
Abstract
The present study assesses the antibacterial effect of a high-voltage prick electric field (HVPEF) on Salmonella enteritidis on gelatin plates and selected food systems. The results showed that S. enteritidis were susceptible to HVPEF treatment, with a mortality rate of 98% on the gelatin plate at 13 kV for 15 min. The survival rate of S. enteritidis decreased linearly with time and then stabilized after a sharp decline. The morphological change and the loss of internal nucleic acids and proteins suggest that the cell membrane was damaged by HVPEF, appearing more permeable and less fluid. Compared with the untreated group, both reactive oxygen species (ROS) and malondialdehyde increased significantly in the treated groups. Flow cytometry results suggest that DNA was severely damaged, which led to cell death, as shown by the elevated levels of intracellular ROS and lipid peroxidation. The death rate of S. enteritidis on the food surface significantly rose with the increase of HVPEF intensity, reaching 91 ± 1.6% (salmon), 88.2 ± 2.2% (chicken breast), 72.9 ± 2.7% (sausage), and 66.9 ± 1.6% (steak), respectively, at 14 kV for 30 min. HVPEF resulted in no significant change in color or texture; the changes in total viable count and pH were less significant than in the untreated group. Finally, HVPEF treatment led to lower levels of total volatile base nitrogen and thiobarbituric acid reactive substances in chicken breasts, which potentially contributes to longer shelf life.
Introduction
Salmonella, a foodborne zoonotic pathogen (Mellor et al., 2019), is found in chicken, turkey, beef, pork, eggs, fruits, sprouts, and even processed foods (Ferrari et al., 2019; Han and Micallef, 2014; Hara-Kudo and Takatori, 2009). Salmonella infections are transmitted through food and cause salmonellosis in humans (McMillan et al., 2019). Salmonella enteritidis develops gastroenteritis, characterized by headache, fever, abdominal pain, diarrhea, nausea, and vomiting (Byun et al., 2021). Its prevention is of great significance in food safety.
Electric field sterilization (EFs) contributes to the protection of nutrition, flavor, color, and environment (El Kantar et al., 2018; Lopez-Alfaro et al., 2013; van Wyk et al., 2019). Pulse electric field (PEF), as a commonly utilized EFs technology, exhibits a strong bactericidal effect (Li et al., 2021) and is widely applied in treating food products like meat, fish (Gómez et al., 2019), and milk (Alirezalu et al., 2020). Currently, it is widely accepted that the sterilization mechanism of PEF primarily involves electroporation or electroosmosis on the cell membrane (Castro et al., 1993; Vega-Mercado et al., 1996; Zimmermann, 1986). High-voltage electrostatic field (HVEF) is another common mode of EFs (Dalvi-Isfahan et al., 2023). Similar to PEF, HVEF can cause damage to the cell membrane through electroporation (Tsong, 1991). The difference is that HVEF has a good bioelectric effect, which causes the change in the cell pH and the displacement of ions. HVEF can ionize the medium to produce ozone-free radicals and other substances to affect the integrity of the cell (Albertos et al., 2017; Wang et al., 2018).
Depending on the purpose, the electrodes of HVEF are configured as parallel-plate, pin-to-plate, or wire-plate forms (Dalvi-Isfahan et al., 2023). The electrode of a high-voltage prick electric field (HVPEF) is a pin-to-plate electrode, which produces corona discharges and forms an inhomogeneous electric field (Wang et al., 2018). Corona discharge produces ozone, which can sterilize bacteria (Chen and Davidson, 2003). Our previous research has found HVPEF possesses a strong antibacterial effect against Staphylococcus aureus (Qi et al., 2021).
This study aims to explore the antibacterial effects of HVPEF on S. enteritidis by determining the leakage of protein and nucleic acid, changes in cell morphology, membrane permeability and fluidity, damage to DNA, and reactive oxygen species (ROS) and malondialdehyde (MDA) levels, as well as the effect of HVPEF on the quality of chicken breasts.
Materials and Methods
Materials
S. enteritidis (ATCC 13076) was purchased from the China Institute of Food and Drug Testing (Beijing, China) and stored at −80°C. Cells were activated on TSA (Hope Bio-Technology Co., Ltd., Hangzhou, Zhejiang, China) plates and then cultured in 20 mL TSB (Hope Bio-Technology Co., Ltd.) at 200 rpm, 37°C for 12 h, followed by incubation until the stationary phase at 4.5∼5 × 109 CFU/mL. Chicken breast, salmon, steak, and sausage slices were from the Changchun Ouya Supermarket. All foodstuffs were prepared into uniform shapes (3 cm × 4 cm × 0.5 cm). HVPEF equipment was provided by Jilin University.
Antibacterial activity assay
S. enteritidis was diluted with 0.9% or 3% NaCl to obtain the concentration of 106 CFU/mL, and 100 μL was inoculated on TSB medium plates (5% gelatin) at 106 CFU/cm2. All plates received 15 min of HVPEF at 0, 1, 3, 5, 7, 9, 11, 12, 13, and 14 kV, respectively, to determine the antibacterial ability.
Gelatin plates were placed in the 37°C water bath to melt and then were diluted and coated for counting. The following calculation was used to compute the death rate:
Antibacterial mechanism assays
Protein and nucleic acid leakage
The release of protein and nucleic acid was determined following Ananda’s method (Ananda et al., 2019). Briefly, plates of 107 CFU/cm2 were treated with HVPEF of 13 kV at intervals of 0, 3, 6, 9, 12, and 15 min. After melting in the 37°C water bath, the bacterial suspensions were immediately 10-fold diluted and centrifuged at 4000 ×g for 5 min. OD values of the supernatant were measured at 260 nm and 280 nm by UV spectrophotometer (Bio-Rad Laboratories, Hercules, CA, USA).
Permeability and fluidity of membrane
S. enteritidis was treated as described in the “Protein and nucleic acid leakage” section to obtain the bacterial suspension. According to previous studies (Helander and Mattila-Sandholm, 2000), the suspension containing 10 μM n-phenyl-1-naphthylamine (NPN, Aladdin Biochemical Technology Co., Ltd., Shanghai, China) was incubated at 37°C for 10 min in the dark. The values were explored at excitation and emission wavelengths of 350 and 401 nm by a SPARK 20M multipurpose microplate reader (Tecan Austria GmbH, Austria). The relative fluorescence intensity was calculated as follows:
S. enteritidis suspension was resuspended in phosphate-buffered saline (PBS) to 106 CFU/mL. After introducing a 4 μM ANS fluorescent probe (Aladdin Biochemical Technology Co., Ltd., Shanghai, China), the mixture was incubated in the dark for 30 min (Domadia et al., 2010). The fluorescence of ANS at particular wavelengths (excitation: 385 nm; emission: 473 nm) was measured by a fluorescence spectrophotometer (Shimadzu RF-5301, Japan). The fluorescence value of the suspension was obtained by subtracting the average value of the stained PBS.
Bacterial morphology
S. enteritidis coated on solid media was collected after HVPEF treatment for 6 min and 15 min, respectively. According to Wang et al.’s approach (Wang et al., 2016), the morphological alterations of S. enteritidis were measured by field emission scanning electron microscopy (FESEM, S-3400 N, Hitachi, Japan).
ROS level and MDA content
S. enteritidis was treated as described in the “Protein and nucleic acid leakage” section, and the ROS test followed Su et al.’s with a few modifications (Su et al., 2019). The mixture with DCFH-DA fluorescent probe (Sigma-Aldrich, Saint Louis, Missouri, America) at 5 μM was cultured at 37°C for 30 min in the dark. Fluorescence was measured by a fluorescence spectrophotometer (Shimadzu RF-5301, Japan) at excitation and emission wavelengths of 485 nm and 520 nm. Values of stained bacteria were obtained by deducting the fluorescence mean of stained PBS.
Electric field treatment is as shown in the “Protein and nucleic acid leakage” section. MDA levels in cells were determined as follows: S. enteritidis was rinsed twice with PBS, followed by measurement with the protein detection kit (BCA, Nanjing Jiancheng Technology Co., Ltd., Nanjing, Jiangsu, China) and the oxidative damage index kit (MDA, Nanjing Jiancheng Technology Co., Ltd.).
Detection of DNA damage
Electric field treatment and heat melting dilution were performed as described in the “Protein and nucleic acid leakage” section. The bacterial solution was placed in the 37°C water bath with 5 mg/mL Rnase A for 30 min. According to the procedures of Tao et al. (2015), 0.5 mL was stained with 20 μL (0.1 mg/mL) Acridine Orange (AO, Solarbio Science & Technology Co., Ltd., Beijing, China) in the dark at 37°C for 15 min. Samples were examined using a flow cytometer (BD FACSCalibur, USA) after being cleaned with PBS. Bivariate dot plots were created with BD CellQuest Pro software.
Antibacterial test on food surfaces
S. enteritidis was diluted and then evenly inoculated to fresh chicken breast, fresh salmon, fresh steak, and sausage slices to obtain 106 CFU/cm2 and then treated with 13 and 14 kV for 0, 10, 20, and 30 min, respectively. Food samples were mixed with 90 mL of sterile distilled water and filtered with sterile gauze to obtain the sample filtrate. The filtrates were diluted and coated for counting. The antibacterial rate was calculated as in Eq. (1).
Application of HVPEF on chicken breast model
Total viable count and pH analysis
Fresh chicken breast samples were exposed to HVPEF for 30 min at 14 kV and then stored in sealed packages at 4°C for 0, 2, 4, 6, 8, and 10 days, respectively. The stored samples were mixed with 90 mL of sterile distilled water in a meat grinder and then filtered. The filtrate was diluted and coated for counting. The pH was detected using a pH meter (PHS-3C, Rex Electric Chemical, China).
Color and texture analysis
Chicken breast was treated as described in the “Total viable count and pH analysis” section. L*, a*, and b* values were measured using a colorimeter (CR-400, Konica Minolta, China). The hardness, elasticity, viscosity, and chewiness in the control and treatment groups were measured using a textural analyzer (CT3, Shanghai iBetter Technology Co., Ltd.). The measurement parameters include TPA mode, TA/5 flat-bottomed column probe, 5 g trigger force, 2.0 mm/s premeasurement speed, 1.0 mm/s mid-measurement speed, 5.0 mm/s postmeasurement speed, 50% deformation, and 5 s compression interval.
Determination of total volatile base nitrogen
Chicken breast was treated as described in the “Total viable count and pH analysis” section. The samples (5 g) were mixed with 25 mL of deionized water in a meat grinder, followed by stirring and shaking for 30 min and then filtration. Boric acid solution (2 mL) and methyl red methylene blue solution were added to the inner chamber of the diffusion dish; the sample filtrate and KOH solution were quickly added to the outer chamber; the diffusion dish was sealed, gently rotated to mix thoroughly, and incubated at 37°C for 2 h. After cooling, HCl (0.01M) was used to titrate until the solution turned blue-purple. The following calculation was used to compute the total volatile base nitrogen (TVB-N) value:
Determination of thiobarbituric acid reactive substances
Chicken breasts were treated and stored as mentioned in the “Total viable count and pH analysis” section. Mixed with 25 mL of 7.5% C2HCl3O2 (containing 0.1% EDTA), the chicken breast was shaken for 30 min and filtered, and the resulting filtrate was mixed with 5 mL of 0.02 mol/L TBA and heated in a 90°C water bath for 40 min. At room temperature, the supernatant was obtained by centrifugation at 2000 ×g for 5 min. Five milliliters of CHCl3 was added, shaken, and left to stratify, and the absorbance of the upper liquid was measured at 532 nm and 600 nm. The thiobarbituric acid reactive substances (TBARS) value was calculated as follows:
Statistical analysis
Each treatment was conducted in triplicate. The experimental data were expressed as mean ± standard deviation. The paired samples t-test was performed using SPSS 21.0 software (SPSS Inc., Chicago, IL, USA), with p < 0.05 indicating a statistically significant difference compared with the control group.
Results and Discussion
Antibacterial activity of S. enteritidis treated by HVPEF
The inactivation rate of S. enteritidis in fluid and solid media is shown in Figure 1. The highest antibacterial rates by HVPEF (14 kV for 15 min) in fluid media were up to 56 ± 2.3% and 68.7 ± 1.8% at 0.9% and 3% NaCl, respectively. The inactivation effect of 3% NaCl was better than 0.9%, and there was no significant difference. The reason for this is that 3% NaCl has better conductivity, which affected the electrical resistance of the electric field treatment chamber, further affecting the sterilizing effect (Wei & Li, 2014).
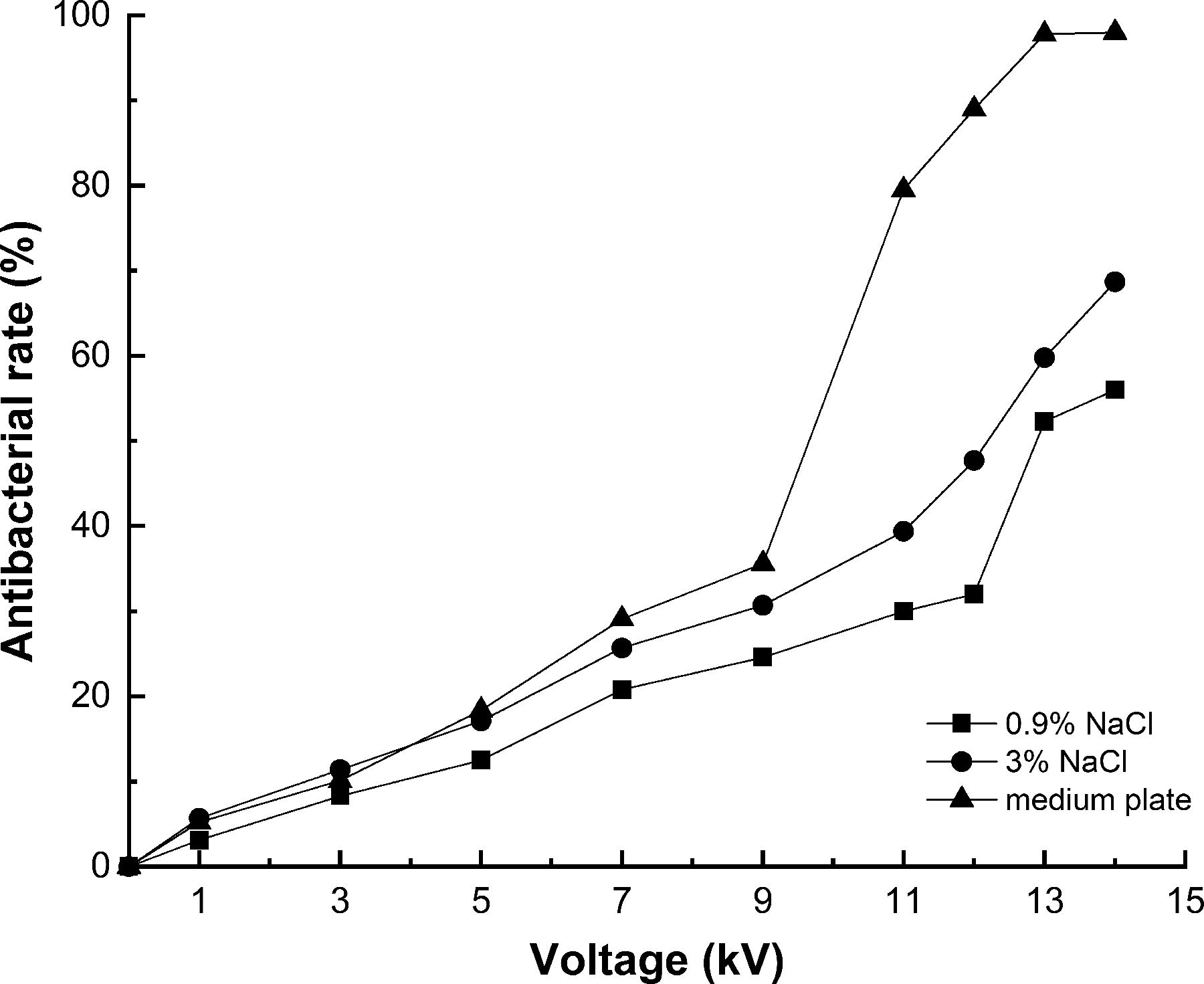
Mortality of Salmonella enteritidis exposed to high-voltage prick electric field with different voltages for 15 min.
As shown in Figure 1, with the voltage rising from 0 kV to 9 kV, the antibacterial effect improved slightly. When the voltage was further enhanced, the bactericidal rates obviously increased in varying degrees, in particular, on the plate, by 97.8 ± 0.6% at 13 kV and 98% at 14 kV, and then the curve flattened out. The sharp increase was likely due to the leakage of cell contents through the pores created by the Maxwell stress, resulting in irreparable cell damage (Sato et al., 2019).
The antibacterial mechanism
Effect of HVPEF treatment on S. enteritidis membrane
As shown in Figure 2a, the nucleic acid content in the treated cells showed a significant difference at 13 kV for 3 min (p < 0.01). The trend was more significant with the increase of time, especially after 9 min. The same trend occurred with the protein content. The aforementioned findings demonstrated that HVPEF treatment damaged the integrity of the cell membrane and caused intracellular substance leaking. Huang et al. explored the effects of HVEF on A. radioresistens and found that it caused abnormal cell morphology, increased membrane permeability, and irreversible membrane integrity damage, which led to the leakage of protein, nucleic acid, and other substances (Huang et al., 2021).
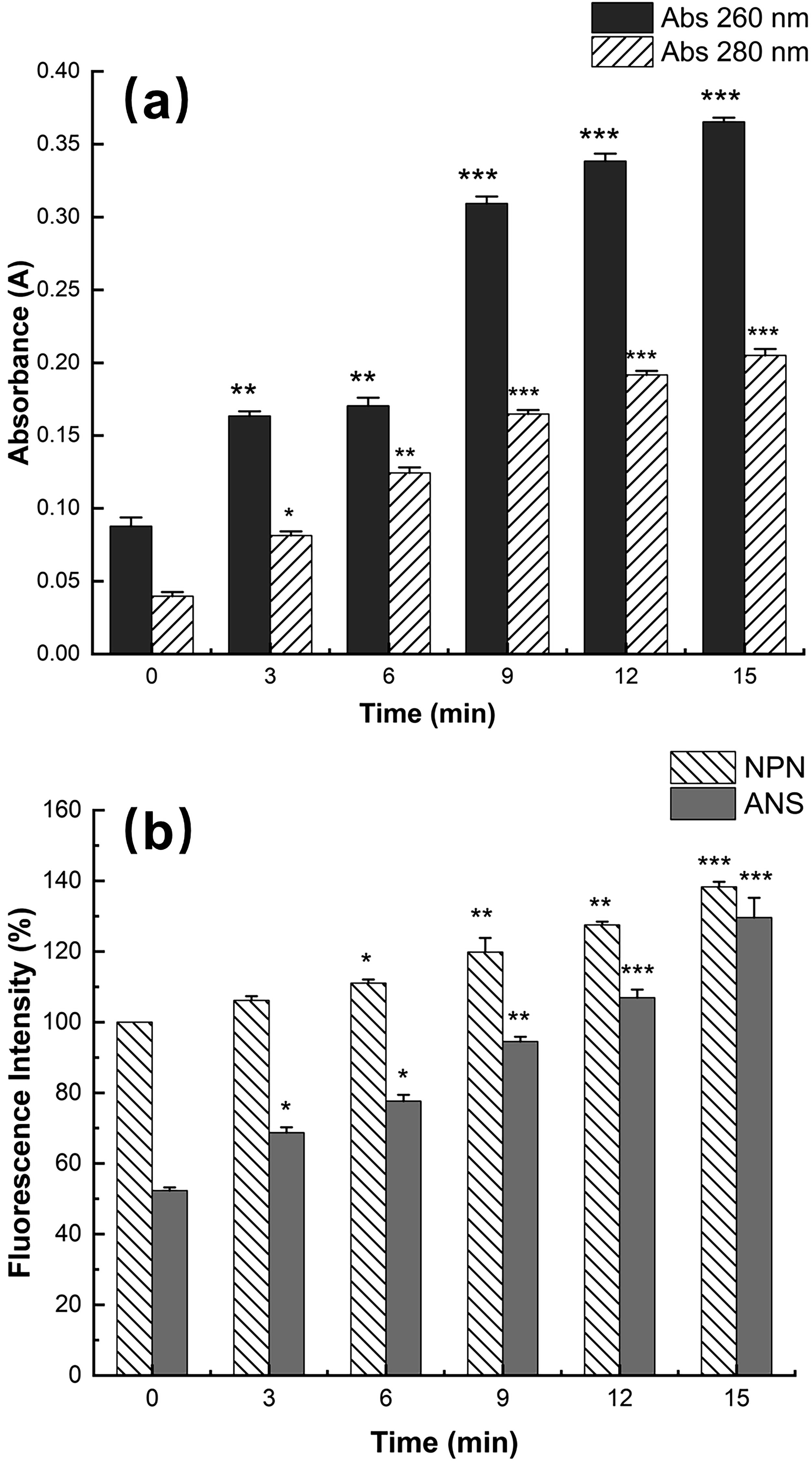
The effect of high-voltage prick electric field treatment on the leakage of intracellular substances
To further assess the integrity of the membrane, we investigated the changes in membrane fluidity and permeability. As shown in Figure 2b, the fluorescence intensity significantly increased after 6 min at 13 kV (p < 0.05) compared with untreated cells. With the extension of time, the fluorescence intensity continued to increase and reached the maximum at 15 min. According to these findings, HVPEF treatment increased the permeability of bacterial OM, which discharged intracellular substances and ultimately resulted in cell death (Garcia et al., 2007). The study shows that a greater lateral stacking degree of liposome C-H leads to a lower fluidity of cell membrane and a smaller liposome permeability (Liu et al., 2016), which explains that S. enteritidis resisted HVPEF by changing membrane permeability and reducing membrane fluidity in the present study.
To sum up, HVPEF treatment led to the leakage of proteins, nucleic acids, etc., and ultimately cell death. The results were further validated using FESEM.
FESEM analysis
Untreated cells exhibited regular shapes, smooth surface, and intact membrane (Fig. 3a). After 6 min of HVPEF treatment, the cell was flattened, with holes, wrinkles, and collapses on the membrane, and the cell content flowed out (Fig. 3b). After 15 min, the cell surface contracted unevenly, with larger holes on the collapsed membrane, and cell contents overflowed. The cell broke into fragments, which gathered around and adhered to each other (Fig. 3c). HVPEF collapsed the morphology of S. enteritidis and induced the leakage of intracellular substances, in agreement with our previous research.

Scanning electron microscopy images of morphological changes of Salmonella enteritidis. Untreated S. enteritidis cells
ROS and MDA levels
The fluorescence intensity grew with time, notably after 9 min, and peaked at 15 min (Fig. 4a). The results showed that HVPEF treatment significantly induced ROS in cells, thus leading to damage to S. enteritidis cells. Previous research has found that extra ROS cause lipid peroxidation, oxidation of DNA, RNA, and thiol-containing proteins leading to cell death (Tardu et al., 2017; Xiao et al., 2019). Thus, the damage of HVPEF to S. enteritidis oxidative stress was explored through lipid peroxidation.
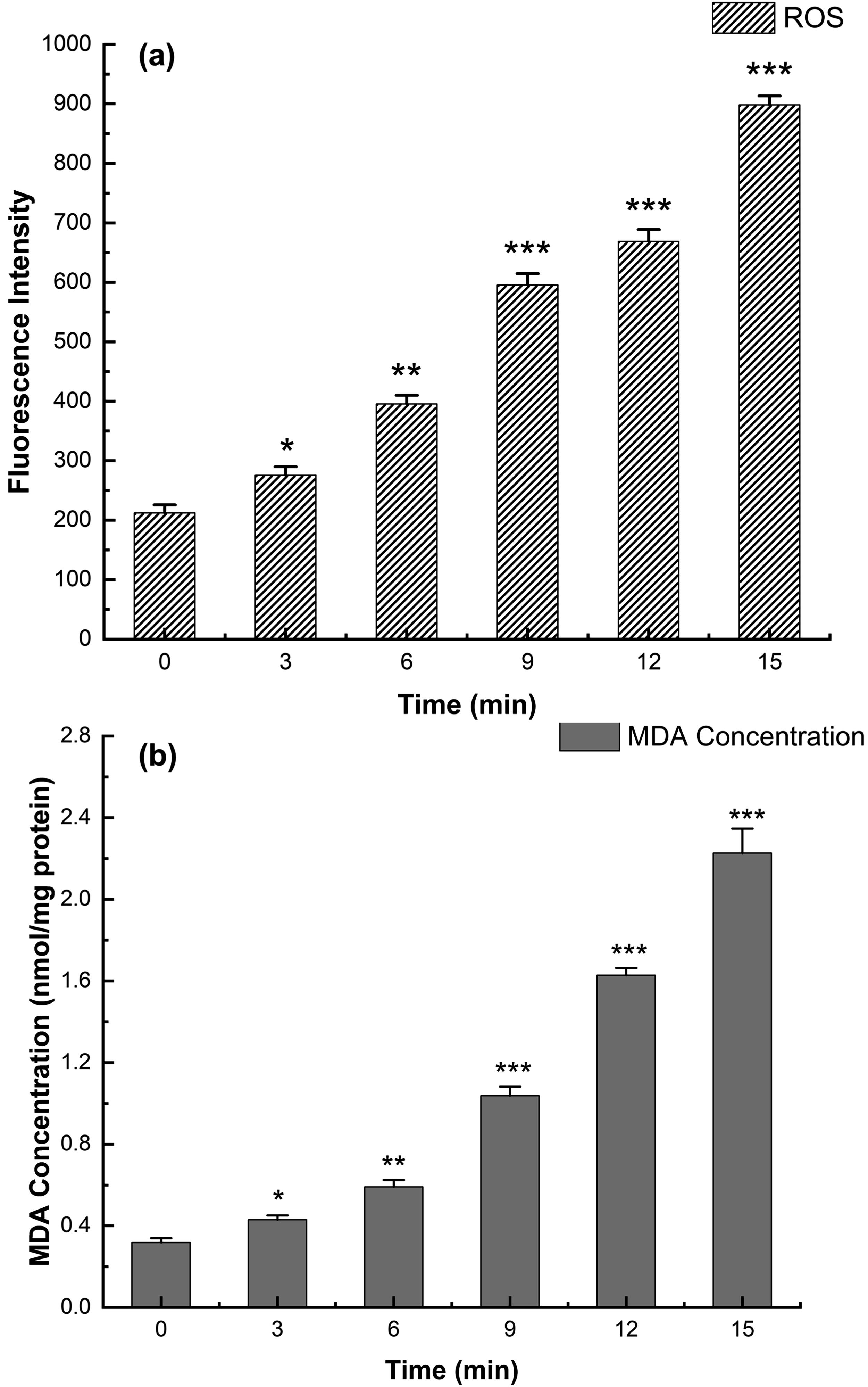
Changes of intracellular reactive oxygen species
The variations in MDA under HVPEF treatment are shown in Figure 4b. Compared with untreated cells (0.31 ± 0.02 nmol/mg protein), MDA in treated cells increased significantly after 3 min, reaching 2.23 ± 0.12 nmol/mg at 15 min, 7-fold more than the control group, in agreement with the trend of ROS. It was hypothesized that HVPEF caused an unbalanced generation and consumption of ROS and that excess ROS on the lipids in cell membranes exacerbated the level of cellular lipid peroxidation. Yun et al. found lipid peroxidation between free radicals and polyunsaturated fatty acids, which led to severe oxidative damage to cells (Yun et al., 2016). As reported, ROS can mediate the oxidation reaction of bases in DNA, which adversely affects the DNA structure (Cadet and Wagner, 2013). To validate the speculation, we observed DNA damage by flow cytometry.
DNA damage analysis
AO staining agent can penetrate intact cell membrane and fluoresces green when bound to double-stranded DNA, orange when bound to single-stranded DNA (Neeraja et al., 2017). Four quadrants Q1, Q2, Q3, and Q4 represented single-stranded DNA, single–double-stranded DNA, double-stranded DNA, and negative control, respectively (Fig. 5a). The cell proportion gradually increased in Q1 and decreased in Q3 with the prolongation of HVPEF treatment corresponding to Figure 5d and Figure 5b, which indicated that double-stranded DNA was gradually unwound. To sum up, HVPEF treatment induced the accumulation of ROS, caused lipid peroxidation, and unwounded DNA, which led to irreversible cell damage and death.
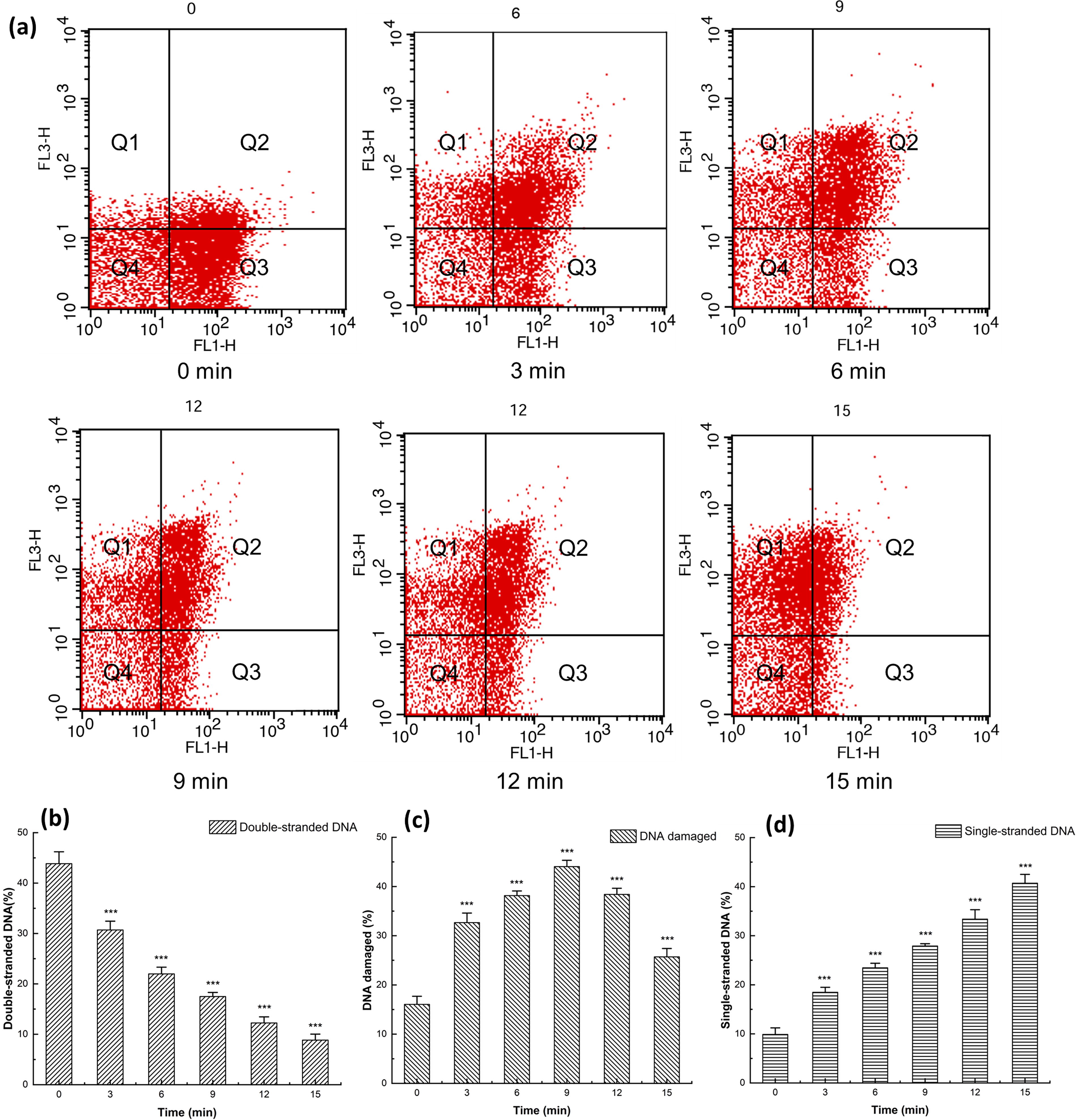
Flow cytometry of Salmonella enteritidis stained with AO after high-voltage prick electric field treatment at 13 kV for 0, 3, 6, 9, 12, and 15 min
Antibacterial effect on food surfaces
The mortality rate on all stuff increased in a positive correlation with treatment time, with the rate at 14 kV significantly higher than at 13 kV (Fig. 6). HVPEF reduced bacteria in salmon, chicken breast, sausage, and steak by 1.04 ± 0.07 lg CFU/cm2, 0.93 ± 0.08 lg CFU/cm2, 0.55 ± 0.03 lg CFU/cm2, and 0.48 ± 0.02 lg CFU/cm2, respectively, and the bactericidal rates reached 91 ± 1.6%, 88.2 ± 2.2%, 72.9 ± 2.7%, and 66.9 ± 1.6% at 14 kV for 30 min (Fig. 6). In contrast, stronger electric field and longer treatment time were required than on the plates, which was likely due to the complexity and heterogeneity of the food matrix as well as the food’s electrical conductivity (Saldana et al., 2014). In comparison with the smooth surface of the culture medium, the crude food surface served as a protection to S. enteritidis. Therefore, 14 kV voltage of HVPEF treatment for 30 min was used for subsequent experiments. As reported, ozone-free radicals, charged particle flow, and other bactericidal substances only acted on the surface of food rather than on cells penetrating into food (Critzer et al., 2007).
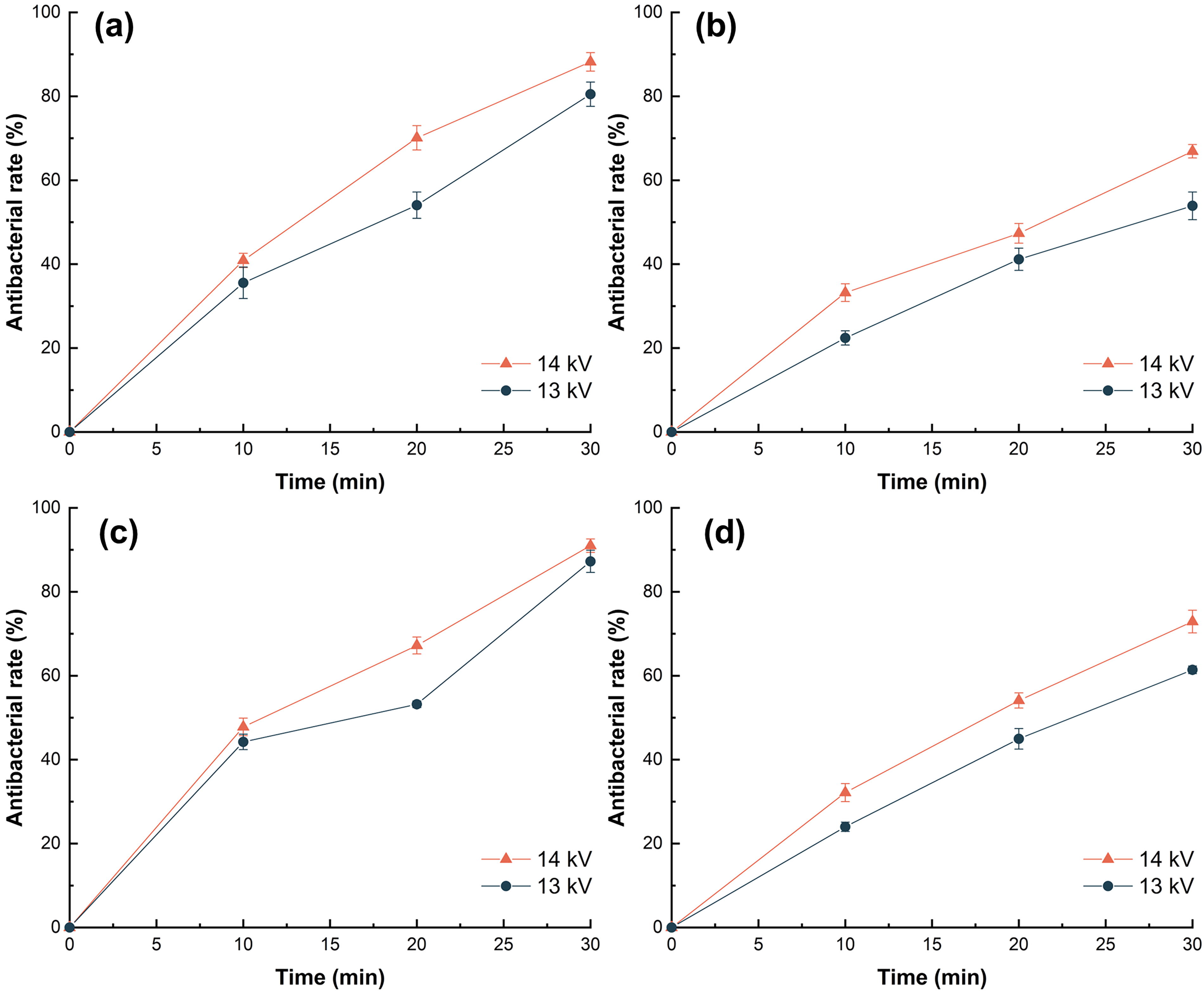
Mortality of Salmonella enteritidis on the surface of chicken breast
Storage quality of chicken breasts
Color and texture
There was no significant difference in the appearance of HVPEF-treated chicken breasts with increasing storage time (Fig. 7a). As shown in Table 1, L*, a*, and b* values decreased significantly after 6–10 days, but HVPEF treatment slowed down the process. There was no significant difference between the HVPEF-treated group in L* and a* values compared with the control group. However, b* values were significantly higher than the control group after 6–10 days, suggesting that HVPEF led to changes in the yellow-blue color. High-voltage electric field treatment caused denaturation of myofibrillar protein (Huang et al., 2023). Denatured myofibrillar protein led to a significant decrease in b* values during storage (Rahbari et al., 2020).

The quality indicators of chicken breasts treated with 0 and 14 kV electric field and stored at 4°C for 0, 2, 4, 6, 8, and 10 days, appearance
Effect of High-Voltage Prick Electric Field Treatment and Nontreatment on the Color Difference of Chicken Breast Meat After 0, 2, 4, 6, 8, and 10 Days
Where 0 kV group as a control is replaced by group C, *p < 0.05, **p < 0.01, and ***p < 0.001 indicate significant difference between storage 2, 4, 6, 8, and 10 days and storage 0 days.
# p < 0.05, ## p < 0.01, and ### p < 0.001 indicate significant difference compared with the control group.
As shown in Table 2, the hardness, elasticity, viscosity, and chewiness of chicken breasts treated by HVPEF decreased significantly after 6 days. The hardness of the HVPEF-treated group increased significantly after 4, 6, and 10 days compared with the control group. Qian et al. showed that electrostatic field treatment inhibited protease activity, maintained intact muscle fiber bundles in morphology, and delayed the hardness (Qian et al., 2022). Changes in elasticity and chewiness are associated with drop loss or connective tissue contents (Choe et al., 2015; Forrest et al., 1975).
Effect of High-Voltage Prick Electric Field Treatment and Nontreatment on the Texture of Chicken Breast Meat After 0, 2, 4, 6, 8, and 10 Days
Where 0 kV group as a control is replaced by group C, *p < 0.05, **p < 0.01, and ***p < 0.001 indicate significant difference between storage 2, 4, 6, 8, and 10 days and storage 0 days.
# p < 0.05, ## p < 0.01, and ### p < 0.001 indicate significant difference compared with the control group.
Total viable count and pH
Total viable count (TVC) values of chicken breasts increased but remained lower than the control group with storage time (Fig. 7b). The maximum limit of TVC in frozen and fresh poultry was 6 lg CFU/g (GB16869-2005); the TVC of the control group was 6.14 ± 0.04 lg CFU/g after 6 days, whereas the TVC of the HVPEF-treated group exceeded the limit after 10 days, in agreement with that electric field processing generates the corona discharge phenomenon and produces ozone, which inhibits the growth of microorganisms (Dalvi-Isfahan et al., 2023).
As shown in Figure 7c, the pH of chicken breasts gradually increased with storage time. Meat quality started to decay at pH > 6.2. The chicken breasts of the control group at pH 6.35 after 4 days, while the HVPEF-treated group began to deteriorate at pH 6.42 after 6 days. Obviously, HVPEF treatment delayed the spoilage process. Microbial growth and reproduction can break down proteins in chicken breast meat, which can degrade into alkaline nitrogenous substances such as ammonia and amines, causing the increase in pH values (Sujiwo et al., 2018).
TVB-N
The EC Regulation stipulated that TBV-N values ranging from 25 to 60 mg/100 g were beyond which the product was considered unfit for consumption. After 10 d, the TVB-N value of the control group reached 25.38 ± 0.88 mg/100 g, which exceeded the limit, whereas the TVB-N value of the treatment group was 19.25 ± 0.87 mg/100 g (Fig. 7d). Clearly, HVPEF treatment slowed down the TVB-N production during the initial storage period. Similarly, Ruan et al. found that the electric field treatment inhibited enzyme activity and reduced proteolysis (Ruan et al., 2019), which delayed the spoilage of chicken breasts during storage.
TBARS
The product was considered rancid if the MDA content exceeded 0.5–1.5 mg/kg (Ke et al., 1984). As shown in Figure 7e, chicken breasts of the control group were rancid after 6 days with an MDA content of 0.63 ± 0.023 mg/kg. The MDA value of the HVPEF-treated group was 0.54 ± 0.023 mg/kg after 8 days and chicken breasts were rancid. During storage, fat was constantly oxidized and decomposed, producing aldehydes, ketones, and other oxidation products, leading to further fat oxidation (Ko et al., 2016). Initially, TBARS values in the HVPEF-treated group were significantly higher than in the control group, perhaps due to the lipid oxidation caused by ozone from corona discharge on the meat surface (Gaebler et al., 2002). Together, HVPEF treatment can delay the oxidation of lipids and extend the shelf life of chicken breasts.
Conclusion
In conclusion, HVPEF effectively inactivated S. enteritidis on gelatin media, increased cell membrane permeability, and decreased membrane fluidity, which led to the leakage of intracellular proteins and nucleic acids. Moreover, HVPEF remarkably induced the increase of ROS in cells and caused lipid peroxidation and DNA damage, suggesting that oxidative stress contributed to the antibacterial effect of HVPEF. In addition, HVPEF exhibited marked bactericidal effects on S. enteritidis colonizing the surface of solid food (salmon, chicken breast, etc.), implying that HVPEF is applicable to food preservation and sterilization. However, HVPEF technology may cause energy loss. The effect of HVPEF treatment on the food nutrition needs further study.
Footnotes
Acknowledgments
This work was supported by On-budget Capital Construction Fund of Jilin Province Development and Reform Commission (2023C014). The authors gratefully acknowledge the fund supports.
Authors’ Contributions
H.Y.: Conceptualization, methodology, writing—reviewing and editing, writing—original draft, and data curation. S.L.: Writing—reviewing and editing, revision, validation, and investigation. Y.X.: Writing—reviewing and editing, validation, and investigation. Q.L.: Conceptualization and methodology. Y.Z.: Resources, validation, and investigation. Y.Y.: Visualization, supervision, revision, project administration, and funding acquisition.
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by On-budget Capital Construction Fund of Jilin Province Development and Reform Commission (2023C014).