
Abstract
Nontyphoidal Salmonella presents a significant threat to animal and human health as a food-borne infectious agent. This study focused on the characterization of Salmonella isolates obtained from fresh animal products consumed in Bejaia, Algeria. In total, 495 beef products, 600 chicken products, and 355 dairy products were collected from retail outlets and slaughterhouses in the region. Out of the 1450 samples collected, a Salmonella positivity rate of 3.5% (51/1450) was observed. Traditional Algerian sausages exhibited the highest prevalence of Salmonella contamination (14.1%, 36/256). Fifteen different serotypes were identified, with S. Kentucky (n = 12), S. Anatum (n = 11), and S. Bredeney (n = 9) being the most prevalent. Antimicrobial susceptibility testing showed resistance to diverse antibiotics, particularly against tetracycline, ampicillin, nalidixic acid, and sulphonamides. Whole-genome sequencing conducted on 21 isolates enabled the comparison of phylogenetic links between isolates. We notably identified clones circulating across the region in different locations and food types, suggesting contamination at the early stages of the food chain (in the herd or slaughterhouses) that disseminated to numerous butcher shops in various cities. We also identified acquired antibiotic resistance genes and point mutations in the quinolone resistance-determining region genes, contributing to the observed resistance patterns. This study describes the genomic and phylogenetic characteristics of several Salmonella clones found in diverse food samples in Algeria. It suggests potential transmission dynamics that could better understand Salmonella’s contamination routes.
Introduction
Nontyphoidal Salmonella (NTS) is a leading cause of food-borne infections worldwide, primarily resulting in gastroenteritis. Although the morbidity rate is high, mortality remains low except among vulnerable populations such as newborns and immunocompromised individuals (Fleckenstein et al., 2021). Annually, NTS is responsible for approximately 93.8 million cases of gastroenteritis and 155,000 deaths (Mezali et al., 2019). The infection is often linked to consuming contaminated food products such as poultry, eggs, meat, milk, and seafood (Djeffal et al., 2021). In Africa, livestock and poultry serve as key protein sources, and in Algeria, the expansion of intensive husbandry systems has increased Salmonella contamination risks, with animals acting as reservoirs (Mezali et al., 2019; Murinda et al., 2002).
NTS contamination can occur at multiple stages of food production, including feed processing and meat handling. Cross-contamination in butcher shops further exacerbates contamination risks (Teklemariam et al., 2023). Improving detection methods is critical, particularly as antimicrobial resistance (AMR) in Salmonella strains continues to rise. The inappropriate use of antimicrobials, especially in developing countries, has contributed to the emergence of AMR (Abreu et al., 2023; Moraes et al., 2024). Monitoring antimicrobial use and resistant strains in food animals is a crucial strategy to mitigate AMR spread (Almansour et al., 2023).
Whole-genome sequencing (WGS) offers a valuable tool for tracking the emergence of resistant Salmonella strains, yet limited studies in Algeria have utilized this method (Wan Makhtar et al., 2021). Our research aims to fill this gap by evaluating NTS’s prevalence, AMR patterns, and genetic characteristics in raw animal food products through WGS analysis.
Material and Methods
Sample collection
This study occurred in Bejaia province on Algeria’s northeastern Mediterranean coast. Sampling covered 31 of the 51 municipalities, and between May 2021 and July 2022, 1450 samples were collected from various sources, including butchers, retail markets, and slaughterhouses.
The samples represented a broad range of potential NTS sources, including red meat, cow and goat milk, traditional cheese, ice cream, eggs, and poultry carcasses. Samples were collected in sterile bags, labeled, and transported in ice boxes to the Microbiological Ecology Laboratory at the University of Bejaia, where they were analyzed within 6 h of collection.
Isolation of Salmonella strains
The isolation of Salmonella was performed following the standard cultivation method outlined in ISO 16140-6:2019 with slight modifications (Mooijman et al., 2019). For egg samples, a modified protocol based on ISO AFNOR 6579 was used, including surface disinfection with alcohol and iodine/potassium iodide solution, followed by cracking and homogenizing the contents of 10 eggs per sample (Freeman and Auer, 2012; McKeen, 2012). After pre-enrichment in Buffered Peptone Water (Fluka, St Louis, USA), selective enrichment was done using Rappaport-Vassiliadis broth (Fluka, St Louis, USA) and selenite cystine broth (Fluka, St Louis, USA) for poultry intestine samples (Hammack et al., 2001; Vassiliadis et al., 1985). Subcultures were grown on Xylose Lysine Decarboxylase (Fluka, St Louis, USA) and MacConkey agar (Fluka, St Louis, USA), followed by incubation at 37°C for 24 h.
Identification of Salmonella colonies was confirmed through MALDI-TOF mass spectrometry, and serotyping was conducted by slide agglutination according to the Kauffmann–White scheme (Grimont and Weill, 2007).
Antimicrobial susceptibility tests
All NTS isolates from raw food underwent antibiotic susceptibility testing to determine their resistance profiles (CLSI, 2020). Each isolate tested for a series of 17 antimicrobials agents, including: ampicillin (10 µg), amoxicillin/clavulanic acid (20 + 10 µg), ticarcillin (75 µg), piperacillin/tazobactam (30 + 6 µg), cefoxitin (30 µg), cefotaxime (30 µg), ceftazidime (30 µg), aztreonam (30 µg), imipenem (10 µg), ertapenem (30 µg), gentamicin (10 µg), tobramycin (10 µg), amikacin (30 µg), nalidixic acid (30 µg), ofloxacin (5 µg), tetracycline (30 µg), and trimethoprim/sulfamethoxazole (1.25 + 23.7 µg). The results were interpreted following the guidelines and recommendations from the CLSI (CLSI, 2020).
Whole-genome sequencing
In the selection process for WGS analysis, we carefully considered factors such as antibiotic resistance profiles, geographic locations of isolation, and the strains’ food origins. Due to resource constraints and funding limitations, we sequenced 21 strains from the pool.
The DNA extraction from the 21 Salmonella strains was performed using the Bacterial DNA Extraction kit on the SaMag automaton (Sacace Biotechnologies). The extracted DNA was generated using the Ion Xpress Plus Fragment Library kit on the AB Library Builder system (ThermoFisher Scientific). WGS was conducted on a Genestudio S5 sequencer (ThermoFisher), following the manufacturer’s instructions. To assess the quality of the raw fastq reads, FastQC v0.11.9 and MultiQC v1.14 were utilized. The reads were aligned against a Salmonella Enteritidis reference genome (GenBank accession number CP025554.1) using the bwa v0.7.17-r1188 and Samtools v1.15.1 algorithms with default parameters, and a minimum depth of 50X was found for all strains. Raw reads were assembled into contigs using SPADES v3.13.0 software. Fastq data are available in the Sequence Read Archive under bioproject PRJNA996503.
Serovar prediction was performed using SeqSero (Zhang et al., 2015), while multi-locus sequence typing (MLST) was carried out using the pubMLST database and analysis platform (Jolley et al., 2018). To identify core genomic single nucleotide polymorphisms (SNPs), Snippy v4.4.5 (available at https://github.com/tseemann/snippy) was used, with S. Enteritidis ATCC BAA-708 (NZ_CP025554.1) as the reference strain. FastTree v2.1.10 and FigTree v1.4.4 software were employed to construct a phylogenetic tree to visualize the relationships among the strains based on SNP comparisons.
Finally, in silico analysis was conducted on the assembled genome sequences to identify acquired antibiotic resistance genes (ARGs), mutations in the Quinolone Resistance Determining Region (QRDR) genes, and plasmid replicon types. The ResFinder 4.1 database (accessible at https://cge.cbs.dtu.dk/services/ResFinder/) was used to detect ARGs, configured with a 90% identity and 80% coverage threshold.
Results
Prevalence and distribution of Salmonella strains
Among the 1450 food samples, 51 (3.5%) tested positive for Salmonella, indicating its presence in the studied area (Supplementary Table S1). Salmonella identification was confirmed with MALDI-TOF scores >2.00 for all the isolates. Of the 51 positive Salmonella isolates, 41 were identified in livestock samples. Specifically, 36 isolates (14.1%) were obtained from 256 traditional Algerian sausage samples screened, 4 isolates (1.7%) from 239 raw red meat samples screened, and 1 isolate (1.5%) from 65 ice cream samples screened. The remaining 10 Salmonella isolates were detected in poultry samples, with 6 isolates (5.5%) from 110 raw poultry meat samples, 3 isolates (2.0%) from 153 gizzard and intestines samples, and 1 isolate (0.6%) from 167 offal samples. No Salmonella was isolated from eggs, milk, traditional cheese, or chicken carcasses (Supplementary Table S2).
Antibiotic susceptibility of NTS isolates
Out of the 51 Salmonella isolates, 9 (17.6%) were pan-susceptible (susceptible to all antibiotics tested), while the remaining 42 strains showed resistance to one or more antibiotics. Of the 51 isolates, 34.6% exhibited resistance to three or more antimicrobial agents, classifying them as multidrug-resistant strains.
The resistance rates varied depending on the antibiotics. The tested strains exhibited a higher resistance rate against tetracycline and nalidixic acid, with a resistance rate of 54.9% and 51.0%, respectively. Ampicillin and ticarcillin exhibited the same resistance rate of 35.3%, while amoxicillin/clavulanic acid showed resistance in only 3.9% of the isolates. Gentamicin demonstrated a resistance rate of 7.8%, while trimethoprim/sulfamethoxazole and ofloxacin displayed 21.6% and 29.4%, respectively.
Serovars and ST identification
Among the 51 Salmonella isolates, 15 different serovars were found: S. Kentucky (n = 12), S. Anatum (n = 11), S. Bredeney (n = 9), S. Hadar (n = 4), S. Virchow (n = 3), S. Altona (n = 2), S. Typhimurium (n = 1), S. Liverpool (n = 1), S. Sundsvall (n = 1), S. Duesseldorf (n = 1), S. Indiana (n = 1), S. Kedougou (n = 1), S. Ohio (n = 1), S. Souza (n = 1), S. Paratyphi A (n = 1), and one strain was Non typable (Supplementary Table S2).
WGS performed on 21 isolates, confirmed the serovar identifications, and enabled to correlate serotypes with serovars: ST306 with S. Bredeney, ST198 and S. Kentucky, ST33 and S. Hadar, ST16 and S. Virchow, ST64 and S. Anatum, ST19 and S. Typhimurium, ST1959 and S. Liverpool, and finally ST488 and S. Sundsvall. Phylogenetic relationships between isolates based on the SNP differences are presented in Figure 1. All sequenced isolates from the same serovar clustered. The SNP number ranged from 50 to 211 depending on the serovar, underlining the solid genetic relationships between isolates from the same serovar (Supplementary Table S3).
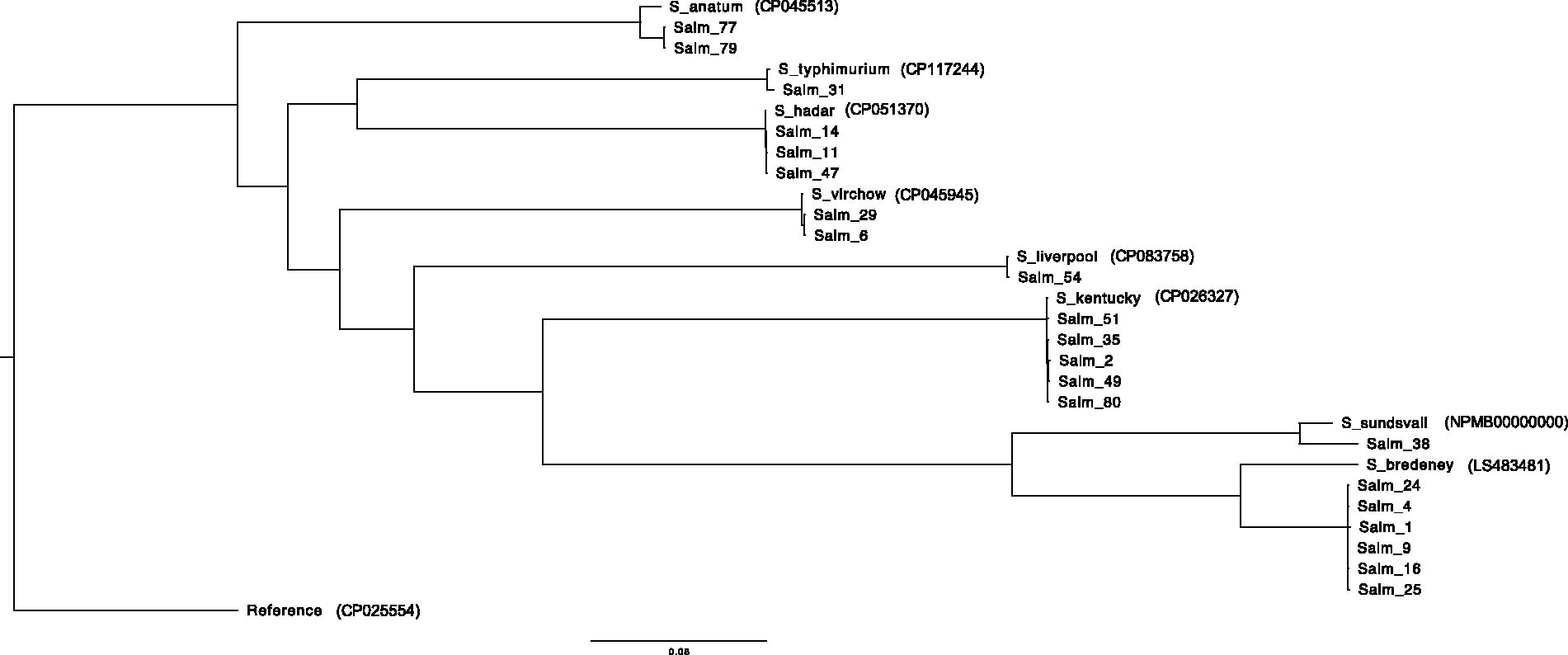
Phylogenetic tree based on SNP differences of the 21 sequenced Salmonella isolates. Reference genome Salmonella Enteritidis (GenBank accession number CP025554) was used to calculate phylogenetic distances between isolates. Salmonella Anatum CP045513, S. Bredeney LS483481, S. Hadar CP051370, S. Kentucky CP026327, S. Liverpool CP083758, S. Sundsvall NPMB00000000, S. Typhimurium CP117244, and S. Virchow CP045945 were used to confirm serovar identification. Branch lengths represent the number of nucleotide substitutions per site (scale: 0.08 substitution per site). SNP, single nucleotide polymorphisms.
Resistome
Various ARGs were identified among the 21 sequenced isolates, contributing to resistance against different classes of antibiotics (Table 1).
Characteristics of the 21 Sequenced Salmonella Strains
B, Butcher; SL, Slaughterhouse.
AMC, amoxicillin-clavulanic acid; AMP, ampicillin; GEN, gentamicin; NAL, nalidixic acid; OFL, ofloxacin; SXT, trimethoprim/sulfamethoxazole; TET, tetracycline; TIC, ticarcillin.
The chromosomal aac(6’)-Iaa gene, encoding a chromosomal aminoglycoside acetyltransferase, was present in all 21 strains. Other aminoglycoside resistance genes included aph(6)-Id (n = 6), aadA7 (n = 3), aac(3)-Id (n = 3), aadA2 (n = 2), and aph(3”)-Ib (n = 1), suggesting resistance through various enzymatic modifications. The bla TEM-1B gene was found in 12 strains and conferred resistance to amoxicillin and ticarcillin. Tetracycline resistance was conferred by the tet(A) gene (n = 12). Sulfonamide resistance was mediated by sul3 (n = 5), sul1 (n = 5), and sul2 (n = 1) genes, while resistance to trimethoprim was associated with dfrA14 (n = 5), dfrA12 (n = 2), dfrA1 (n = 1), and dfrA5 (n = 1) genes. Macrolides’ resistance mph(A) gene was found in two isolates, and the florfenicol resistance gene floR in 1 isolate. Finally, the quaternary ammonium qacE gene was present in five isolates.
Quinolone resistance was mediated either by plasmid-mediated quinolone resistance genes, including qnrS1 (n = 5) and qnrB19 (n = 1), or by different mutations in the QRDR of the gyrA or parC genes. Four different mutations were observed within the gyrA subunit: gyrA D87N (n = 8), D87G (n = 2), D87Y (n = 2), and S83F (n = 5), whereas parC T57S and S80I mutations were present in 18 and 5 cases, respectively. S. Kentucky isolates exhibited four mutations in their QRDR, including gyrA S83F, gyrA D87N, parC T57S, and parC S80I.
Discussion
A global study conducted between 2000 and 2017 across 29 countries reported a Salmonella prevalence of 9.5% in apparently healthy cattle (5,010 Salmonella-positive cases). This highlights the varying prevalence of Salmonella contamination depending on the region, type of sample, and study context (Gutema et al., 2019). In the Middle East and North Africa (MENA) region from 2011 to 2021, a prevalence of 13.5% for NTS was reported from food (Bellil et al., 2023).
The highest prevalence of Salmonella in our study was in the traditional Algerian sausages (14.1%). However, it was lower than previously reported (Ed-dra et al., 2017), with 43.7% beef sausage and 23.5% poultry sausage (Mezali and Hamdi, 2012). In Algeria, the prevalence of beef and poultry sausages varied greatly depending on the food type and the studied region (Djeffal et al., 2021). For traditional sausages, a 13.2% Salmonella prevalence was reported in MENA countries (Bellil et al., 2023), 5% in Morocco (Amajoud et al., 2017), 3.9% in the Reunion Island (Hamed et al., 2021), 1.5% in Turkey (Büyükünal et al., 2015), and 0.0% in Egypt (Morshdy, 2018). These results indicate that the prevalence of Salmonella in sausages is higher than in other sample sources. Contamination can be attributed to the raw materials or improper handling of guts and ground meat during preparation. These products pose a consumer risk due to undercooking trends in Algeria (Hachemi et al., 2019).
In our study, the prevalence of Salmonella was lower in other meat products (0% and 5.5%), 0.65% and 2.0%, respectively, in chicken internal organs (offal) and intestines and lower than those reported in chicken internal organs from MENA countries (12.5%) (Bellil et al., 2023).
We did not detect Salmonella in milk samples, poultry carcasses, and eggs, as in three other Algerian investigations (Adjlane-Kaouche et al., 2014), unlike reports for carcasses in Algeria (1,3%) (Ayachi et al., 2015) in eggs in China (0.5%) (Li et al., 2020), and from eggshells in Iran (Moosavy et al., 2015). Our results and findings demonstrate a favorable situation regarding the absence of Salmonella in these samples.
Our study showed that several serovars were involved in food contamination, with S. Kentucky, S. Bredeney, and S. Anatum being the most prevalent. These three serovars are commonly associated with foodborne outbreaks in different countries (Bellil et al., 2023). They are often considered the most prevalent serotypes in human and animal clinical isolates (Ferrari et al., 2019). S. Kentucky is an emergent human pathogen, even in developed countries (Neelambike et al., 2023), and is frequently associated with multidrug resistance (Turki et al., 2012). It was first reported from a chicken in the United States in 1937 (Edwards, 1938). It was the third most prevalent serotype in the MENA region (Bellil et al., 2023).
The range of SNP differences between the isolates of the same serovars was low in our study, spanning from 50 to 211 SNPs. These isolates were found in various types of meat and locations. Thus, specific clones appear circulating in different butcher shops in Bejaia (same serovar, same ST, with few SNPs). The same Salmonella clone might be transmitted at the beginning of the food chain, either in the herd or the slaughterhouses, then spread to different areas. These results highlight the use of WGS to precisely detect clones along the food chain and better understand Salmonella clones’ transmission dynamics and evolution.
We observed various levels of resistance to antimicrobial agents among Salmonella isolates, as in France, Belgium, Slovak Republic, Morocco, Ethiopia, and MENA countries (Abo Hashem et al., 2022; Oueslati et al., 2022). The percentage of multidrug resistance isolates MDR strains in this study (34.6%) is higher than in South Dakota (Erickson et al., 2018), lower than in Egypt, and similar to previous reports in Algeria (32.2%) (Sallam et al., 2014). Genetic analyses of Salmonella isolates identified several antibiotic-resistant genes and chromosomic mutations. Penicillinase gene bla TEM-1B was present in four serovars (S. Bredeney, S. Kentucky, S. Sundsvall, and S. Typhimurium) and was the unique beta-lactam gene detected in this study. Quinolone resistance was mainly mediated by mutations in the gyrA and parC genes and the qnrS1 and qnrB12 genes in the S. Bredeney and S. Liverpool serovars, respectively. Several mutations in the gyrA and parC genes were associated with fluoroquinolone resistance, in agreement with the literature (Chang et al., 2021). Resistance profile varied between isolates, even within the same serovar. This is mainly due to the acquisition or loss of plasmids carrying AMR genes.
Our findings underscore the importance of adopting a One Health approach to fight AMR, addressing the collaboration between animal, environmental, and human health sectors. AMR is a significant public health threat, and revising food hygiene regulations is crucial to mitigate the spread of multidrug-resistant Salmonella (Pinto Ferreira et al., 2022). Genomic investigations can provide further insights into the characteristics and phylogenetic links between isolates, thus determining if similar trends are occurring in Algeria. Implementing monitoring and prevention measures has, for example, successfully eradicated Salmonella pathogens, particularly in poultry and egg production. Good agricultural practices at the farm level and proper manufacturing and hygiene practices in slaughterhouses or during vegetable processing are essential to minimize Salmonella contamination in cattle meat (Andres and Davies, 2015).
Conclusion
Our study revealed a moderate prevalence of 3.5% Salmonella in various food-animal products, but mainly in traditional Algerian sausages (14.1%), indicating inadequate hygiene practices in Bejaia, Algeria. Here, we hypothesized that Salmonella contamination may have occurred at the animal herd level or the slaughterhouses and spread to the butcher shops in various cities. A complete genomic analysis is needed to understand Salmonella transmission dynamics and evolution in food. High resistance rates to ampicillin, nalidixic acid, tetracycline, and sulphonamide were observed. The association between food-animal products and multidrug resistance underscores the urgent need for a collaborative, multinational approach to address AMR on a global scale.
Footnotes
Authors’ Contributions
Z.B. was responsible for sample collection, analysis, and article writing. S.M. conducted whole-genome sequencing, analyzed the data, and corrected the article. V.T. contributed to the bioinformatic analysis. A.M. analyzed the microbiological data. C.D.C. conducted antibiotic testing and also made corrections to the article. O.B. supervised, reviewed, and edited the article. A.T. provided guidance and supervision and made corrections to the article. All authors have read and approved the final version of the article.
Author Disclosure Statement
The authors have no affiliations or financial involvement with any organization or entity that has a financial interest in or conflicts with the subject matter or materials discussed in the article.
Funding Information
This article did not receive any funding.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.