
Abstract
Nutritional manipulations can reduce the detrimental effects of heat stress on animal health and production. Akkermansia muciniphila (AM) is an innovative beneficial bacteria and can be used for conventional use as dietary supplements and pharmaceutical application. This study aimed to investigate the effects of administering AM on gut morphology, antioxidant indices, and gut microbiome of mice during heat stress. A total of 24 BALB/c mice were randomly assigned to three groups including the control group (CON), heat stress group (HS), and AM administration under heat stress group (AM). Our results showed heat stress significantly increased the water consumption of mice. Administration of AM did not improve feed intake or weight gain. The serum levels of alanine aminotransferase and aspartate aminotransferase as well as antioxidant parameters were not different among the three groups. Heat stress decreased the jejunal villus height, and AM could reverse this effect. AM administration significantly increased the relative abundance of Verrucomicrobiota at the phylum level. At the genus level, heat stress and AM groups tended to have a lower abundance of Alloprevotella. In addition, AM tended to increase the relative abundance of [Eubacterium]_xylanophilum_group in comparison with the other two groups. In summary, administration of AM can alleviate the damage of heat stress to the jejunum. However, it has no effect on serum antioxidant parameters, and its effect on the cecal microbiota is limited.
Introduction
Heat stress threatens the economic development of agriculture and animal husbandry. When the ambient temperature is higher than the animal’s body temperature and beyond its adjustable range, the heat generated and absorbed by the animal is greater than the heat dissipation. This disparity affects the heat metabolism balance of the animal and induces heat stress (Lees et al., 2019). Alkalosis is an abnormal pathophysiological condition characterized by the buildup of excess base or alkali in the body. Long-term high-frequency breathing of animals during heat stress decreases the carbon dioxide content in blood, causing alkalosis (Hales and Webster, 1967). In addition, heat stress causes damage to various organs and tissues, including liver, kidney, etc., and cause inflammatory reactions (Cui et al., 2019; Park et al., 2021; Sejian et al., 2017). Heat stress damages the intestinal mucosal barrier, leading to an increase in intestinal permeability and making it easier for pathogenic bacteria and harmful toxins to pass through the intestinal mucosa (Cui et al., 2019; Suzuki, 2020). Thus, high environmental temperature affects the growth of young mammals, reduces meat production, milk production, and meat quality, and even leads to animal death (Du D et al., 2022; Liu et al., 2020; Song et al., 2014). Production performance and egg quality of laying hens are reduced under heat stress (Abd et al., 2019; Tang et al., 2022b). Therefore, it is particularly important to seek methods, which can effectively alleviate the impacts of heat stress.
Akkermansia muciniphila (AM) is a type of Gram-negative anaerobic bacterium that can degrade mucus to produce short-chain fatty acids, providing energy for the host and promoting colonization of itself. AM showed competitive inhibition of pathogenic bacteria (Zhang et al., 2019). The abundance of AM is significantly reduced in animals under certain pathological conditions, such as obesity, diabetes, and inflammatory bowel disease (Depommier et al., 2019; Peeters et al., 2019; Yassour et al., 2016). A previous study found that AM can promote intestinal epithelial cell proliferation and alleviate intestinal damage of the chicken caused by Salmonella (Zhu et al., 2020). Another study showed that AM reduced systemic inflammatory factors (tumor necrosis factor-α [TNF-α], interleukin [IL]-1α, IL-6, and IL-12A), demonstrating anti-inflammatory effects in the intestines (Bian et al., 2019). Therefore, the objective of this study was to investigate the protective effects of AM on gut morphology, antioxidant indices, and gut microbiome of mice during heat stress.
Materials and Methods
Ethics statement
The animal study was reviewed and approved by the Institutional Animal Care and Use Committee of Foshan University (FOSU2023198, Foshan, China).
Preparation of AM bacterial solution
Standard strain AM (ATCC BAA-835) was purchased from the Guangdong Microbial Culture Collection Center. Using a sterile Pasteur pipette, 0.3–0.5 mL of suitable rehydration fluid was added into the ampoule. The sample was spread on a brain heart infusion (BHI) plate and incubated under the directed condition. After activation, it was cultured in improved BHI liquid medium (Guangdong Envitech Microbial Technology Co., Ltd.) with the addition of 2.5% mucin (Shanghai Yuan Ye Technology Co., Ltd.) under anaerobic conditions at 37°C.
Animal experiment design
Twenty four 8-week-old BALB/C male mice (obtained from Guangdong Provincial Medical Experimental Animal Center; Foshan City, Guangdong Province) were randomly divided into three groups including the control group (CON), heat stress group (HS), and AM administration under heat stress group (AM). Each treatment group had eight mice. During the adaptation period, all mice had free access to sterile water and SPF feed. The study lasted for 26 days, and the room temperature was kept constant at 24–25°C (humidity 50–60%). During the study, the control and HS group mice received 200 μL phosphate-buffered saline orally every morning at 9 a.m., whereas the AM group mice were orally gavaged 200 μL AM solution (8 × 108 CFU/day). The heat stress model was constructed by simulating a high temperature environment (40 ± 1°C, 50–60% humidity) using an incubator. From day 13 to 26, the HS and AM group mice received 2-h heat stress every day from 11:00 a.m. to 13:00 p.m. During the study, body weight, feed intake, and water consumption were recorded daily. On the last day, blood samples from both orbital sinuses of each mouse were collected. Then, all mice were euthanized by cervical dislocation. Samples of different intestinal segments, liver, and cecal digesta were collected.
Measurement of indicators
Feed intake, water consumption, organ index, and gut morphology
The body weight, food intake, and water consumption were recorded daily. The segments of duodenum, jejunum, and ileum were collected and fixed with 4% paraformaldehyde for 24 h. Then, the samples were dehydrated using ethanol, embedded in paraffin, cut into 0.4 μm slices, and stained using the hematoxylin–eosin method. The slices were observed under an optical microscope (40×); the villus height and crypt depth were measured, and their ratio was calculated.
Measurement of serum parameters
The serum levels of aspartate aminotransaminase (AST), alanine aminotransaminase (ALT), and antioxidant parameters including total antioxidant enzymes (T-AOC), catalase (CAT), malondialdehyde (MDA), superoxide dismutase (SOD), and glutathione peroxidase (GSH-Px) were measured according to the user instructions (Nanjing Jiancheng Institute of Bioengineering).
High-throughput sequencing analysis
Genomic DNA was extracted from cecal digesta using the Cetyltrimethylammonium bromide (CTAB) method (Trojanek et al., 2017). After determination of DNA quality and quantity, total digested DNA was diluted to prepare amplicons for high-throughput sequencing. Conventional polymerase chain reaction (PCR) was used to amplify the V3V4 region of the 16S rRNA genes using primers 515F (5′-GTGCCA GCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). The resulting DNA libraries were sequenced on the Illumina NovaSeq platform (Novogene, CNH) to generate reads, which were merged and then filtered, clustered into amplicon sequence variants (ASVs), and subjected to species annotation and abundance analysis to reveal the composition of the samples. Taxonomic analysis was performed at the phylum and genus levels. Alpha diversity analysis was conducted to study the complexity of species diversity using Quantitative Insight into Microbial Ecology (V1.9.1).
Statistical analysis
The experimental data were subjected to one-way analysis of variance (ANOVA) using SPSS Statistic 22.0 (SPSS, Inc.). Significant differences between means were compared using Duncan’s multiple comparisons. Significance was declared at p < 0.05 and trends at p < 0.1.
Results
Feed intake, water consumption, organ index, and gut morphology
According to Table 1, mice subjected to heat stress had significantly higher water consumption than the control group (p < 0.05; Table 1). The AM group mice had significantly lower feed intake than mice in the control group (p < 0.05). There was no significant difference in the percentages of liver and spleen (% of BW) among the three groups. The length of the cecum and colon were not different. According to Table 2, heat stress significantly decreased the villus height of the jejunum and ileum (p < 0.05; Table 2). Administration of AM increased the villus height of the jejunum, not the ileum, compared with the HS group. The AM group had a higher crypt depth of jejunum than the other two groups (p = 0.00).
Effects of AM on Feed Intake and Organ Indices of Mice During Heat Stress
Means within a row with different superscripts differ significantly (p < 0.05) is the footnotes for indicators “a and b”.
CON, control; HS, heat stress; AM, Akkermansia muciniphila; SEM, standard error of the mean.
Effects of AM on Intestinal Villus Height and Crypt Depth of Mice During Heat Stress
Means within a row with different superscripts differ significantly (p < 0.05) is the footnotes for indicators “a and b”.
CON, control; HS, heat stress; AM, Akkermansia muciniphila; SEM, standard error of the mean.
Serum ALT, AST, and antioxidant parameters
According to Table 3, the serum levels of ALT and AST were not different among the three groups. Heat stress treatment tended to decrease the serum levels of AST in comparison with the control group (p = 0.08; Table 3). There was no significant difference observed in levels of T-AOCs, CAT, GSH-Px, MDA, and SOD among the three groups.
Effects of AM on Serum ALT and AST, and Antioxidant Parameters of Mice During Heat Stress
Means within a row with different superscripts differ significantly (p < 0.05).
ALT, alanine aminotransferase; AST, aspartate aminotransferase; T-AOC, total antioxidant enzymes; CAT, catalase; MDA, malondialdehyde; SOD, superoxide dismutase; GSH-Px, glutathione peroxidase; CON, control; HS, heat stress; AM, Akkermansia muciniphila; SEM, standard error of the mean.
Species diversity and gut microbiome
A total of 655 ASVs were shared by all three groups, and the CON, HS, and AM groups have 182, 284, and 290 unique ASVs, respectively (Fig. 1). According to Table 4, the alpha diversity indices, including Chao1, Shannon, Simpson, and observed_OTUs, were not different among the three groups (p > 0.05). At the phylum level, Firmicutes and Bacteroidota were the dominant bacteria in all groups (Table 5). The AM group had a significantly higher relative abundance of Verrucomicrobiota than the CON and HS groups (p < 0.05; Fig. 2A). The rest of the top 10 phyla were not different among the three groups (p > 0.05). At the genus level, it can be seen that heat stress tended to decrease the abundance of Alloprevotella (p = 0.08; Table 6). Heat stress tended to increase the relative abundance of Colidextribacter. Oral gavage of AM tended to increase the abundance of Eubacterium_xylanophilum_group as its relative abundance was numerically higher than the other two groups (p < 0.05). AM was not detected in the CON and HS groups, and it was significantly higher in the AM group due to oral gavage (p = 0.03; Table 6 and Fig. 2B).

Venn diagram of shared ASVs among the three groups. ASV, amplicon sequence variant; CON, control; HS, heat stress; AM, Akkermansia muciniphila.
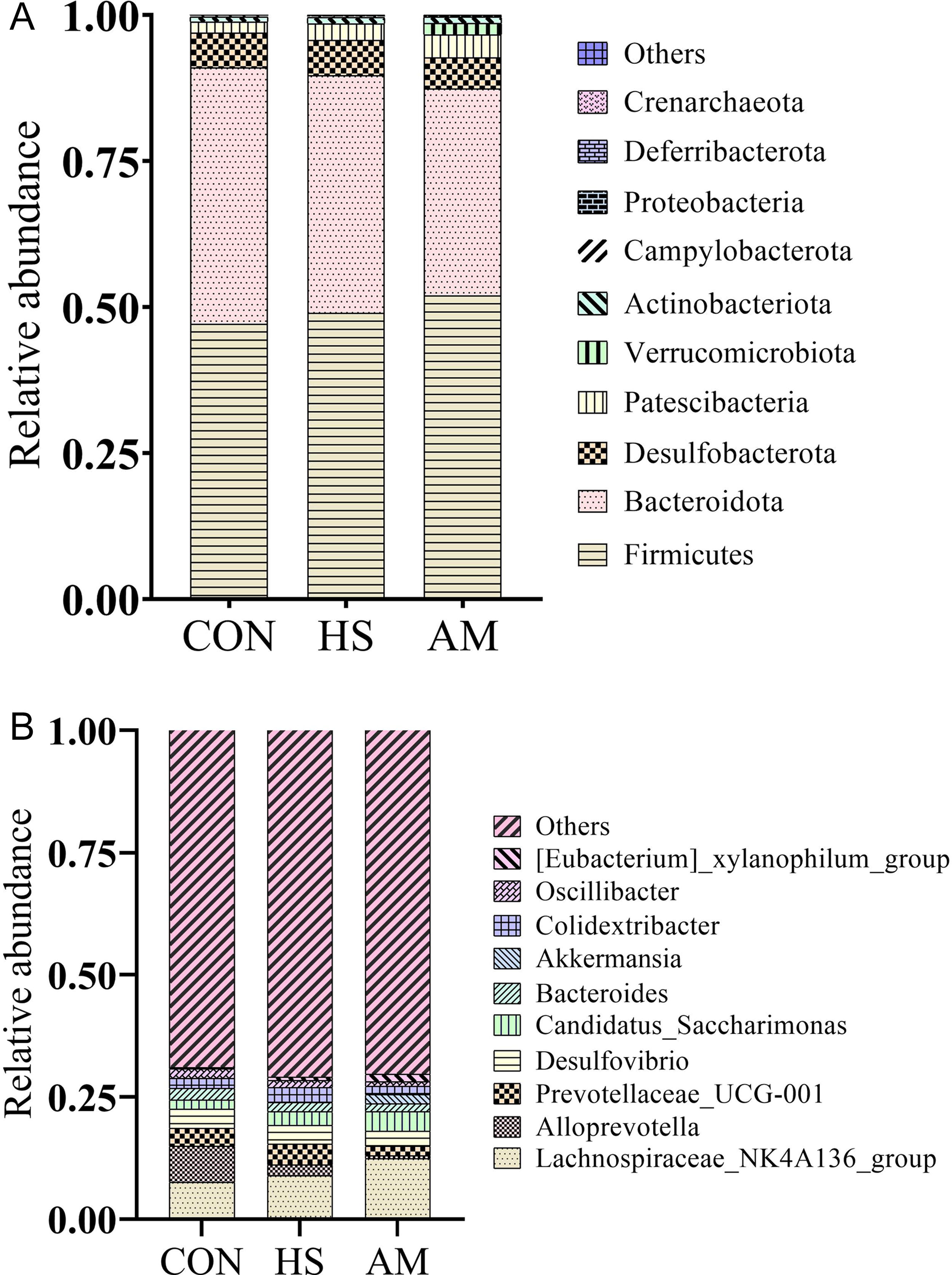
Phylum-level
Effects of AM on Alpha Diversity of Cecal Microbiota in Mice During Heat Stress
CON, control; HS, heat stress; AM, Akkermansia muciniphila; SEM, standard error of the mean.
Effects of AM on the Cecal Microbiota Composition at the Phylum Level in Mice During Heat Stress
Means within a row with different superscripts differ significantly (p < 0.05) is the footnotes for indicators “a and b”.
CON, control; HS, heat stress; AM, Akkermansia muciniphila; SEM, standard error of the mean.
Effects of AM on the Cecal Microbiota Composition at the Genus Level in Mice During Heat Stress
Means within a row with different superscripts differ significantly (p < 0.05) is the footnotes for indicators “a and b”.
CON, control; HS, heat stress; AM, Akkermansia muciniphila; SEM, standard error of the mean.
Discussion
High temperature has a significant negative impact on the productivity of livestock. It reduces animal feed intake, exacerbates intestinal inflammation, disrupts gut microbiota balance, and alters metabolite composition (Tang et al., 2021;Tang et al., 2022a; Zhang et al., 2022). The intestine plays an extremely important role in the growth and production processes of animals, as it functions in digestion and absorption. Heat stress has a significant adverse effect on the relative weight, villus height, and crypt depth of the duodenum, jejunum, and ileum of the animals (Goel, 2021). AM is considered a promising probiotic. It can degrade adhesive proteins, and the degradation products can modulate the host immune system through signals such as TNF-α, interferon-γ, IL-10, and IL-4 (Andersson et al., 2013). The decrease of AM is closely associated with the occurrence of inflammatory bowel disease, and the abundance of AM in the intestinal mucosa of patients with enteritis is significantly decreased (Rajilic-Stojanovic et al., 2013). AM may have the function of protecting the intestines and alleviating oxidative stress or intestinal inflammation caused by heat stress. Our study demonstrated that heat stress significantly increased water consumption but did not affect the feed intake and weight of the mice. Heat stress decreased villus height in the jejunum and ileum (Table 2). Administration of AM effectively alleviated the damage of heat stress to the jejunal villi, indicating that it could be used as effective supplements during heat stress.
Heat stress can also lead to degeneration in immune-related organs and tissues in animals, such as the spleen, thymus, and lymphoid organs (Wasti et al., 2020). Heat stress has been proven to cause oxidative damage in various animal species, such as broiler chickens, pigs, and ducks (Liu et al., 2016; Nanto-Hara et al., 2021; Yang et al., 2021). Studies have indicated that high temperatures can lead to oxidative damage in the livers of broiler chickens, resulting in disrupted lipid metabolism (Emami et al., 2020). Oxidative stress leads to an increase in the levels of oxygen-free radicals, whereas the activity of antioxidant enzymes and the ability to remove free radicals decrease (Miao et al., 2020). The liver, due to its metabolic and detoxification properties, is the main site of oxygen-free radical production. When the redox balance shifts toward the oxidative state, it is regarded as the initial step of liver damage (Sadasivam et al., 2022). In experiments on heat-stressed broiler chickens, it was found that heat stress significantly increased the levels of AST in the blood serum and significantly reduced the amount of SOD in the liver (Ding et al., 2023). A study on stress-induced rearing of Japanese quails has shown a significant increase in the level of MDA and a decrease in the activity of SOD, CAT, and GSH-Px (Sahin et al., 2012). In our experiment, compared with the CON group, there were no significant changes observed in the weight and organ index of the liver and spleen in the HS group (Table 1). The length of the cecum and colon also did not show significant changes. However, mice treated with AM via gavage showed a trend toward a shortened cecum and colon compared with the CON and the HS. Heat stress did not significantly affect the levels of AST and ALT and the antioxidant capacity of the mice, as there were no significant changes in T-AOC, CAT, GSH-Px, MDA, and SOD in the serum (Table 3). This may be due to the fact that 2 h of heat stress (40 ± 1°C) is not sufficient to cause enough damage to the liver, or it is possible that the mild damage caused by heat stress was recovered. For heat stress studies on mice, temperature of 39–43°C was normally used (Chen et al., 2022; Lee et al., 2015; Wen et al., 2021). However, the heat exposure time was different. For high temperature such as 43°C, the time of heat exposure daily was short such as 15 min (Lee et al., 2015). In our study, we were interested in the impact of chronic heat stress, so we chose 40°C and 2 weeks of heat exposure in the current study. Further research is needed to investigate the potential of gastric administration of AM for alleviating liver damage.
The relative equilibrium between microorganisms and intestinal mucosa is of great significance for the integrity of the intestinal barrier and host health in animals. However, under high temperature conditions, endoplasmic reticulum stress-induced apoptosis occurs in animal cells, especially intestinal cells, which disrupts the intestinal barrier and increases intestinal permeability (Koch et al., 2019; Yi et al., 2021). The occurrence of general intestinal inflammation is due to increased permeability of intestinal epithelial cells, which allows pathogenic bacteria, their metabolites, and some harmful substances to enter the bloodstream more easily (Koch et al., 2019). There are studies showing that the composition of the gut microbiota is influenced by heat stress. For example, in high temperature environments, the proportion of Proteobacteria, Bacillales, Planococcaceae, Kurthia, and Streptococcaceae increases, whereas the proportion of Bacteroidetes and Bacteroidia decreases. Furthermore, heat stress also affects the abundance of Firmicutes, Proteobacteria, and Spirochaetes (Le Sciellour et al., 2019; Xiong et al., 2022). In experiments on broiler chickens, it was found that the proportion of Firmicutes and Bacteroidota changed significantly under heat stress conditions (Yin et al., 2021). Heat stress can downregulate and alter the microbial composition related to bile acid metabolism, such as Bacteroides, Clostridium, and Lactobacillus (Cai et al., 2022; Jia et al., 2018; Sato et al., 2021). In addition, it is very likely that the increased intestinal permeability is achieved by an increase in the proportion of Proteobacteria and a decrease in the proportion of Bacteroidetes (He et al., 2019; Koch et al., 2019). Research has found that heat stress increases the Chao 1 and observed_species indices in the ceca of broiler chickens (Wang et al., 2018). In this experiment, alpha diversity analysis showed no significant changes among the three groups (Table 4). The abundance of Verrucomicrobiota in the cecal contents of AM-treated mice significantly increased (Table 5). This increase could be due to the fact that AM is a representative species in the phylum Verrucomicrobiota (Upreti et al., 2022). At the genus level, the abundance of Alloprevotella and Colidextribacter in AM group mice showed a decreasing trend (Table 6). Alloprevotella is commonly recognized as a probiotic with anti-inflammatory effects (Xie et al., 2022), and it showed a decreasing trend in the HS group in this experiment. However, the relationship between AM and Alloprevotella, as well as the mechanisms of their mutual influence, is still unclear and requires further investigation. There is a study showing that the content of Colidextribacter in mice with colitis increased fivefold (Xie et al., 2022). In this experiment, there was an increasing trend of Colidextribacter abundance in the cecum microbiota of the HS group mice, whereas the AM group showed a decrease in Colidextribacter abundance (Table 6). The [Eubacterium]_xylanophilum_group is generally considered a harmful bacterium (Li et al., 2022), which was increased in both the HS group and the AM group in this experiment, with a higher increase observed in the AM group. Further research is needed to understand the correlation between AM and other microbial communities, as well as the mechanism of regulating gut microbiota.
Conclusion
Oral gavage of AM effectively alleviated the damage of heat stress to the jejunal villi, but it did not improve feed intake or weight gain. It had no effects on serum antioxidant parameters, and its effect on cecal microbiota was limited.
Footnotes
Authors’ Contributions
C.L. and Y.D. conducted the study and collected samples as well as drafted the article. C.L., R.Z., and Q.Q. analyzed the samples. S.L. performed the data analysis. X.F. reviewed and revised the article. All authors read and approved the final article.
Disclosure Statement
No potential conflict of interest was reported by the authors.
Data Availability Statement
Funding Information
This work received financial support from the Discipline Construction Program of Foshan University (CGZ0400162), the Research Start-Up Fund for Postdoctoral Fellows from Foshan City (BKS209059), and the Scientific Research Start-Up Fund for High-Level Talents of Foshan University (Gg07145).