
Abstract
Lactiplantibacillus plantarum is a member of lactic acid bacteria that improves the quality of fermented foods while also having a positive impact on human health. In this study, L. plantarum F2 was studied for characteristics such as biochemical and genetic identification, metabolite production, antimicrobial activity, and plasmid content. This strain exerts antimicrobial activity against some Gram-positive and Gram-negative pathogens (Listeria monocytogenes, Staphylococcus aureus, Salmonella, and Escherichia coli) with inhibition zone diameters ranging between 17.0 and 29.0 mm; it can ferment glucose, arabinose, galactose, lactose, and demonstrated the ability to grow at high temperature (50°C). Another physiological specification of the strain was the morphology of the isolate in selective medium, the de Man, Rogosa, Sharpe medium (MRS medium containing triphenyl tetrazolium chloride), which exhibits a chromogenic colony (characterized as purple colonies) on the modified-MRS (mMRS) medium. Metabolites such as lactic acid and diacetyl production of the strain F2 were also investigated using chromatography and found to be 10.07 and 0.05 µg/mL, respectively. The peptides of the isolate’s cell-free supernatant were determined to be ∼80 kDa, and finally, the plasmid isolated from the strain F2 was identified as L. plantarum strain KLDS1.0386 plasmid p4, which may be responsible for some characteristic properties, such as antimicrobial peptide production of the strain.
Introduction
Lactiplantibacillus plantarum (formerly Lactobacillus plantarum, L. plantarum) is usually found in many fermented foods (dairy, meat, fish, vegetable fermented products) enhancing food quality and desirable properties associated with health benefits as well (Corsetti and Valmorri, 2011). Fidanza et al. (2021) have currently cited L. plantarum as an ideal probiotic as it exhibits ecological and metabolic flexibility that allows it to adapt easily to various environments. This strain can be a good candidate for food preservation processes against food pathogens by using bacterial cells or metabolites produced by the bacteria.
L. plantarum is capable of producing bioactive substances such as exopolysaccharides, γ-aminobutyric acid, folic acid, and riboflavin, besides contributing to food safety via bacteriocin and organic acid production (Abdalla et al., 2021; Ayyash et al., 2020; Cui et al., 2021). This strain can have a high potential in food industry due to its properties and adaptability to various environments. The species belonging to L. plantarum may exhibit different physiological behaviors, and therefore, novel strains should be characterized to screen the potential usage of the strains. Lactobacillus genus is heterogeneous with more than 60 species and 33–55 ymol% G + C content. One-third of this genus’s members are known as strictly heterofermentative. Certain strains have had their genus changed due to displaying micro- and macromorphologies in addition to physiological and genetic profiles (Stiles and Holzapfel, 1997).
In this study, L. plantarum F2 was isolated and identified, and physiological and genetical characterization of the isolate was performed. The antimicrobial behavior of the strain and the origin of the antimicrobial activity were also investigated. This study introduces a novel strain to food processes as a starter or bioprotective culture, and the presented strain, L. plantarum F2, has the potential to be a probiotic as well.
Materials and Methods
Isolation and growth conditions of the bacteria
The strain F2 (isolated from undried tarhana, a traditional Turkish food, Project No. 119O343, Celik et al., 2024) was inoculated with a 1% inoculation rate in MRS broth De Man, Rogosa, Sharpe medium, Merck, Germany) and activated for 24 h at 30°C. For some experiments, the tryptone glucose yeast extract (TGYE) medium (tryptone 10.0, glucose 10.0, yeast extract 10.0, MgSO4 trace amount, MnSO4 trace amount, and Tween 80 1.0 mL) is also used for culture activation. The culture was activated for at least two passages before trials.
16S rRNA sequencing
Gram staining (Norateks, Catalog No. 106092), microscope imaging, catalase activity, and growing at 50°C were evaluated during the preliminary identification of the isolate in the study, and the 16S rRNA sequencing method was preferred for genetic identification (Sanger et al., 1977) at this stage. Genomic DNA was isolated according to the protocol recommended by the manufacturer, Qiagen DNeasy Blood & Tissue (Qiagen, MD, USA) DNA Isolation kit. Polymerase chain reaction (PCR) amplification of the 16S region was performed with universal primers (27Forward: 5′-AGAGTTTGATCCTGGCTCAG-3′; 1492Reverse: 5′-GGTTACCTTGTTACGACTT-3′). Representative 16S rRNA fragments of the different strains were sequenced with the Sanger sequencing technique (Sanger et al., 1977). The 16S rRNA gene sequences of the isolates were blasted for identification by using the National Center for Biotechnology Information database (http://www.ncbi.nlm.nih.gov/BLAST/).
The characteristics of F2 in the selective media
An mMRS medium was used as the selective media for distinguishing the F2 strain in the current study (Leuschner et al., 2003). Triphenyl tetrazolium chloride (TTC) 0.01% was added to the medium as a “color indicator” to allow chromogenic distinguishing of the colonies. The F2 culture was streak-plated on the mMRS agar and then incubated at 30°C for 24 h to screen the colony morphologies. P. pentosaceus ATCC 43201 was used as a reference strain in this test.
Substrate preference of the strain
The use of carbohydrates (L-arabinose, D-xylose, galactose, and lactose) was screened in this assay. Table 1 shows the usage of these carbohydrates by Lactobacillus curvatus, Lb. pentosus, Lb. sakei, Pediococcus acidilactici, P. parvulus, and P. damnosus. According to Table 1, glucose free-TGYE medium containing related substrate at 1% concentration was used to investigate the carbohydrate utility of F2. After incubation at 30°C for 24 h, the turbidity in the broth was accepted as positive.
The Comparison of Substrate Utility of Some Lactobacillus and Pediococcus Strains and F2 Strain

W: Varies according to subspecies.
Antimicrobial activity of the strain
The antimicrobial activity of the strain was investigated against Listeria monocytogenes ATCC 7644, Staphylococcus aureus ATCC 25923, S. aureus ATCC 43300, Salmonella enterica serotype Enteritidis ATCC 13076, Escherichia coli ATCC 25922, and E. coli ATCC 8339 with a two-stage experimental setup: (1) agar spot test and (2) well diffusion assay (Altuntas, 2011; Schillinger and Lücke, 1987). The active culture was directly used in the agar spot test, while the cell-free supernatant (CFS) of the culture was used in the well diffusion assay. The results obtained in the trials were evaluated through inhibition zones, which were classified as “strong,” “moderate,” or “weak” based on their size (strong: inhibition zones of >20 mm, moderate: inhibition zones between 10 and 20 mm, weak: inhibition zones of <10 mm; Aladeboyeje et al., 2022).
Lactic acid and diacetyl production of the strain
Determination and quantification of lactic acid by HPLC
The culture of F2 strain was centrifuged at 12,000 rpm for 20 min. CFS was obtained using a 0.22 µm cellulose acetate filter (Sartorius, Germany). The modified version of De Baere et al. (2013) was used as an extraction method. During the chromatographic separation, an XBridge C18 (4.6 × 250 mm, 5 µm) analytical column was preferred due to its high peak resolution and selectivity. Acetonitrile (A) and 15 mM phosphate buffer (pH 2.1, adjusted with 1M NaOH) (B) were used as the mobile phase. The International Council for Harmonization (ICH) guidelines were used for the validation of the developed method (ICH 2014). All results were calculated after three repetitive analyses.
Determination and quantification of diacetyl (2,3-butanedione) by HS-GC/FID
In this assay, 2,3-butanedione 97% (Sigma-Aldrich, Germany), and sodium chloride (Merck, Germany) were used for analysis. All gas chromatography (GC) experiments were performed using an Agilent (Santa Clara, USA) 6890 GC coupled with a flame ionization detector (FID), equipped with an HP-INNOWax (Bonded, crosslinked polyethylene glycol phase) GC column (30 m × 0.25 mm i.d. × 0.5 µm film thickness). The extraction step was carried out using an Agilent 7694E Headspace Sampler. Vials (10 mL), caps, and PTFE-coated silicone septa, from Agilent, were used in all headspace analyses. The samples were analyzed using the HS-GC-FID technique.
Heat resistance, peptide, and plasmid profile of the CFS of the strain F2
The supernatant of L. plantarum F2 was obtained by centrifugation at 6000 rpm for 10 min and then filtered through a 0.45 µm cellulose acetate membrane (Sartorious, Germany). The filtrate was treated at different temperatures (20, 37, 40, 50, 60, 70, 75, 80, 90, and 100°C) for 15 min to observe the activity after heat application. All the heat-treated samples were evaluated for antilisterial activity by comparing them to the control sample (nonheated sample).
The cell-free filtrate was fractionated by SDS-PAGE (4% deposition gel and 20% separation gel ratio) and 5× Laemmli buffer was prepared to load the samples. The gels were stained with oriole fluorescent gel dye after the run. The stained gels were washed with distilled water, and the BIO-RAD Versa-Doc imaging device was used by selecting Sypro Ruby tab with a 20.0-s exposure time.
Plasmid isolation from the bacterial cells was performed with the Fermentas GeneJet Plasmid MiniPrep Kit (#K0502). The isolated plasmid sample was investigated for the papA, papB, papC, and papD regions responsible for bacteriocin synthesis, which can be located on lactic acid bacteria (LAB) plasmids. PCRs were performed with two different primer combinations (forward 1: 5′-CACCATCTCCGGGCTTTCTT-3′ and forward 2: 5′-GCCAGGTTTCAAAAATTGACC-3′; reverse 1: 5′-GAACTGTCAGCACTCACCGA-3′ and reverse 2: reverse primer5′- CAGTTGTACCGTCAGCAG-3′). DNA sequencing was performed on the Applied Biosystems 3500 Genetic Analyzer (Thermo Fisher Scientific).
Results and Discussion
Isolation and identification of the bacteria
The strain F2 was determined as a Gram-positive, catalase-negative strain, and the strain was able to grow at 50°C, which is particularly characteristic for Pediococcus acidilactici species, which are a close relative of L. plantarum (Matejčeková et al., 2016).
16S rRNA sequencing results indicated that F2 was genetically similar to L. (Lactiplantibacillus) plantarum with a 99.93% similarity rate (Reference Accession No. MT613638.1). Fig. 1 depicts the phylogenetic tree of the strain created with the MEGAX Program (Kumar et al., 2018).
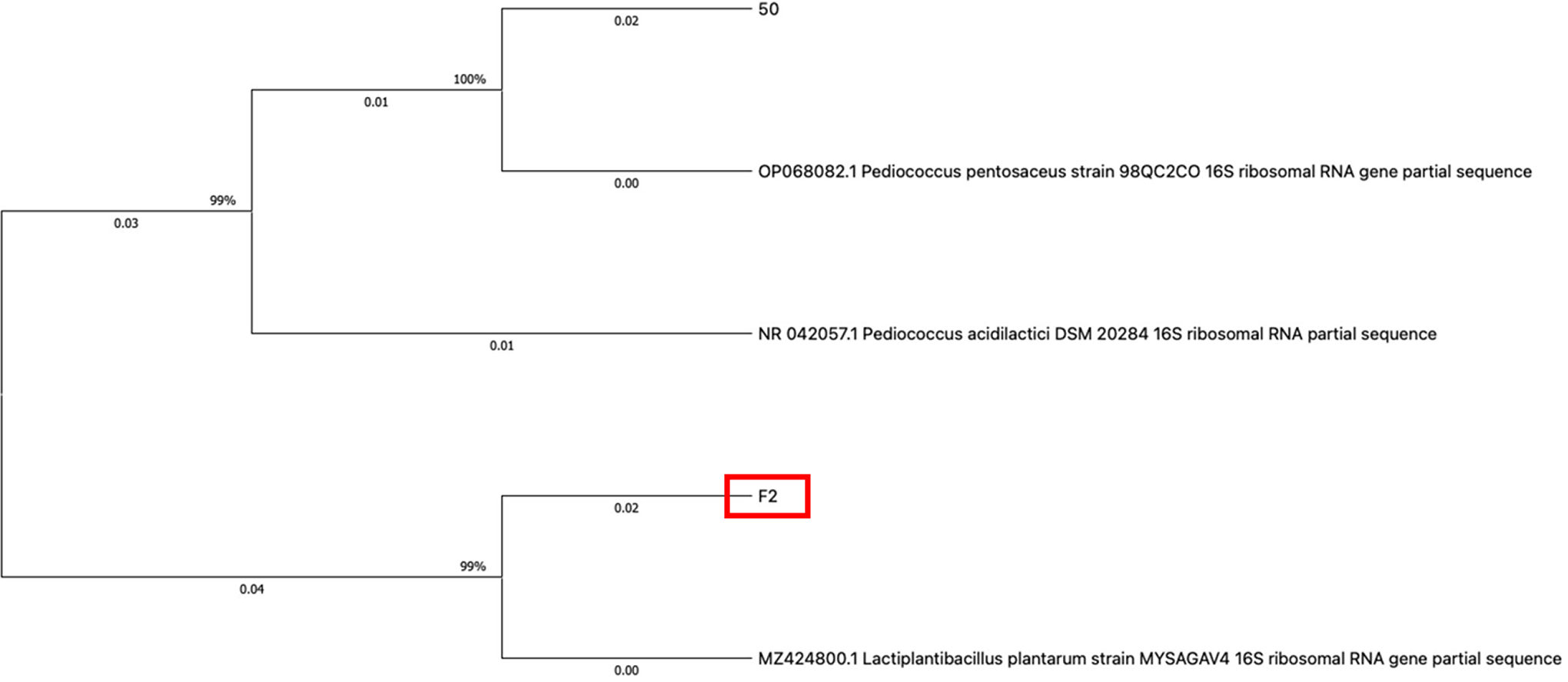
The phylogenetic tree of the strain F2 and reference strains; MZ424800.1 (Lactiplantibacillus plantarum), OP068082.1 (Pediococcus pentosaceus), and NR042057.1(Pediococcus acidilactici) created with the MEGAX Program.
Micro- and macromorphological and genetic identifications are generally important tools in bacterial identification systems. In some cases, it is necessary to contribute to this identification system with different supportive tools. The 50°C growth simple test is a very basic way to demonstrate the growth behavior of the bacteria at high temperatures. While Pediococcus acidilactici can grow at high temperatures such as 50°C, L. plantarum is not supposed to grow easily at this degree. A study performed to evaluate the growth dynamics of L. plantarum found that the cultivation temperature led to an increasing intensity of growth in the exponential phase for temperatures ranging from 8°C to 37°C but not for 38°C and 40°C (Matejčeková et al., 2016).
The behavior of the strains in selective medium and carbohydrate utility results
Several useful selective media are available for LAB characterized by specific properties associated with their adaptation to special environments (Schillinger and Holzapfel, 2003); Leuschner et al. (2003) and Simpson et al. (2006) reported using modified media for pediococci by adding cysteine hydrochloride, novobiocin, vancomycin, and nystatin to the MRS medium. This newly developed mMRS is supposed to provide a selective medium for pediococci by inhibiting the growth of other bacteria, yeasts, and molds. In addition, Leuschner et al. (2003) stated that the MRS chromogenic medium containing 0.01% TTC was used as a distinguishing “color” parameter by the formation of different colored colonies of pediococci from other LAB in TTC-containing media.
In the current study, the isolate was grown in MRS agar containing TTC, and the resulting colony images are shown in Fig. 2. Fig. 2 shows a noticeable difference; isolate F2 has a purple-colored colony morphology in TTC-MRS medium. To compare the results obtained in this experiment, P. pentosaceus ATCC 43201 was also grown in the modified media. White-colored colonies formed by P. pentosaceus ATCC 43201 were observed in TTC-MRS medium which is completely different from isolate F2.
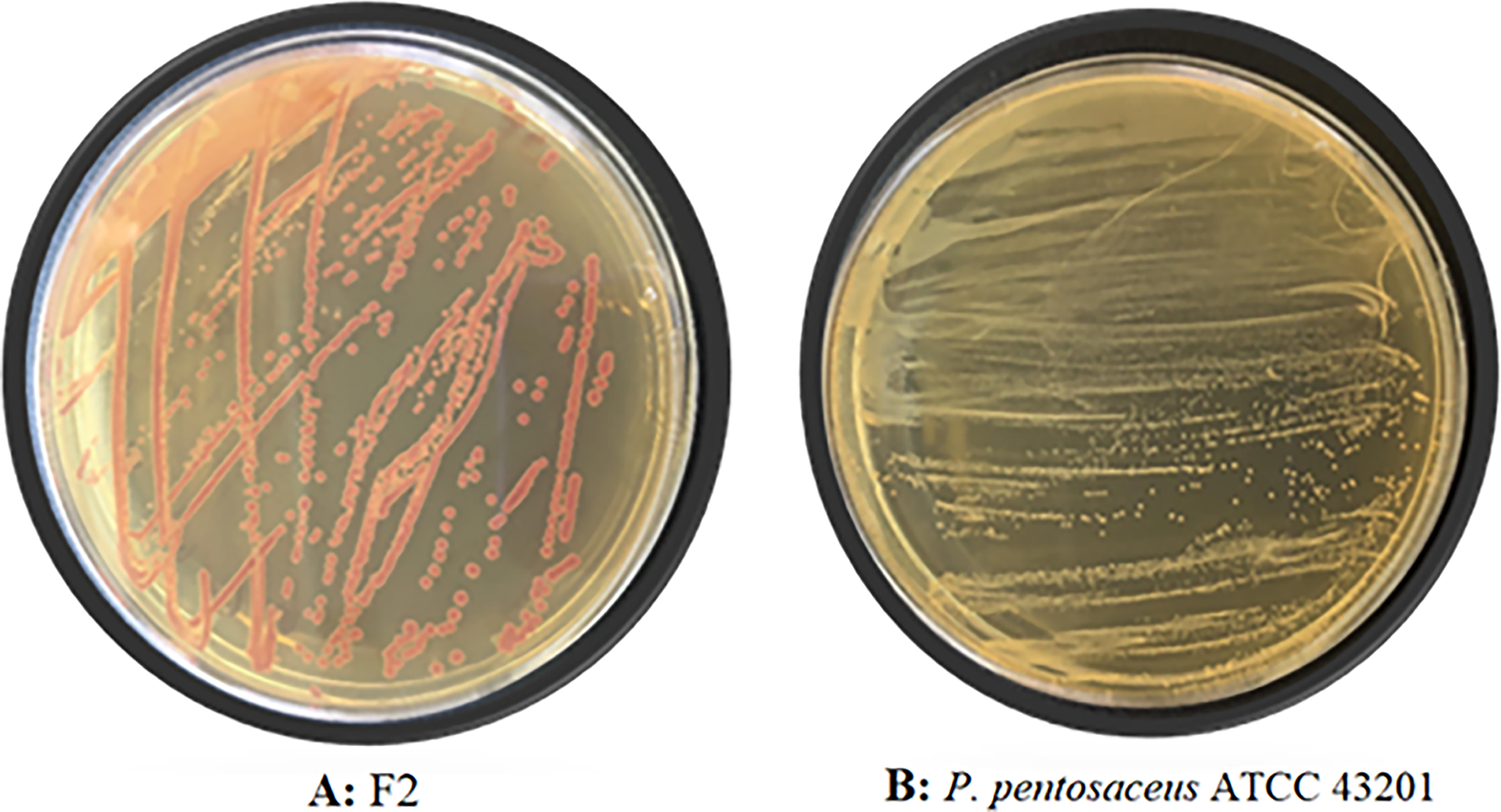
Colony morphologies observed in TTC-MRS medium of F2 and Pediococcus pentosaceus ATCC 43201. (
According to the results of the carbohydrate utility of the isolates, it was observed that isolate F2 fermented all the carbohydrate sources used in the assay such as glucose, arabinose, galactose, and lactose. The result was the same as the reference strain, P. pentosaceus ATCC 43201, which can grow when glucose, arabinose, galactose, and lactose are present in the medium. In this experiment, the substrate selection and comparison criteria were performed depending on the knowledge shared in the Materials and Methods section (Substrate preference of the strains) in Table 1, which shows the supposed results for P. acidilactici, P. parvulus, P. damnosus, Lb. curvatus, Lb. pentosus, and L. sakei (Abriouel et al., 2017; Cai et al., 1999; Kim et al., 2020; Mcleod et al., 2010; Santo et al., 2003). Contrary to the results obtained in this study, Bamigbade et al. (2023) observed that the two strains defined as L. plantarum (BLW103 and BLM102) could not ferment lactose and arabinose, while they were able to ferment glucose and galactose.
The antimicrobial activity spectrum and lactic acid/diacetyl production of the strain
To observe the antagonistic activity of the isolate F2, agar spot assay and well diffusion assay were performed against food pathogens. The isolate showed a wide antibacterial spectrum on the pathogens, according to the agar spot test. F2 had a 29 ± 1.41-mm zone-diameter average on L. monocytogenes, indicating a strong antimicrobial activity (Rota et al., 2008; Weerakkody et al., 2010; Zhang et al., 2016). It was observed that all the pathogens were sensitive to the F2 isolate. In the well diffusion experiment, the activity spectrum of the isolate was narrowed (between 11.5 and 16.0 mm), and the inhibition effect was totally lost on S. aureus ATCC 43300 (Table 2). In parallel with these results, Tarique et al. (2022) found that LAB were more effective on pathogens when they were directly used in the antimicrobial analysis. However, when postbiotic samples of LAB were used, the antimicrobial activity on pathogens was observed to decrease. Viable LAB were generally more effective on pathogens than the CFS. The antimicrobial activity of supernatant sample is associated with metabolites such as bacteriocins, organic acids, and volatile compounds. The active culture continues to produce metabolites while growing in the media, which may explain why the agar spot test exhibits more effective results compared with the well diffusion assay (Tarique et al. 2022).
Antimicrobial Activity Results of the Strain F2 on Some Pathogens

Strong activity.
Moderate activity.
Lactic acid is the main organic acid produced by LAB members. Therefore, it is supposed to be detected in the growth medium of these bacteria in different amounts. Unlike lactic acid, some metabolites, such as diacetyl, depend on the metabolic pathway of the strain (Reis et al., 2012). In the current study, lactic acid productions of the strains were examined by HPLC, and the F2 isolate was detected as the lactic acid producer strain with 10.07 mg/mL production. Diacetyl production was detected by the HS-GC/FID method; the F2 isolate produced this metabolite at a concentration of 0.05 µg/mL.
L. plantarum mainly produces lactic acid after fermentation processes and also synthesizes diacetyl as volatile component. However, the amount of these metabolites depends on the subspecies of this strain (Filannino et al. 2014). Lactococcus and some Lactobacillus perform homolactic fermentation, while Leuconostoc, Weissella, Oenococcus, and some Lactobacillus perform heterolactic fermentation. In the process of pure lactic acid fermentation, LAB use glucose as a carbon source to produce pyruvate through glycolysis, and then produce lactic acid under the action of lactate dehydrogenase. In heterolactic fermentation type, glucose can be decomposed into lactic acid, ethanol, and CO2 through the phosphoketolase pathway. On the other hand, diacetyl is produced by the metabolic intermediate α-acetolactate through oxidative decarboxylation (Wang et al. 2021).
Heat resistance, peptide and plasmid profile of the antimicrobial CFS of the strain
The bacterial supernatant of F2 was treated for 15 min at 20, 37, 40, 50, 60, 70, 75, 80, 90, and 100°C and the change in activity was followed up as a result of this temperature application. Fig. 3 demonstrates the obtained zone diameters compared with the control and depicts the obtained zones of inhibition.

The antimicrobial activity loss graphic of the cell-free supernatant of the strain F2 at high temperatures
In this preliminary test, only a 14% loss in activity was detected for isolate F2, even at 100°C, and the antimicrobial activity results obtained from almost all temperature experiments were close to the control sample. In general, the CFS of bacteria associated with bacteriocin production produced a positive result in the well diffusion assay. The strain F2 had antilisterial activity. Pediocin is a powerful antilisterial peptide among bacteriocins. It is a class II bacteriocin generally produced by pediococci, known as a small peptide (30–60 amino acids, molecular weight is lower than 10 kDa), and heat-stable up to 121°C (Raccach, 2014). The most significant characteristic of this peptide is its stability at high temperatures. The CFS of F2 showed antilisterial activity at high temperatures, even at 100°C, where the activity was saved up to 86%. Many bacteriocins produced by L. plantarum have been detected to date; especially, these bacteriocins have been classified in plantaricins. However, some of the plantaricins have a very similar operon structure to pediocin PA-1 (Todorov, 2009). Furthermore, pediocin AcH production by L. plantarum strains is reported by some researchers (Ennahar et al. 1996, Bernbom et al. 2009; Loessner et al. 2003).
This finding suggests the possibility that antimicrobial activity may be due to a peptide or peptides that are resistant to high temperatures. In this sense, bacteriocins are the strong candidates among all the bacterial metabolites. While there are many bacteriocin systems described in L. plantarum species, pln loci organized into different operons are the best-known L. plantarum strains that generally share the common feature of the plnABCD regulatory system and showed slight differences in the presence of the bacteriocin peptide genes. Such that pln genes are thought to be involved not only in the antagonistic activity of competing bacteria by biosynthesis of antimicrobial peptides but also in other bacterial responses (Ruiz-Larrea, 2011).
The CFS of the strain was investigated to screen the peptide content. The F2 strain was found to have antilisterial activity (14.25 mm inhibition zone diameter) and resistance to high-temperature degrees such as 100°C. Among bacteriocins, antilisterial activity and heat-stability are generally associated with pediocins and other class II bacteriocins, which revealed itself to be an indicator of pediocin or pediocin-like inhibitory substance biosynthesis of the isolate F2. SDS-PAGE image showed two bands on the gel, which are sized around 80 kDa (data are not shown). The strain F2 is identified as L. plantarum and the antimicrobial peptide produced by this strain is thought to be similar to class II bacteriocins, such as plantaricin or pediocin. Plantaricins can be divided into subgroups according to the characteristic properties. However, these peptides share a common property of being small-size peptides. Although there are peptides as small as 1.3 kDa among plantaricins, they are generally peptides of 3.5 kDa in size, and all known plantaricins are less than 5 kDa (Kareem and Razavi, 2020). Similarly, pediocin PA-1 is a small peptide with a molecular weight generally lower than 10 kDa. Differently from these peptides, pediocin PA produced by P. pentosaceus ATCC 43200 is determined to be 80 kDa in size, which is encoded by the pMD136 plasmid and belongs to Class III bacteriocins (large, heat-labile bacteriolysins) (Raccach, 2014).
Bacteriocins are generally encoded by bacterial plasmids. Due to the plasmid transfer system between relative strains, it is also common to isolate same or similar sequenced plasmids from isolates, particularly from relative bacterial strains. The pSMB74 plasmid is a well-known plasmid isolated from many LAB, although it is associated with pediocin AcH synthesis (pediocin PA-1). Another LAB plasmid, pSRQ11, also encodes pediocin PA-1 synthesis. pPLA4 is generally found in L. plantarum to encode plantaricin synthesis. Van Reenen et al. (2006) reported these three plasmid similarities in plasmid operons, but these plasmids can differ in DNA sequence apart from bacteriocin-related genes.
The strain F2 was discovered to be an antimicrobial compound producer with antagonistic activity against many foodborne pathogens. In this study, a bacterial plasmid larger than 10.000 bp was isolated (Fig. 4). Different primer combinations were used to search for the plasmid, and the amplicon obtained by PCR assays was sequenced.
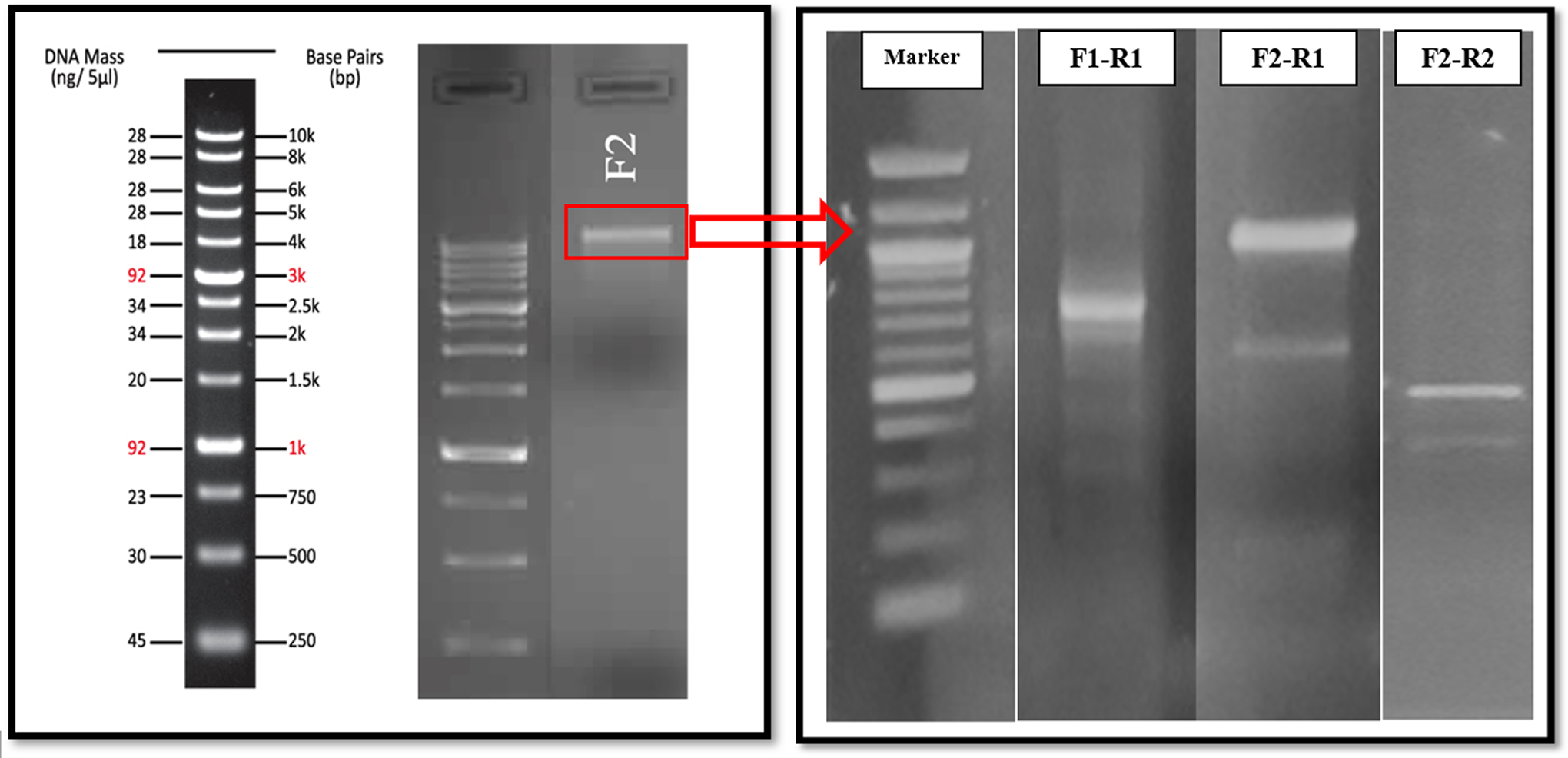
The agarose gel image of the bacterial plasmid of the strain F2 (Panel A) and the PCR amplicons, which are sequenced and used in determining the plasmid of the strain (Panel B).
A study by Miller et al. (2005) observed that P. parvulus ATO77, L. plantarum WHE92, and P. pentosaceus S34 plasmids had immune and synthesis system genes linked, and the promoter sequences were the same in the ATO77, pS34, and pWHE92 operons. In general, however, bacteriocin encoding genes can be located on a plasmid that is even differently named depending on the isolation source. In this study, the plasmid PCR product of the F2-coded bacteria was readable in the sequence reading of the PCR products. The region read matched with “L. plantarum strain KLDS1.0386 plasmid p4.” This strain is known for its acid and bile salt resistance, strong cell adhesion, lipid-metabolism regulation activity, and high-fat-induced obesity inhibition activity (Yilmaz et al. 2022). Furthermore, the L. plantarum KLDS1.0386 strain is known for its antioxidant capacity as well as the production of acetic acid and butyric acid, which can alter gut microbiota (Dong et al., 2022).
In conclusion, the isolate F2 was identified as L. plantarum as a result of 16S rRNA sequencing. Different experiments were carried out in this study, which determined that F2 belongs to this genus in terms of growth in selective media and the production of lactate and diacetyl metabolites. The bacteria exhibited a broad spectrum of antimicrobial activity, and the CFS, which could indicate bacteriocin production, was found to have an antimicrobial activity on foodborne pathogens. L. plantarum is a highly adaptive species that survives in a wide variety of habitats such as food, silage, and human gastrointestinal systems. In this sense, this bacterium can show its bioactivity from many different perspectives, from food fermentation processes to improving the gut health of humans. In this sense, the current study presents a fermented food isolate F2 as a strong bioprotective culture; investigating its behavior and probiotic properties in food systems is a necessity in terms of the application of this bacterium in the food industry.
Authors’ Contributions
Conceptualization: E.G.A. Analysis and data curation: E.G.A., A.A.C., B.S., K.C.T., M.G., H.M., and V.K.J. Funding acquisition: E.G.A. Methodology: E.G.A., M.G., H.M., and V.K.J. Investigation: A.A.C., B.S., and K.C.T. Supervision: E.G.A. Writing—original draft—review & editing: E.G.A. All authors have read and agreed to the published version of the manuscript.
Footnotes
Funding Information
This study was supported by the Scientific and Technological Research Council of Turkey (TÜBİTAK), grant no.: TOVAG-119O343.
Disclosure Statement
No competing financial interests exist.