
Abstract
This retrospective investigation (2019–2022) identified two plasmid-mediated mcr-10 from 6800 food samples in Shanghai, China and localized in a conjugative plasmid (pEC1918-mcr10) in Escherichia kobei from ready-to-eat food with high-level polymyxin B resistance, and a nonconjugative plasmid (pEC2001–mcr10) in E. coli from chicken. These genes were adjacent to ISEc36. This report highlights the emergence of mcr-10 from food samples in Shanghai, China. Active surveillance of vital resistance genes along food production chain should be performed.
Although polymyxin is an old generation antibiotic, it is still used as the last line defense against fatal infections caused by multidrug resistant Gram-positive bacteria (Paterson and Harris, 2016). Consequently, polymyxin resistance has gradually increased in the past few years. While bacteria mostly acquire polymyxin resistance via chromosome point mutations (Carroll et al., 2019), and recent studies have reported that such resistance can also occur because of plasmid-borne mcr genes (Bortolaia et al., 2020). The first plasmid-encoded mcr-1 from Escherichia coli and Klebsiella pneumoniae isolated from patients, food, and animals in 2015 has been described (Liu, et al., 2016). Until now, nine other mcr variants (from mcr-2 to mcr-10) have been identified (Liu et al., 2022). In 2020, Wang et al. first reported the novel mcr gene in the IncFIA plasmid, mcr-10, from a clinical Enterobacter roggenkampii isolate (Wang et al., 2020). Subsequently, mcr-10 was identified in several Enterobacteriaceae species found in animals, the environment, and humans in many countries. E. roggenkampii isolates harboring mcr-10 were found from anal swabs from chickens and hospital sewage water in China (Lei et al., 2020; Xu et al., 2021), Klebsiella quasipneumoniae with mcr-10 was first identified in human wastewater samples in Italy (Formenti et al., 2022). The presence of mcr-10 in Enterobacter kobei from neonatal blood cultures in Nepal (Manandhar et al., 2022), as well as in Cronobacter sakazakii, obtained from the fecal sample of a healthy volunteer in China (Yang et al., 2021). Notably, these mcr genes can transmit between bacterial isolates and species, even among various genera found in animals, humans, and the environment by horizontal transmission (Liu et al., 2022). These strains harboring mcr-10 pose significant public health threats and can cause treatment failure in humans. Here, we report two plasmid-mediated mcr-10 in Escherichia kobei and E. coli from ready-to-eat food and chicken, respectively, in China.
Results
Presence of mcr-10 in E. kobei and E. coli
Herein, we screened 6800 food samples, including raw meat from poultry and others, salads, and cooked meat, to detect pathogenic bacteria with mcr-10 gene between 2019 and 2022 in Shanghai, China. The mcr-10 gene was identified using real time PCR to detect mcr-10 using the primers mcr-10-F(GATATCCTGAGCCGTCTTGAAC), mcr-10-R (CGAACCGAGAAGGGCATAAA) and mcr-10-P (FAM-ACAAACGGAATGGAGATGACGAACCC-BHQ1). Only two mcr-10 genes were found (0.029%, 2/6800). Using MOLDI-TOF, we identified these two genes in E. kobei (EK1918) in ready-to-eat food (RTE food) and E. coli (EC2001) from chicken, respectively. The antibiotic susceptibility testing revealed that EC1918 was polymyxin B (PB) resistant [minimum inhibitory concentration (MIC) ≥8 μg/mL], whereas EC2001 was susceptible to PB (MIC = 0.25 μg/mL) (Table 1). These two strains harboring mcr-10 were susceptible to other antibiotics, including ampicillin-sulbactam, compound sulfamethoxazole, cefazolin, cefotaxime, ceftazidime, cefoxitin, gentamicin, imipenem, nalidixic acid, azithromycin, sulfisoxazole, and ciprofloxacin.
Characteristics of Mcr-10-Positive E. Kobei, and Its Transmission in E. coli, and Their Sources, Antimicrobial Resistance Profiles and Resistance Determinants Along with MIC for Polymyxin
MIC, minimum inhibitory concentration; PB, polymyxin B.
Transferable and nontransferable mcr-10
We conducted conjugation experiments using ampicillin-resistant E. coli JM109 as the recipient, which had the plasmid pUC18 with Amp gene. The transconjugants were selected on Mueller–Hinton agar plate containing ampicillin (100 mg/mL), and confirmed using real time PCR to detect mcr-10. The results confirmed that pEK1918-mcr10 could be successfully transferred to JM109 to form a transconjugant termed as JM1918-mcr10 with transfer efficiency values of ∼10−7, which decreased susceptibility to polymyxin (MIC = 0.5 mg/mL) (Table 1). However, pEC2001-mcr10 could not be transferred to JM109.
We obtained the genome and complete plasmid sequence of EK1918 using PromethION nanopore sequencing platform (Oxford, United Kingdom). A total of 1280 Mb clean data with an N50 of 30,981 bp was generated. The complete genome was 5.03 Mb with a 54.79% GC content. The ribosomal multilocus sequence typing (rMLST), an approach indexing variation of the 53 genes encoding the bacterial ribosome protein submits (rps genes), was used to identify EK1918. And rMLST of EK1918 showed 100% similarity with that of E. kobei (https://pubmlst.org/bigsdb?db=pubmlst_rmlst_seqdef_kiosk). EK1918 harbored the genes blaACT-52 and mcr-10 that confer resistance against β-lactam antibiotics and polymyxin, respectively, identified using the Comprehensive Antibiotic Resistance Database (https://card.mcmaster.ca/analyze/rgi). EK1918 harbored one mcr-10-bearing plasmid, which has 92% nucleotide identity with that in pEC81-mcr10 and pl9455333 with mcr-10 (Fig. 1A) (GenBank accession numbers: NZ_CP088133.1 and CP122443.1 respectively).
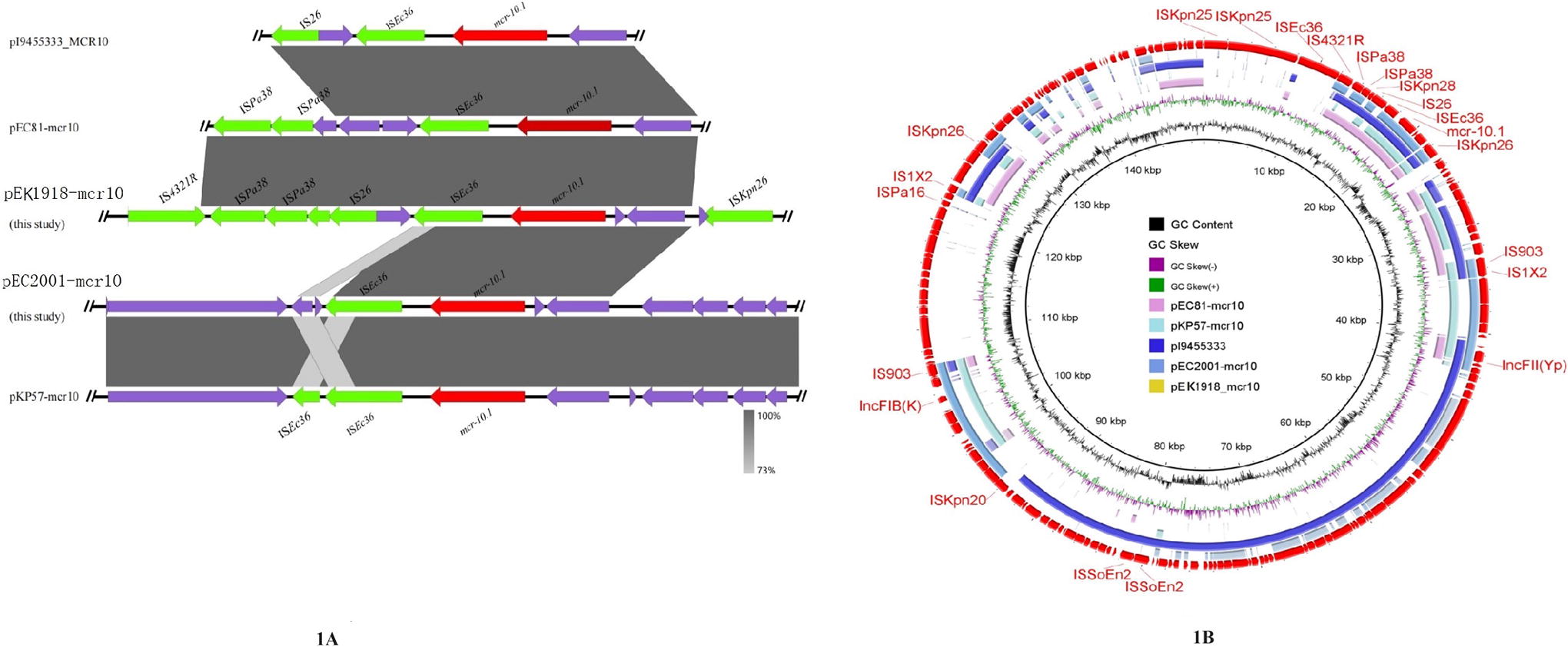
Genetic environment of mcr-10 in the pEK1918-mcr10 and pEC2001-mcr10.
The genome and complete plasmid sequence of EC2001 were sequenced using PromethION nanopore sequencing platform (Oxford, United Kingdom), which generated 1285 Mb clean data with an N50 of 32,636 bp. The draft genome was 5.56 Mb with a 50.99% GC content. The rMLST of EC2001 demonstrated 100% with that of E. coli. Furthermore, EC2001 contained the virulence gene astA, which identified EC2001 as Enteroaggregative E. coli (EAEC). In addition, EC2001 harbored two plasmids, and one harbored mcr-10, which exhibited 100% nucleotide identity with that in pKP57-mcr10 from Klebsiella pneumoniae (Fig. 1A) (GenBank accession number: NZ_CP088129.1).
The complete sequence of the mcr-10-harboring plasmid in EK1918, termed as pEK1918-mcr10, was 149,596 bp in size with 52.39% of GC content. Using Plasmid-Finder 2.1, we predicted that the genes encoding the replication initiation proteins IncFII(Yp) and IncFIB(K) (http://www.genomicepidemiology.org/services/). The complete sequence of the mcr-10-harboring plasmid in EC2001, termed as pEC2001-mcr10, was 139588 bp, has 53.09% of GC content. Plasmid-Finder 2.1 predicted that the genes encoding the replication initiator proteins IncFII(Yp), IncFIB(K).
Genetic characterization of plasmids harboring the mcr-10 gene
On analyzing the genetic context of the mcr-10-containing plasmids, we found that mcr-10 in all five plasmids (pEC1918-mcr10, pEC2001-mcr10, pI9455333_MCR10, pEC81_mcr10, and pKP57_mcr10) were adjacent to ISEc36 (Fig. 1B). The genetic context of pEK1981-mcr10 and pEC2001-mcr10 were IS4321R-ISPa38-ISKpn28-IS26-ISEc36-mcr-10-ISKpn26 and IS4321R-ISEc36-mcr-10-ISKpn26, respectively. Additionally, pEC1918-mcr10 harbored the multiple inserted sequences, including ISSba14, IS102, ISSoEn2, ISKpn28, ISKpn20, IS903, IS1X2, IS5075, and IS1618, and pEC2001-mcr10 harbored the inserted sequences including IS1A, IS903B, IS1222, IS5708, ISEc10, and IS1X3 (Fig. 1B). Reportedly, mcr-10-containing plasmids are related to IncFII and IncFIB (Lei et al., 2020; Zhang et al., 2022). In addition to these two replicons, the replicon IncR was detected in pEC2001-mcr10, further expanding the repertoire of mcr-10-associated plasmids.
Discussion
The spread of polymyxin-resistant bacteria is due to the transmission of plasmid-mediated mcr genes via the horizontal transfer of plasmids. To date, 105 slightly different variants of the mcr genes have been identified in bacteria (https://card.mcmaster.ca/ontology/36006). Since its initial discovery in a human clinical sample in 2020 (Wang et al. 2020), mcr-10 has been found in animals, environment, and human (Hussein et al., 2021; Lei et al., 2020; Xu et al., 2021). Here, we detected two mcr-10 genes in different food samples, although the detection rate was low (0.029%, 2/6800). These genes were found in EAEC and E. kobei isolated from chicken and RTE food, respectively. To our knowledge, this is the first report showing the presence of mcr-10 showing high-level polymyxin B resistance from RTE food. In 2022, E. kobei strains with mcr-10 caused and outbreak in a tertiary care hospital in Nepal (Manandhar et al., 2022). Therefore, it is important to monitor foodborne diseases caused by E. kobei with the mcr-10. The RTE food contained beef salad made of medium beef and vegetables, cooked at 63°C, which might not be sufficient to kill the E. kobei with mcr-10. Therefore, further microbiological surveillance of beef and cross-contamination among different foods will be necessary for food safety. Moreover, as the EAEC strain with astA is pathogenic to humans, continuous surveillance of this strain in food especially meats is important for food safety.
It is worth noting that E. kobei EK1918 isolate carrying the mcr-10 showing high-level polymyxin B resistance in this study. However, it has been reported that the MIC of the mcr-10-carrying isolates ranged from 0.5 μg/mL to 1 μg/mL (polymyxin susceptible) (Liu et al., 2022). Furthermore, the transconjugant EK1918-mcr10 showed susceptibility to polymyxin (MIC = 0.25 mg/mL). Unknown resistant mechanism of the mcr-10 might exist in E. kobei in this study. For example, efflux pumps and other genetic mutant might play a key role in polymyxin resistance (Webber and Piddock, 2003). Therefore, further study is necessary to make clear the resistance mechanism of high-level polymyxin of E. kobei with the mcr-10, which help broaden the knowledge of mechanism of the mcr-10.
These two mcr-10 genes originated from chicken and beef, and differed from those present in bacterial strains, although both the strains belonged to Enterobacteriaceae. Taken together, the mcr-10 genes could be transmitted among bacterial belonging to Enterobacteriaceae via clone transmission. Newer mcr-10 gene variants can emerge owing to the rapid transmission of mcr-10 genes. Therefore, understanding the transmission mode of mcr-10 genes is crucial for actively surveilling plasmid-mediated mcr-10 genes to ensure food safety.
Our findings indicate that E. kobei from RTE food harboring the mcr-10 in the transferrable plasmid pEK1918-mcr10 exhibits high-level polymyxin B resistance. The polymyxin-susceptible EAEC harbored mcr-10 in non-conjugative plasmid pEC2001-mcr10 in chicken. These results suggest that mcr-10 might transmit among food-producing animals and their products via horizontal gene transfer. Therefore, the continuous monitoring of these resistance genes in food is essential to understand and take measures against the transmission of antibiotic resistance genes from foods to humans and health care facilities and from food-producing animals to food.
Footnotes
Authors’ Contributions
H.Z. and W.J. performed the research and draft the article. X.Y., Y.Y., Q.X., and T.Q. performed the tests. H.U. and R.C. designed the study, supervised the project, analyzed data. All authors reviewed, revised, and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Three-Year Initiative Plan for Strengthening Public Health System Construction in Shanghai (2023–2025) Key Discipline Project, No. GWVI-11.1–09