regel (Lot: L23S21C124236, HPLC ≥98%) (15 g), Phellodendron chinense C.K. Schneid. (Lot: L22S41C125306, HPLC ≥97%) (12 g), Coptis chinensis Franch. (Lot: L23S31C125436, HPLC ≥96%) (6 g), and Cortex fraxini (Lot: L23S11C125305, HPLC ≥98%) (12 g) were all purchased from the Affiliated Hospital of Changchun University of Chinese Medicine (Changchun, China) and identified by Professor Jinwei Zhao. The above components were put into 1,000 mL of distilled water for 30 min and heated twice after adding more water. The filtrates were combined, and the solution was concentrated to 100% (1 g crude drug/mL) and stored at 4°C for use in subsequent experiments.
Animal grouping
Fifty healthy 8-week-old SPF C57BL/6 mice were purchased from Changchun Yisi Experimental Animal Technology Co., Ltd, half male and half female. Experimental animals (license number 01021684806951458), weighing 18–22 g, were raised in an environment with a controlled temperature (22 ± 2°C), relative humidity (50 ± 5%), and a 12-h/12-h light/dark cycle. The mice were allowed to move freely and had access to distilled drinking water ad libitum. The experiment followed the relevant regulations of animal welfare and was approved by the Institutional Animal Care and Use Committee of Jilin University (SY202311024). After one week of acclimatization to the laboratory environment, mice were randomly divided into five groups as follows: Control (C) group, Salmonella Typhimurium infection model (M) group, BTW low-dose treatment (BTWL) group, BTW medium-dose treatment (BTWM) group, and BTW high-dose treatment (BTWH) group. After fasting for 4 h, mice were given 20 mg/kg streptomycin (MedChemExpress, New Jersey, United States) orally. The mice were then given water immediately and food. After 20 h, water abstinence and fasting for 4 h were used for follow-up experiments. There were 10 mice in each group. The experiment lasted ten days.
Bacterial culture
Salmonella Typhimurium (ATCC14028) was preserved in our laboratory and cultured in broth medium (LB) to a logarithmic growth period at 37°C and 180 rpm. HeLa cells were purchased from ATCC (Maryland, USA) and maintained in RPMI 1640 medium (GIBCO, NY, USA) supplemented with 10% fetal bovine serum (FBS, GIBCO, NY, USA). The cells were incubated at 37°C with 5% CO2.
Protection test
The groups were treated as follows: the C group was treated with equal amounts of normal saline; the M group was treated with 105 CFU/mL of bacteria by gavage as previously reported (Zhao et al., 2018); the BTWL group was administered 7.5 g/kg BTW; the BTWM group was administered 22.5 g/kg BTW; and the BTWH group was administered 45 g/kg BTW. Cell counting plates directly calculated the number of Salmonella Typhimurium. The M, BTWL, BTWM, and BTWH groups were administered a uniform dose of bacteria (105 CFU/mL in 1 mL PBS), and the BTW treatment was administered 3 times by gavage per day. The number of mice that died was recorded daily.
Weight index monitoring
Each mouse was weighed every 24 h after dosing. The weight recovery of mice after administration of different doses of BTW was analyzed.
Measurement of inflammatory factor levels
Blood from each group of mice was collected from the head after sacrifice, left at room temperature for 30 min, placed in a 4°C refrigerator overnight, centrifuged at 3,000 rpm for 10 min, and stored at 20°C. Interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α) levels in serum were measured by indirect enzyme-linked immunosorbent assay (Wu et al., 2012).
Colonization number detection of different organs
The colonization number of different organs was measured by reference to previous experiments (Myhal et al., 1982). Briefly, different groups of mice were sacrificed daily. Mouse liver, spleen, and colon tissues were aseptically harvested and weighed, and 0.2% Triton X-100 was used to grind tissues and organs so that the final concentration is 1.0 g/mL and a 10% tissue homogenate was prepared. The tissue homogenate was diluted again. Fifty microliters of tissue homogenate was applied to LB plates and the number of bacteria in the organ tissues of different treatment groups was counted after overnight incubation in a biochemical incubator at a 37°C anaerobic environment.
Pathological sectioning and hematoxylin and eosin staining
The mice were sacrificed, and the liver, spleen, and colon were harvested and photographed. Parts of the liver, spleen, and colon were removed, the tissue was fixed with 4% formaldehyde after rinsing with PBS, dehydrated, permeabilized, embedded, sectioned, and stained with hematoxylin and eosin (H&E) (Luo et al., 2017). The sections were put onto slides, dried, and cover slipped to observe the morphological changes of the liver, spleen and colon tissues in different treatment groups under a microscope.
Herb compounds, herb compound target, and disease prediction target preparation
Herb compound preparation
We logged into the HERB database (http://herb.ac.cn/) to find the main therapeutic ingredients in BTW, HL, HB, and QP. The given information only referred to ingredient ID, ingredient name, and ingredient alias. Therefore, we identified other indicators one by one, such as the SMILES format (PubChem website, https://pubchem.ncbi.nlm.nih.gov/), molecular weight (MW), and other indicators of the compounds.
Screening each active herb compound of BTW and their targets
In our research, we screened the active compounds of BTW. Screening each compound of TCM can help elucidate their pharmacokinetics (Li et al., 2022). An ideal drug would comply with the physicochemical property guidelines of Lipinski’s rule of five (RO5) (Lipinski et al., 2001). According to RO5, a drug compound should have an MW of <500 g/mol, a log p value of ≤5 representing its hydrophobicity, rotatable bonds ≤10, hydrogen bond donors ≤5, and hydrogen bond acceptor sites ≤10 (Doak et al., 2014; Veber et al., 2002). We used the Swiss ADME web tool (http://www.swissadme.ch/) to evaluate the compounds according to RO5 (Daina et al., 2017; Zeng et al., 2021). Furthermore, the compound targets were predicted through the following two databases: PharmMapper (http://lilab-ecust.cn/pharmmapper/, PM) and Swiss Target Prediction (http://www.swisstargetprediction.ch/, STP) (Daina et al., 2019; Wang et al., 2017). We compared the active ingredients identified by the two databases according to normal fit scores and finally selected ingredients that met the normal fit scores ≥0.8 for the PM database and the normal fit scores ≥0.1 for the STP database for subsequent research. These active ingredients corresponded to the four main ingredients of BTW.
Salmonella Typhimurium infection target prediction
Salmonella Typhimurium infection targets were comprehensively retrieved from the following 6 online disease databases with “Salmonella Typhimurium infection” as the search term: Comparative Toxicogenomics Database (https://ctdbase.org/, CTD), DisGeNET (https://www.disgenet.org/home/), GeneCards (https://www.genecards. org/), DrugBank (https://go.drugbank.com), OMIM (https://omim.org/), and GEO (https://www.ncbi.nlm.nih.gov/gds/). The official names of the target genes were obtained from the UniProt website and uniformly converted into UniProt IDs (Wang et al., 2019), and the species was selected as Homo sapiens. The target genes with the highest scores were chosen for subsequent research.
Network construction method
Comparison of the potential BTW compound targets and Salmonella Typhimurium infection target genes
Network construction using Cytoscape software (version 3.9.3) systematically explored the relationships between the effective ingredients and the corresponding targets. We established a list of key ingredients contained in BTW and the target genes of Salmonella Typhimurium infection and then combined the two lists to construct a complete Herb
Venn diagram of shared effective targets
A Venn diagram (http://bioinfogp.cnb.csic.es/tools/venny/index.html) was drawn to determine the shared target genes of BTW active ingredients and Salmonella Typhimurium infection. Finally, the shared targets were identified and visualized.
Protein–protein interaction network of target protein interactions
According to the Molecular Complex Detection (MCODE) algorithm in Metascape, the BTW target genes associated with Salmonella Typhimurium infection identified by the Venn diagram were input into the STRING database (https://string-db.org/cgi/input.pl) to identify the target proteins (Chin et al., 2014). In the protein–protein interaction (PPI) network, nodes (targets, compounds, pathways) and lines (relationships between targets and ingredients) were shown. Finally, the importance of nodes in networks was tested by the topological parameter “Degree,” which represents the number of links between lines and nodes (Wan et al., 2019).
Finding highly interacting gene modules and top 10 genes
To further visualize the PPI network results of the STRING analysis, we found highly interacting gene modules and the top 10 genes. The STRING analysis result is in the form of data in an Excel table, which can only form a preliminary visual PPI network. We used the Cytoscape MCODE plug-in to find the top gene modules (with K-Core set to 2) (Yu et al., 2017).
Target functional analysis
Gene Ontology analysis
We performed Gene Ontology (GO) enrichment analyses to explore the possible mechanisms of BTW against Salmonella Typhimurium infection by submitting shared targets to the database. The GO terms included biological process (BP), cellular components (CC), and molecular function (MF), and three components with p ≤0.05 were significantly enriched (Huang et al., 2009).
The Kyoto Encyclopedia of Genes and Genomes enrichment analysis
The Kyoto Encyclopedia of Genes and Genomes (KEGG, https://www.kegg.jp/) was used to describe molecular interactions and reveal known metabolic pathways (Kanehisa and Goto, 2000). The first 10 KEGG items were obtained based on gene ID numbers and p values. Then, the results of KEGG pathway enrichment analysis were put into the Cytoscape ClueGo plug-in (version 2.5.4) for visualization. (Li et al., 2018). An enriched pathway had a Fisher p value <0.05.
Molecular docking
AutoDock Vina was applied to visualize the docking of the active herbal compounds to the top 10 screened proteins. First, we analyzed the receptor and ligand separately. (1) The pdb crystal structures of the top 10 proteins were obtained from the PDB database (https://www.rcsb.org/) (Burley et al., 2017), and the docking active sites were determined. AutoDock Tools (ADT, version 1.5.6) was used to remove water molecules, isolate proteins, add hydrogen, and calculate the Gasteiger charges of each molecule for each ligand (Li and Fang, 2015). (2) The sdf structures of the compounds were obtained from the PubChem website (https://pubchem.ncbi.nlm.nih.gov/) and converted into pdb structures by ChemDraw software (ChemDraw, 2013). Second, we performed molecular docking via AutoDock Vina, with each protein undergoing three replicate docking with the compound. Finally, the result of the best binding energy score (affinity) was selected as the group representative. We used PyMOL (Version 2.5.4) software to visualize the molecular docking results (Seeliger and de Groot, 2010). The binding between ingredients and genes was visualized by the Biovia Discovery Studio visualizer (Wang et al., 2015).
In vitro experimental verification
The cellular thermal shift assay
The cellular thermal shift assay (CETSA) was used to detect the efficiency of menispermine and quercetin binding to the AKT1 protein in HeLa cells (Seeliger and de Groot, 2010, Jafari R et al, 2014). Menispermine and quercetin were purchased from Shanghai Yuanye Biotechnology Co., Ltd. (Shanghai, China, Lot: L23S31C125016, HPLC ≥98%; Lot: L23S31C125045, HPLC ≥97%). HeLa cells were seeded in 24-well plates at a density of 2 × 105 cells/mL. HeLa cells in the logarithmic growth phase at 18 h were collected and washed three times with cold PBS. Cells were resuspended in RIPA lysate buffer containing protease inhibitor cocktail (Beyotime, Shanghai, China) for 10 min. Cell lysates were prepared by centrifugation at 12,000 × g for 20 min at 4°C, and supernatants were collected. The protein concentration of the lysates was measured using the bicinchoninic acid (BCA) assay. The supernatant was equally divided into three fractions and treated with Dimethyl sulfoxide (DMSO), 50 µM menispermine, and 15 µM quercetin. After incubating at room temperature for 1 h, the three supernatants were divided into ten samples (60 µL each) and heated at different temperatures (25, 45, 56, 65, and 80°C) for 5 min, followed by cooling on ice for 5 min. The products were centrifuged at 15,000 rpm for 20 min at 4°C, and the precipitate and soluble fraction were separated in an ice bath. The supernatant was transferred to a new centrifuge tube and analyzed by SDS-PAGE, and Western blot analysis was performed using an AKT1 antibody (1: 1,000; ab81283, Abcam, United Kingdom).
The drug affinity responsive target stability assay
The drug affinity responsive target stability (DARTS) assay was performed according to a reported method with modifications (Huo et al., 2020). Briefly, see CETSA assay for the collection and cleaning of cells. The cell lysates were incubated with 10 × TNC buffer (500 mM Tris-HCl pH 8.0, 500 mM NaCl, 100 mM CaCl2) and mixed well. The lysates were equally divided into two tubes. DMSO was added to one tube, and the other was incubated with the drug for 1 h at room temperature. Five aliquots were prepared from each of the two protein samples, and the lysates were treated with 1:1000, 1:500, and 1:300 dilutions of pronase E (www.sigmaaldrich.com). They were kept at room temperature for 5, 10, 15, 20, and 30 min. Limited proteolysis was allowed to occur for 10 min at room temperature. Protease inhibitor cocktail (2× final concentration) was added to stop the digestion reaction. Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer (5×) was mixed with cell lysates and boiled at 100°C for 5 min. Each sample was separated by 10% SDS-PAGE and then subjected to immunoblotting analysis.
Statistical analysis
The data are expressed as mean ± standard deviation (mean ± SD). A two independent sample t test was used when the data satisfied normality distribution and homogeneity of variance. “*”Indicates a significant difference (p ≤0.05), “**”indicate a statistically significant difference (p ≤0.01), and “***”indicate a statistically significant difference (p ≤0.001) using the ordinary one-way ANOVA multiple comparison analysis.
Results
BTW exhibited suitable efficacy against Salmonella Typhimurium infection in vivo
BTW reduces the mortality rate of Salmonella Typhimurium-infected mice
Our experiments showed that the mortality rate was high in the Salmonella Typhimurium-infected mouse groups compared with the normal saline group (control group, C group) but that treatment with BTW at three doses slowed the mortality of infected mice. In the Salmonella Typhimurium infection groups, mice began to die on the fourth day after infection, and all mice died on Day 9. In the drug treatment groups, 4, 3, and 2 mice died after infection with survival rates of 70%, 70%, and 80% in the 3 different treatment groups (p ≤0.001), which indicated that BTW significantly reduced the mortality rate of Salmonella Typhimurium-infected mice (Fig. 1A).
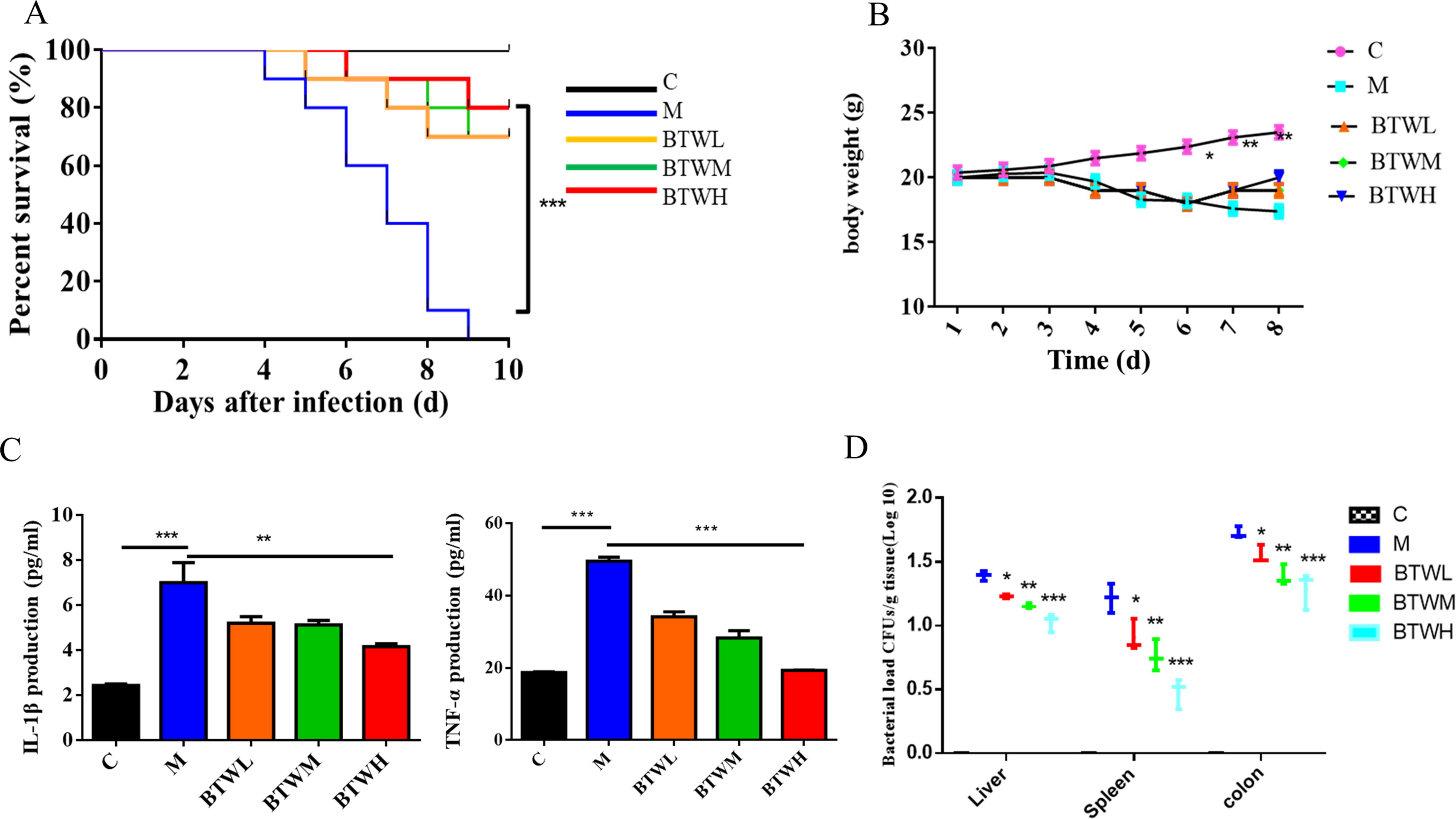
BTW exhibited suitable efficacy against Salmonella Typhimurium infection in vivo. (
BTW treatment significantly slowed weight loss caused by Salmonella Typhimurium infection
Mice in the Salmonella Typhimurium-infected group began to lose weight on the third day after infection, and the mice in the other groups started to lose weight on the fourth day after infection. The difference in weight loss from Day 6 onward was significant between groups, and BTW treatment significantly slowed weight loss caused by Salmonella Typhimurium infection (Fig. 1B, p ≤0.05).
BTW reduces cytokine levels in the colon tissues of Salmonella Typhimurium-infected mice
In this study, the levels of the inflammatory factors IL-1β and TNF-α (Fig. 1C) in the serum of mice were measured, and the results showed that the levels of the two inflammatory factors in the M group were significantly higher than those in the C group, while the levels of the IL-1β and TNF-α were significantly lower in the BTW treatment groups than in the model group (p ≤0.01, p ≤0.001). This result suggested that BTW has an anti-inflammatory effect on mice infected with Salmonella Typhimurium.
BTW reduces bacterial colonization numbers in the colon, liver, and spleen of Salmonella Typhimurium-infected mice
The competition index method was used to guide the experiment. After overnight incubation in a biochemical incubator at 37°C, the bacteria coating on LB plates was counted. We found that the colonization numbers of bacteria in the colon, liver, and spleen in the Salmonella Typhimurium-infected mice treated with BTW were significantly lower than those in the Salmonella Typhimurium-infected mice (p ≤0.05, p ≤0.01, p ≤0.001) (Fig. 1D), and this reduction was dependent on the concentration of BTW. This result showed that BTW has suitable efficacy against Salmonella Typhimurium infection.
BTW alleviates pathological damage caused by Salmonella Typhimurium infection in mice
BTW ameliorates pathological damage in the liver
The livers of mice in the C group were pink in color, with distinct lobulations, intact structures, and intact membranes, and the liver lobules of mice in this group had clear structures, uniform and neat arrangements of hepatocytes, no degeneration or necrosis, regular and neat arrangements of liver plates, and no obvious expansion or extrusion of the hepatic sinus. There were no obvious abnormalities in the door management area; no significant inflammatory changes were observed (see Fig. 2A).

BTW alleviated pathological damage caused by Salmonella Typhimurium infection in mice. (
The livers of mice in the M group were darker in color, and the edges were uneven to the naked eye, indicating that the lobulations were not clearly defined. A small amount of hepatocyte steatosis was observed, small vacuoles appeared in the cytoplasm (black arrow), more necrotic foci were observed, and a small number of hepatocytes steatosis were accompanied by a large amount of lymphocyte infiltration (blue arrow) (Fig. 2B).
The livers (Fig. 2B) of Salmonella Typhimurium-infected mice treated by BTWL, BTWM, and BTWH had a similar visual outlook to the C group. In addition to the cytoplasmic vacuoles (black arrow) visible in the liver of mice in the BTWL group and BTWM group, necrotic foci were occasionally seen, while the cytoplasm of the BTWH group was demineralized and lightly stained; necrotic foci were not common. The results showed that BTW has no toxic effect and can relieve the pathological damage caused by Salmonella Typhimurium.
BTW improves pathological damage in the spleen
The spleens of mice in the C group (Fig. 2C) were bright red and oval in shape and had an intact structure. The spleens of mice in the M group were dark red with obvious swelling. The spleens of mice in the BTWL group were dark red without obvious swelling, and the spleens of mice in the BTWM group and BTWH group were bright red in color and complete in structure.
Under 40× and 200× magnification (Fig. 2D) under electron microscopy, the splenic tissue of mice in the C group was composed of dense connective tissue with uniform thickness and rich in elastic fibers and smooth muscle fibers. The connective tissue of the splenic tissue extended into the spleen to form trabeculae without obvious abnormalities. The splenic parenchyma was composed of red and white pulp, and the white pulp consisted of the lymphatic sheath, lymph nodules, and the marginal area around the central artery. The layer structure was clear, and the number and size of each layer were not changed. The red pulp was distributed in the subcapsular area, around the trabeculae and outside the margin of the white pulp. It was composed of splenic cords and splenic blood sinuses. The distribution was uniform, and the white pulp was clearly demarcated. The spleen of mice in the M group (Fig. 2D) showed more dilatation of the medullary sinus (blue arrow). A large amount of neutrophil infiltration was seen (black arrow). However, the spleen of BTWL group mice occasionally showed medullary sinus dilatation (blue arrow). A small amount of neutrophil infiltration was seen (black arrow). No significant lesions were observed in the BTWM group except for a small amount of neutrophil infiltration (black arrow). Occasional neutrophil infiltration (black arrow) was observed in the spleen of mice in the BTWH group, and no significant abnormalities were observed in the rest of the mice. These results suggested that BTW can improve the pathological damage in the spleen caused by Salmonella Typhimurium infection.
BTW attenuates pathological damage in the colon
The colons of mice in the C group appeared normal, and the stool was dry, hard, and spherical (Fig. 2E). Colonic atrophy was observed in the model group, and the colonic length was shortened (7.3 ± 0.2 cm), which was significantly different from that in the C group (9.6 ± 0.1 cm) (p ≤0.001). There was obvious redness, swelling, and congestion and fecal deformity. Compared with that in the C group, the colon length (8.6 ± 0.1 cm, 8.7 ± 0.2 cm, 9.0 ± 0.1 cm) in the BTWL, BTWM, and BTWH groups was not significantly different, and the appearance of the colon was slightly but not significantly different.
The enteric inflammation induced by Salmonella Typhimurium was mainly in the anterior midsection. Colonic histological changes in mice treated with different concentrations of BTW were observed under a microscope (Fig. 2F). In the C group, the colonic mucosal epithelium was intact, goblet cells and crypts were intact, intestinal glands were abundant, and no obvious inflammatory changes were observed. The mucosal injury area of mice in the M group was apparent under a 40× objective lens. At 200× magnification, the intestinal tissue of mice in M group showed occasional erosion, a small amount of intestinal gland necrosis in the lamina propria, structural disappearance, connective tissue hyperplasia, and a small amount of focal infiltration of lymphocytes (black arrow). The mucosal epithelium of the BTWL and BTWM groups was intact, the proportion of goblet cells was slightly reduced, and there was a small amount of inflammatory cell infiltration. In the BTWH group, the colonic mucosal epithelium was intact, goblet cells and crypts were intact, and pathological scores were increased. We randomly selected 10 fields of view for each group of samples for histological index (HI) scoring. HI scores of edema, the proportion of polymorphonuclear granulocytes, goblet cell numbers, and epithelial damage in M group were significantly higher than those in the blank control group (p ≤0.001), while the HI scores of the BTWL, BTWM and BTWH groups were higher than those in the blank control group, but the difference was not significant. Moreover, with the increase in BTW concentration, the pathological index scores showed a downward trend. These results suggested that BTW can attenuate pathological damage in the colon caused by Salmonella Typhimurium infection.
Mechanism by which BTW combats Salmonella Typhimurium infection
Active herb compounds of BTW and their predictive targets
There were 465 compounds of BTW retrieved from the HERB website, including 115 in BTW, 179 in HB, 76 in HL, and 95 in QP. Forty-two overlapping compounds were removed, resulting in 423 identified compounds (Supplementary Table S1). We carefully searched and screened for the active compounds of the four ingredients of BTW. A total of 227 bioactive compounds were identified after Swiss ADME screening with RO5: 30 in BTW, 106 in HB, 51 in HL, and 40 in QP. There were 174 targets from PM and 204 targets from STP. We selected targets with a normal fit score ≥0.8 in PM and ≥0.1 in STP for follow-up analysis. Finally, a total of 911 potential BTW-related targets were identified (Supplementary Table S2).
Prediction of Salmonella Typhimurium infection-related targets
By searching disease databases, including CTD, DisGeNET, GeneCards, DrugBank, OMIM, and GEO, all Salmonella Typhimurium infection-related targets were finally summarized and obtained by removing duplicate targets. The number of disease-related targets obtained per database is shown in Figure 3. A total of 1,602 disease-related targets, including TNF, STAT3, JAK1, were considered potential therapeutic targets. This finding is illustrated in Supplementary Table S3.
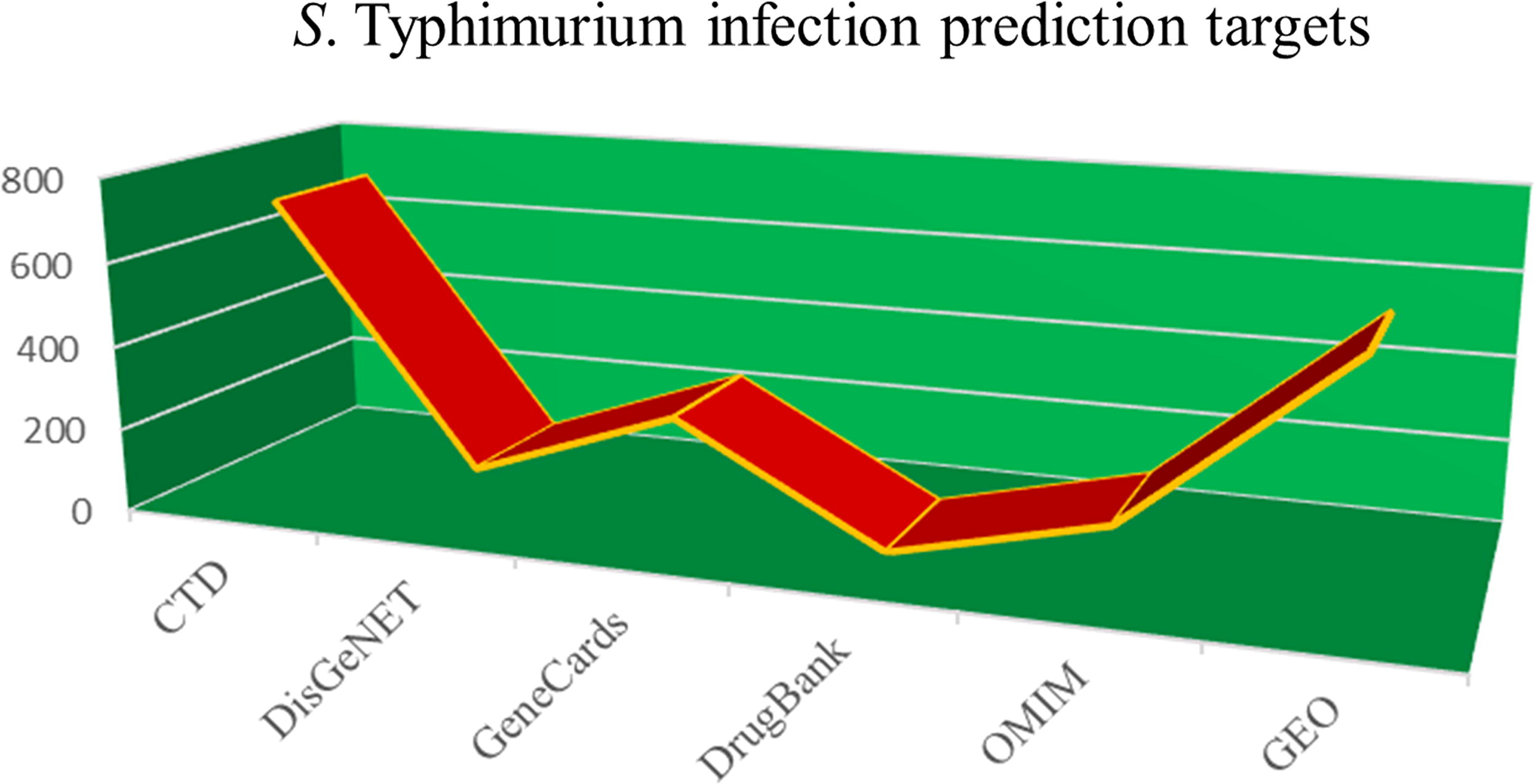
The number of Salmonella Typhimurium infection-related targets predicted by six databases.
Constructed Herb–Ingredient–Target network
The Herb

Herb–Ingredient–Target network. The green triangles represent the four ingredients of the Baitouweng decoction, the pink circles represent the compounds contained in the four ingredients, and the blue squares denote the disease-related target of the disease and the target of the compound.
Venn diagram of shared compound targets and disease-related targets
Comparing the 911 target genes of active compounds and 1,602 targets of Salmonella Typhimurium infection by Venn diagram analysis resulted in the identification of 188 genes as effective targets of BTW against Salmonella Typhimurium infection (Fig. 5, Supplementary Table S4). These shared targets were regarded as candidate targets of BTW against Salmonella Typhimurium infection (BTW-Salmonella Typhimurium).
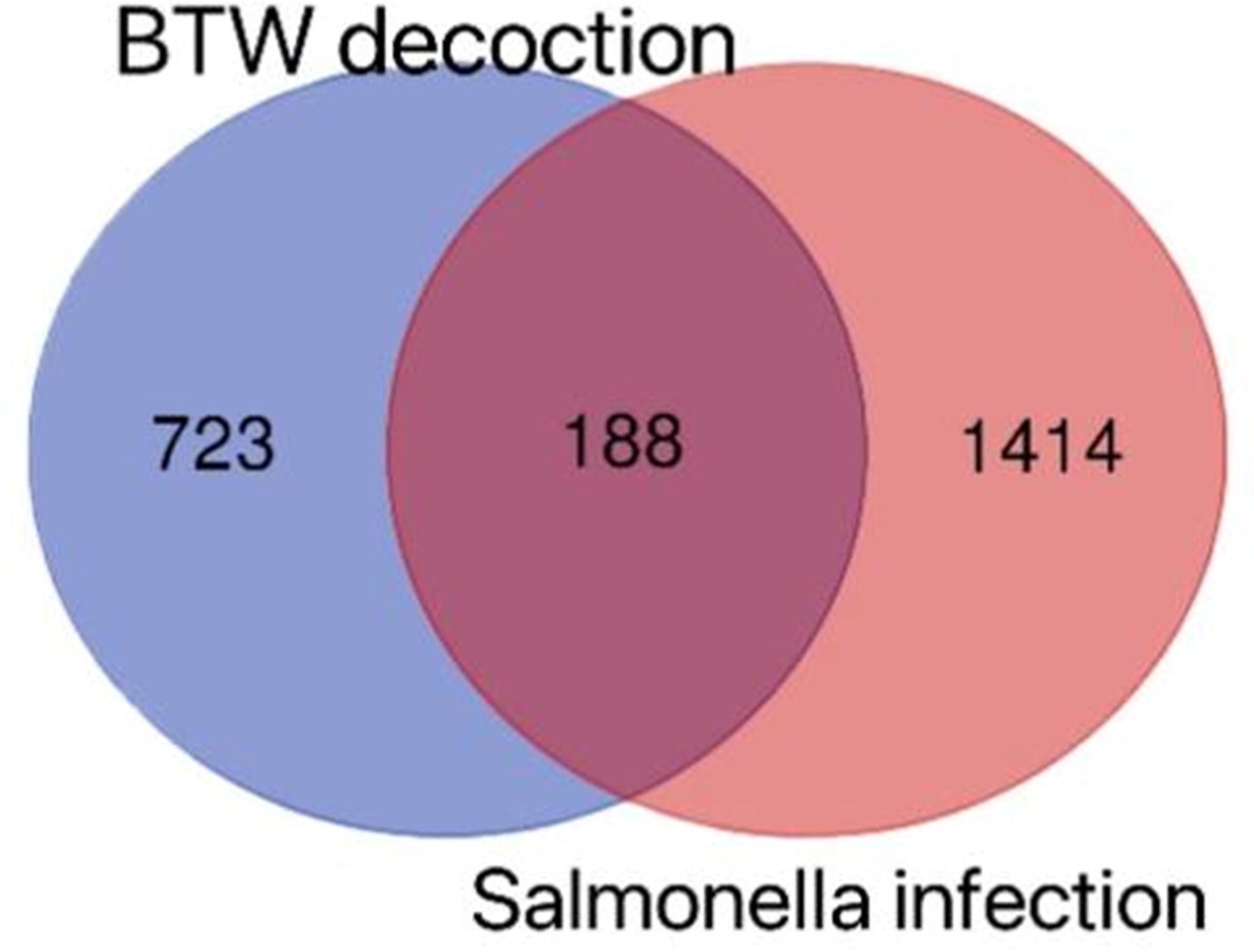
Venn diagram of the active compound target genes and disease-related targets.
PPI network analysis
PPI networks of 188 candidate targets of BTW-Salmonella Typhimurium were constructed by STRING 11.5 software with the species Homo sapiens, and the interaction score >0.40 as criteria. The PPI network is delineated in Figure 6, which contains 188 nodes (proteins) and 2,558 lines (protein–protein associations).
Target intersection and protein–protein interaction (PPI) network diagram.
Highly interacting gene modules and top 10 genes
Based on the 12 parameters of the key index in “Betweenness,” “BottleNeck,” “Closeness,” “Clustering Coefficient,” “Degree,” “DMNC,” “EcCentricity,” “EPC,” “MCC,” “MNC,” “Radiality” and “Stress,” the core nodes of BTW-Salmonella Typhimurium were obtained. We eventually chose the top 10 hub genes (Fig. 7a) that were major therapeutic targets of BTW-Salmonella Typhimurium, namely, tumor necrosis factor (TNF), AKT serine/threonine kinase 1 (AKT1), caspase 3 (CASP3), albumin (ALB), epidermal growth factor receptor (EGFR), Jun proto-oncogene (JUN), mitogen activated kinase-like protein (MAPK), signal transducer and activator of transcription 3 (STAT3), vascular endothelial growth factor A (VEGFA), and tumor protein p53 (TP53).
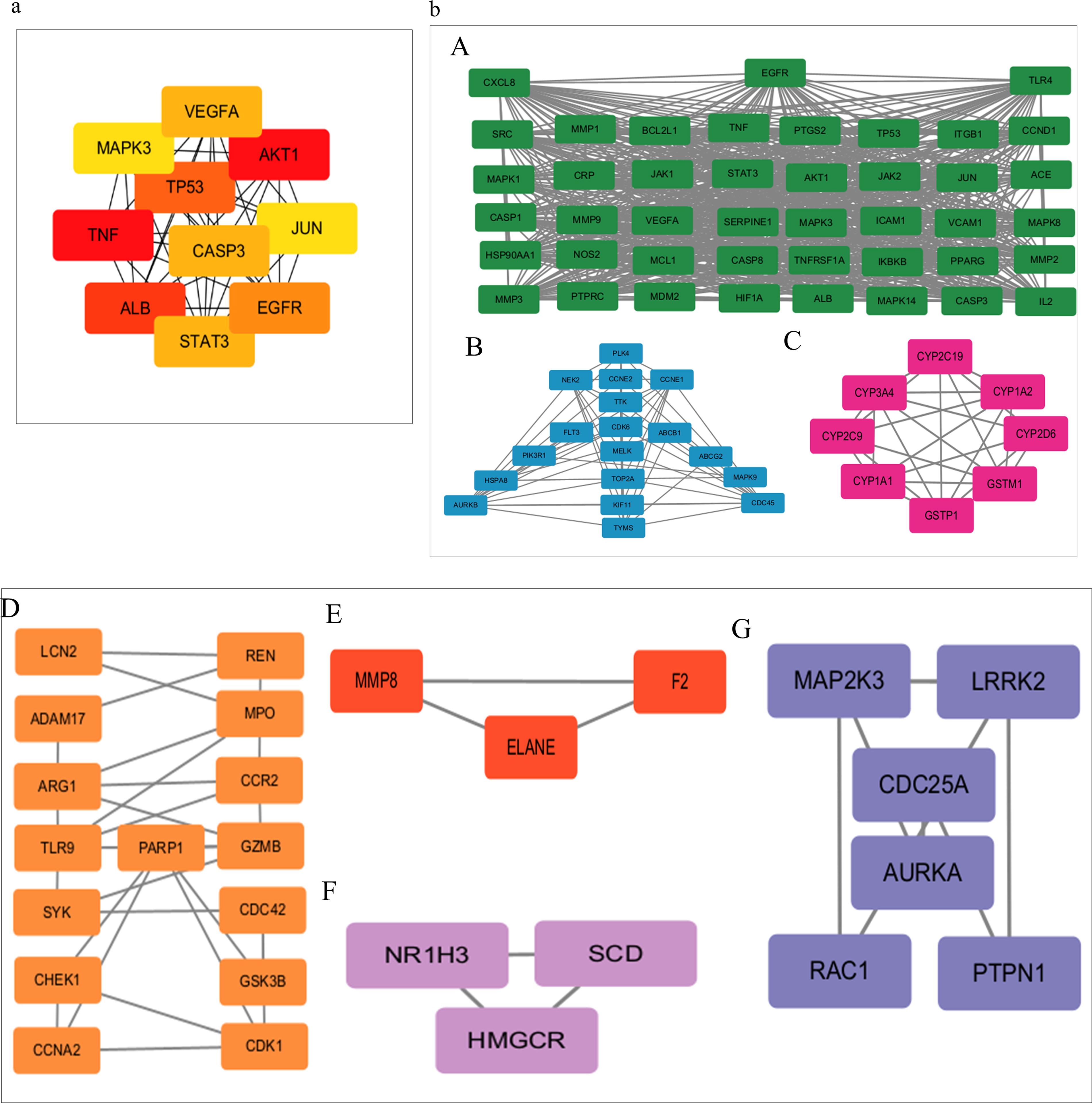
Highly interacting gene modules and top 10 hub genes.
Furthermore, the STRING analysis results were put into Cytoscape software in TSV format for further analysis, and the Cytoscape plug-in MCODE algorithm was used to analyze potential protein function modules, with the K-Core parameter set to 2. As shown in Fig. 7b, the seven clusters represent target proteins that have similar MFs and closely interact with one another. Among them, each cluster’s score, number of nodes and lines and involved proteins are shown in Table 1.
Seven Highly Interacting Gene Clusters
GO and KEGG pathway enrichment analysis
GO analysis
GO analysis results showed that 188 targets of BTW-Salmonella Typhimurium were enriched in many MFs, BPs, and cellular components (CC) (Fig. 8a A-C). Among them, the enriched MF was mainly kinase binding and monooxygenase activity; the enriched BP was mainly protein phosphorylation, response to inorganic substance, positive regulation of cell death, and response to bacterium; and enriched CC was mainly side of membrane, membrane raft, membrane microdomain, and vesicle lumen. A p value of < 0.05 was considered significant. These findings indicated that various BPs participated in the synergistic effects of BTW against Salmonella Typhimurium infection.
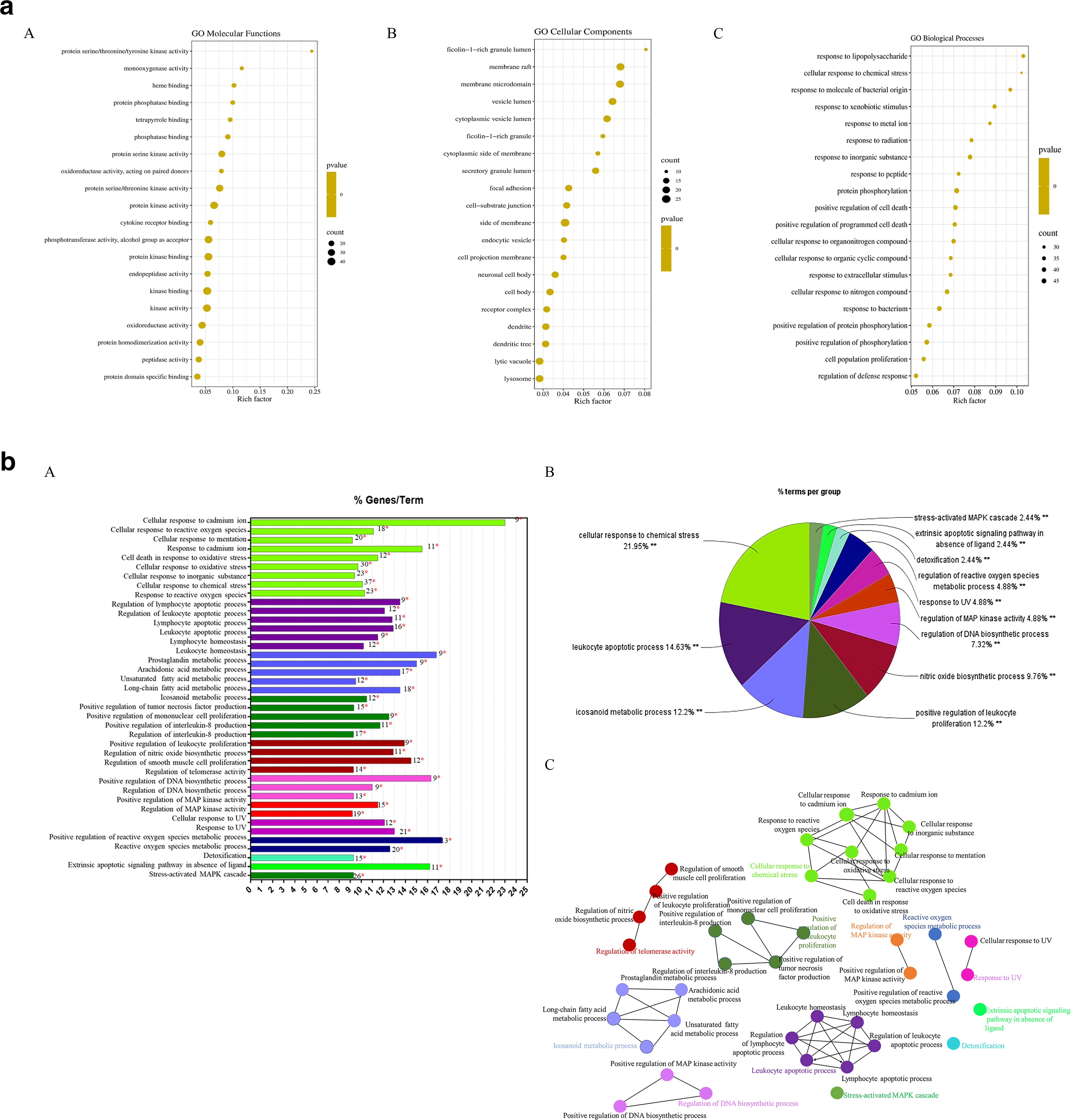
GO and KEGG pathway enrichment analysis.
KEGG pathway analysis
According to KEGG enrichment analysis by the Cytoscape ClueGo plug-in, 188 targets of BTW-Salmonella Typhimurium were divided into 41 clusters and included in 12 groups (Fig. 8b A), which were closely correlated to the cellular response to chemical stress, leukocyte apoptotic process, eicosanoid metabolic process, and positive regulation of leukocyte proliferation (Fig. 8b B). Furthermore, KEGG enrichment analysis demonstrated that BTW treatment affected 94 pathways (p < 0.05), including those involved in cell apoptosis, inflammation, and metabolism (Fig. 8b C).
Verification of the interaction of hub targets with active compounds by molecular docking
Through AutoDock Vina, the top 10 hub targets TNF, AKT CASP3, ALB, EGFR, JUN, MAPK, STAT3, VEGFA, and TP53 were docked with the 89 corresponding active compounds (Table 2). According to the best binding energy (affinity), each docking experiment was conducted in triplicate to finally select the optimal compound. The optimal structure of both the target gene and the optimal compound as the receptor and ligand is also presented in Table 2, all of which met the following conditions: the binding energies were <-4.25 kcal/mol (general binding activity), 90.9% of binding energies were <-5.0 kcal/mol (moderate binding activity), and 63.6% of binding energies were ≤-7.0 kcal/mol (strong binding activity). The 2D and 3D visualizations of the molecular interaction analysis of the top 10 proteins with selected compounds are shown in Fig. 9A and B.

A 2D and 3D visualization of molecular docking between receptors and ligands.
Acceptor and Ligand Results of Molecular Docking
In vitro experimental verification of the interaction of hub target genes with active compounds
CETSA verified that active compounds bind to hub target genes
To further verify the interactions of hub target genes with the corresponding active compounds of BTW, we evaluated the binding stability of selected hub target gene AKT1 and BTW active compounds menispermine and quercetin in HeLa cells by CETSA. As shown in Fig. 10A and 10B, the stability of the AKT1 protein decreased continuously with increasing temperature. The heat stability of the AKT1 protein in HeLa cells treated with menispermine and quercetin was significantly higher than that in HeLa cells treated with DMSO. This result suggested that AKT1 may be a direct target of menispermine and quercetin.
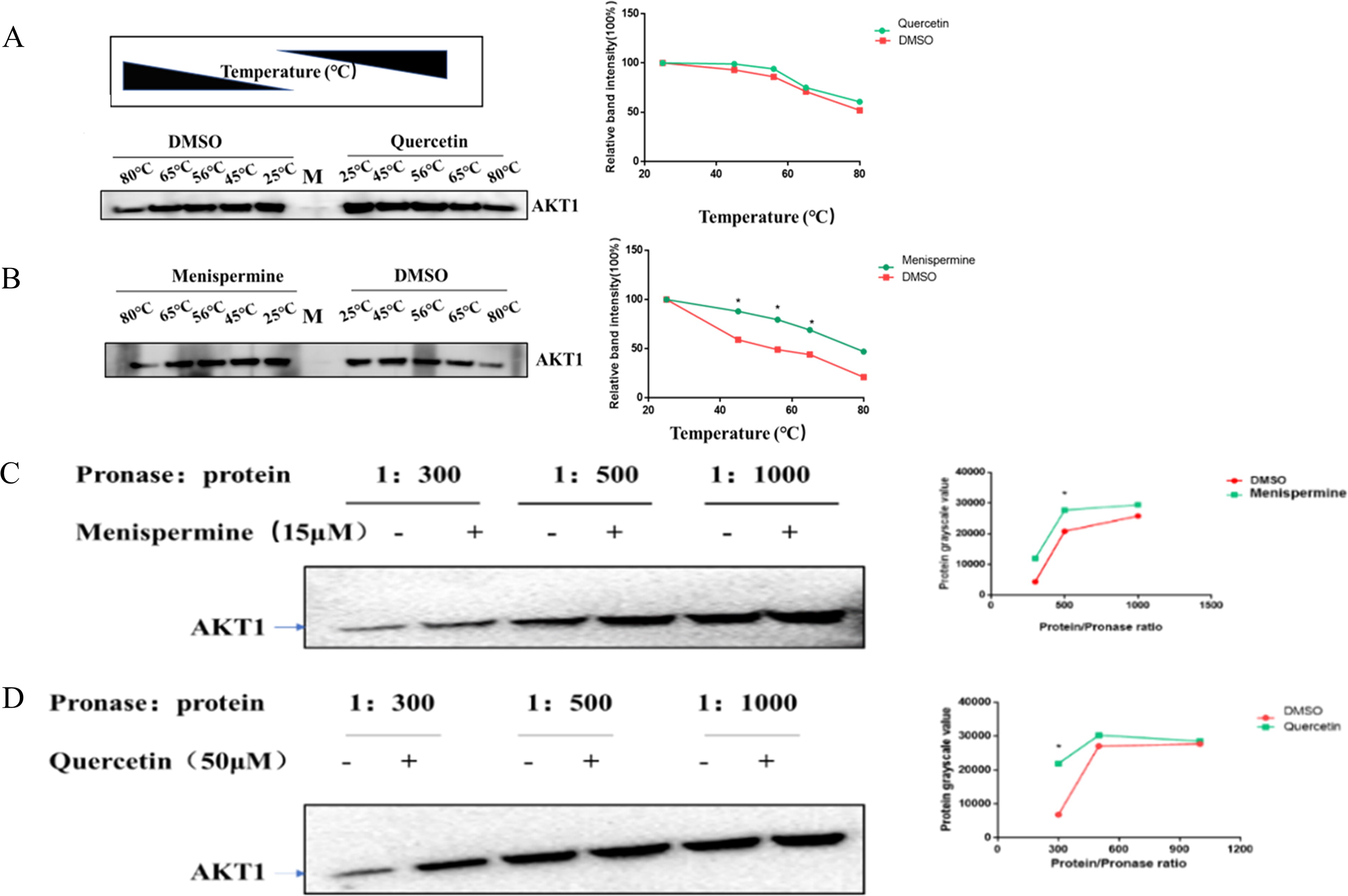
In vivo experimental verification.
DARTS verified that active compounds bind to hub target genes
Moreover, we also verified the binding stability of selected hub target gene AKT1 and BTW active compounds menispermine and quercetin in HeLa cells by DARTS assay. Under the gradient of pronase E-protein ratios of 1:300, 1:500, and 1:1000, the AKT1 protein degraded less in HeLa cells treated with quercetin and menispermine than in HeLa cells treated with DMSO (Fig. 10C, 10D). This result demonstrated that the interaction of AKT1 with quercetin and menispermine reduced the amount of protein degraded by pronase E. Thus, in vitro CETSA and DARTS assay verified the direct interaction of BTW active compounds with Salmonella Typhimurium-related targets.
Discussion
Pulsatilla chinensis is commonly used to treat cancers and inflammatory disorders in China. It has been illustrated that anemoside B4, a major ingredient of Pulsatilla chinensis, ameliorates Lipopolysaccharides-induced kidney and lung inflammation damage by inhibiting the proinflammatory NF-κB pathway in mice (Kang et al., 2019). The existence of the main drug (Pulsatilla chinensis) in BTW decoction makes BTW decoction play an important curative effect. Our results showed that BTW lowered the levels of inflammatory factors such as IL-1β and TNF-α. This phenomenon most likely involves the signal transduction between the active ingredients of BTW and the host target cells. Studies have shown that the balance between helper T (Th)17 and regulatory T (Treg) cells is also regulated by BTW, and BTW also decreases IL-1β, IL-6, and TNF-α levels and increases IL-10 levels (Miao et al., 2020). In addition, Hua YL et al. found that the improvement of DSS-induced ulcerative colitis in mice was due to the BTW-mediated regulation of pathways involving FXR and TGR5 by BTW, which affected the intestinal microbiota and bile acids (Hua et al., 2021). Our results showed that BTW has a mitigating effect on colon length and pathological damage in mice. BTW protects mice from Salmonella Typhimurium infection. This resulted in improved mouse weight and survival. It is worth noting that the anti-inflammatory and immunosuppressive effects of BTW were responsible for the reduction in the bacterial load of various immune organs in mice. However, more studies are needed to clarify the detailed mechanism of BTW action on Salmonella Typhimurium infection.
At present, studies focused on identifying specific targets to elucidate the mechanism of action of BTW on Salmonella Typhimurium infection. To better clarify the close relationship between the ingredients of BTW and Salmonella Typhimurium infection, we screened the highly interacting genes of BTW-Salmonella Typhimurium, including TNF, AKT1, CASP3, ALB, EGFR, JUN, MAPK, STAT3, VEGFA, and TP53, by network pharmacology. A finding emphasized that the tumor invasion of Salmonella enterica serovar Typhimurium is accompanied by strong hemorrhage promoted by TNF-α release (Leschner et al., 2009). In an inflammatory response, macrophages produce proinflammatory molecules such as TNF-α, IL-1β, and IL-6. BTW effectively reduced the symptoms and histopathological score of UC model mice, and it reduced the production of IL-6, IL-1β, and TNF-α (Chen et al., 2021). Furthermore, pulsatilla saponin D (PSD) significantly inhibited tumor growth in an HCC xenograft model and induced apoptosis by effectively suppressing the phosphorylation of factors downstream of PI3K, such as Akt, mTOR, and p70S6K, both in vitro and in vivo (Hong et al., 2012). C-Jun is a protein involved in the formation of transcriptional regulatory activator protein 1 (AP-1), which binds to homologous DNA. MAPK indirectly regulates the transcriptional activity of AP-1 by regulating the phosphorylation of c-Jun and the expression of JNKs and ERKs. Josely et al. indicated that SipA (T3SS-1-translocated effector proteins in Salmonella) induced chemokine expression through the phosphorylation of c-Jun and p38 MAPK (Figueiredo et al., 2009). Notably, the AKT1 target was a top target gene inputted in our GO and KEGG analyses. In recent years, Akt, which plays a role in the regulation of epithelial cell apoptosis in the intestines, has been considered an important prosurvival kinase by researchers (Zhang et al., 2004). A previous study revealed that sustained ERK activation and the subsequent upregulation of IL-8 expression were generated by the inhibitory effect of plasma membrane cholesterol depletion on Akt activation (Huang, 2011). Furthermore, EGFR is a transmembrane glycoprotein with intrinsic tyrosine kinase activity. After binding to the EGFR-binding ligand, the two complexes activate cell signaling. Bertelson et al. further reported that the Salmonella Typhimurium effector SopB can activate EGFR signaling in T84 epithelial monolayer cells through downstream PI3K signaling and pointed out that this activation may induce different actions than those observed in the EGFR pathway (Bertelsen et al., 2004). Mambu et al. found that the outer membrane expression of Salmonella Typhimurium and the target host cytoplasm membrane EGFR interacted with the protein Rck. This interaction activated the downstream signaling pathway, leading to internalization of Salmonella Typhimurium (Mambu et al., 2020). Studies have shown that PJ-34 protected intestinal epithelial cells from Salmonella Typhimurium-induced damage by upregulating IL-6 through the ERK and NF-κB pathways, while the P38 MAPK, JNK, and PI3K/Akt signaling pathways were not regulated by PJ-34 (Huang, 2009). In addition, Yin et al. demonstrated that the activation of the MAPK pathway and a series of inflammatory responses in HeLa cells infected with Salmonella Typhimurium were caused by the downregulation of ERK and FAK molecular phosphorylation by IpaJ (Yin et al., 2022). However, we still need to further explore the role of signaling pathways in host–pathogen interactions.
The binding energy score represents the binding affinity between the active ingredients and the target genes, and the lower the score is, the stronger their binding affinity is. The molecular docking results in silico showed that the binding score between EGFR and 6-(3-oxoindolin-2-ylidene) indolo (2,1-b) quinazolin-12-one derived from BTW was the lowest. Furthermore, it was found that 6-(3-oxoindolin-2-ylidene) indolo (2,1-b) quinazolin-12-one could stably bind the EGFR protein, and hydrogen bonds play a critical role at amino acid residues Lys-721 with a bond distance of 2.30 Å. Moreover, the binding score of 6-(3-oxoindolin-2-ylidene) indolo (2,1-b) quinazolin-12-one to CASP3 was −6.8 kcal/mol. ALB binding to dehydrotanshinone II A and AKT1 binding to quercetin, derived from HB and HL, respectively, had the same binding free energy score. The reason is that the AKT1 active sites to which the two compounds bind are not the same. For example, it was found that the binding sites of SDSS and AKT1 protein were the ASN-53, ARG-86, and lys14 residues in the PH domain of AKT1 (Gao et al., 2022). We further verified the interaction of AKT1 with active ingredients in vitro. The results of CETSA and DARTS showed that the interaction between menispermine, quercetin, and AKT1 was less affected by temperature and enzyme degradation in vitro. This finding may be attributed to the drug making the protein structure more stable. Wang RK et al. suggested that Biyuan Tongqiao granules greatly enhanced the heat stability of PI3K, AKT1, and ERK1/2 in nasal mucosa lysates (Wang et al., 2023). Furthermore, Peng et al. showed in a molecular docking experiment that dioscin binds to AKT1, Caspase3, TP53, C-JUN, and IL-6, and DARTS indicated that dioscin could bind to AKT1 (Xi et al., 2022). Similarly, the DARTS approach was used to test the relationship between periplocin and AKT (Lin et al., 2023).
Conclusions
In this study, BTW showed suitable efficacy against Salmonella Typhimurium infection by lowering the bacterial load of crucial organs and reducing the production of serum inflammatory factors. Network pharmacology analysis, in silico molecular docking technology, and in vitro binding analysis corroborated that BTW exerts anti-Salmonella Typhimurium effects through multicomponent, multitarget, and multipathway mechanisms (see Graphical Abstract), and the hub target gene AKT1 being a potential target of anti-Salmonella Typhimurium infection. It still needs to be further explored.
Footnotes
Acknowledgments
This work was supported by the National Key Research and Development Program of China (2021YFC2600200), the Science and Technology Research Project of the Jilin Provincial Department of Education (JKH20211179KJ; 2016444), and the Jilin Provincial Nature Science Foundation of Jilin Provincial Department of Science and Technology (20210101341JC). This work was also supported by the National Natural Science Foundation of China (No. 81801972).
Data Availability
The original data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researchers. The original data used to support the findings of this study are included within the Supplementary Information file.
Disclosure Statement
The authors declare no conflicts of interest.
Authors’ Contributions
C.X., W.Y., and Y.L. contributed to the conception and design of the study; C.X., W.Y., and Z.M. performed the experiments; S.J. and X.H. analyzed and interpreted the data; C.X. wrote the article. All authors contributed to article revision and approved the submission.
Funding Information
This work was supported by the National Key Research and Development Program of China (2021YFC2600200), the National Nature Science Foundation of China (32473029), the Science and Technology Research Project of the Jilin Provincial Department of Education (JKH20211179KJ; 2016444), and the Jilin Provincial Nature Science Foundation of Jilin Provincial Department of Science and Technology (20210101341JC). This work was also supported by the National Natural Science Foundation of China (No. 81801972).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.