
Abstract
Does temperature abuse during storage, distribution, marketing, and consumption of unpasteurized frozen açaí pulp increase microbial hazards? This study investigated the behavior of potentially pathogenic (Escherichia coli, Listeria monocytogenes and Salmonella spp.) and spoilage (mesophilic bacteria, yeasts and molds) microorganisms in two simulated thawing conditions: under refrigeration and at room temperature. The effect of repeated cold chain abuse was observed by thawing and refreezing (−20°C) açaí pulp four times over a period of 90 days. Freezing resulted in inhibition of all microorganisms except for mesophilic aerobic bacteria in one single sample. After thawing at 5°C, the kinetic parameters obtained by the Weibull model indicated that mesophilic aerobic bacteria, yeasts and molds and L. monocytogenes showed a longer inactivation time with δ values reaching 35, 126, and 46 days, respectively. The shortest inactivation time for a reduction of 4 log CFU.g−1 was for E. coli. The concentration of Salmonella spp. and L. monocytogenes in control samples was higher (p < 0.01) than in samples exposed to abusive conditions after 90 days of storage. The results indicate that the abusive thawing conditions studied do not increase the potential hazards of pathogens.
Introduction
Açaí is the most important dietary supplement and one of the main sources of income of the Amazon region. The consumption of the açai pulp was boosted in the 1990s, and it stood out as a “superfood” due to its high antioxidant activity (Yamaguchi et al., 2015). As it is a highly perishable fruit, freezing the pulp in its natural state is one of the main preservation methods.
The initial contamination of the fruit or pulp, processing without pasteurization, and improper storage conditions are considered critical factors for the safety and quality of the product (Bezerra et al., 2017). The palm tree of açaí can be contaminated from various sources, mainly because it serves as shelter and food for a variety of pollinating insects, birds, and other animals (Freitas et al., 2021; Oliveira et al., 2007; SEBRAE, 2013). After harvesting, storing, and transporting, the concentration of microorganisms in açaí fruit can reach 109 CFU.mL−1 if it is not processed within 40 h (Tribst et al., 2009). In the Amazon region, transportation often takes place in small boats during hours or even days, with the fruit stored inside or outside the holds (Aguiar et al., 2013).
Maintaining the cold chain of frozen açaí pulp from production to consumption is a challenge. Throughout the chain, the product may be exposed to abusive temperature without showing any noticeable sensory changes. Could it be that açaí pulp, if contaminated with potentially pathogenic microorganisms, undergoing freeze-thaw cycles could increase the potential hazards? Considering this question, the aim of this work was to study the behavior of potentially pathogenic and spoilage microorganisms simulating the thawing conditions of unpasteurized açaí pulp during storage.
Materials and Methods
Materials
Unpasteurized açaí pulps three different batches (A, B, and C) were produced in Manaus, Brazil. The pulps were frozen and stored in Styrofoam boxes with ice until they arrived in the laboratory and then kept at −20 ± 2°C until the experiments.
Pathogenic bacteria were obtained from the INCQS/FIOCRUZ collection (Brazil). The Escherichia coli cultures used were ATCC 11229, O45:H2 (CDC 00-3039), CDC H6, CDC H27, and CDC H34; Salmonella spp. cultures were Choleraesuis (ATCC 10708), Enteritidis (ATCC 13076), Typhimurium (ATCC 13311), Typhimurium (ATCC 14028), and S. brazil (Embrapa collection, isolated from açaí); and the Listeria monocytogenes cultures were 1/2b (CLIST 2032), 1/2a (CLIST 2044), 4b (CLIST 2045), 4b (CLIST 3436/Scott A), and 10403S (CLIST 4165). The strains were tested for cross-growth compatibility by genus, as aiming at not inhibiting each other during growth (Beuchat et al., 2001). The cultures were stored in Eppendorfs, diluted in 20% glycerol and frozen until the moment of the experiments.
Preparation of the inoculum and inoculation of açaí pulp
The cultures were activated in 10 mL of Tryptic Soy Broth (TSB; Becton, Dickinson & Company, USA) at 37°C for 24 h, with two new transfers in TSB afterward. The grown cultures were centrifuged (4000 rpm for 15 min at 4°C) twice. Then, the strains of each genus were resuspended in 0.1% peptone water (5 mL/strain) (Becton, Dickinson & Company) and combined in a single Falcon tube. The centrifuged cultures of E. coli, L. monocytogenes, and Salmonella spp. were resuspended and added to the same diluent until 10 mL was completed. The inoculum of each pathogen was obtained between 5 and 6 log CFU.mL−1, enumerated by drop plating on plate count agar (PCA; Merck, Germany) and also on selective media for each pathogen: on methylene blue eosin agar (EMB, Becton, Dickinson and Company) for E. coli, on deoxycholate–lysine–xylose agar (XLD, Merck) for Salmonella spp. and on Oxford agar (Becton, Dickinson and Company) for L. monocytogenes, in dilutions 0 to −5.
The açaí pulp samples (9.9 g) received 0.1 mL of each pathogen pool with initial concentration between 105 and 106 CFU.mL−1. The control sample (10 g) received no inoculum. Triplicate samples were homogenized and stored at −20 ± 2°C (slow freezing).
Microbiological analysis
After one week of inoculum and storage, microbiological analyses for prediction were performed to simulate the thawing stage in the industry. Continuing the simulation of thawing at distribution, marketing, and point of consumption, samples were thawed again after 30, 60, and 90 days of storage. Control samples (without inoculum) were analyzed under the same conditions. Control samples not subjected to thaw-freeze cycles were also analyzed after 90 days of storage.
Sample aliquots (1 g) thawed at 5 or 30°C were serially diluted (1:10) in 0.1% peptone water, followed by drop or surface plating (three triplicate dilutions) of 10 µL or 100 µL aliquots of the diluted sample, respectively. Analyses for mesophilic aerobic bacteria and yeasts and molds were performed on control samples, while pathogen analyses were performed on inoculated samples. The enumeration of E. coli, Salmonella spp., and L. monocytogenes was performed in their respective selective media and mesophilic aerobic bacteria in PCA (Merck, Germany) at 37°C/24 h, and yeasts and molds on Dichloran Rose Bengal Chloramphenicol Agar (DRBC, Merck, Germany) at 25°C/5 days (Salfinger and Tortorello, 2015).
Predictive modeling
Inactivation models were generated using the Geeraerd and Van Impe Inactivation Model Fitting Tool (GInaFiT) version 1.7 software (Geeraerd et al., 2005). The curves were adjusted by the Weibull model (Equation 1), as proposed by (Mafart et al., 2002):
Statistical analysis
The average results of the microbiological analyses (item 2.3) were statistically evaluated using the BioEstat software (version 5.3, Belém, Brazil) for analysis of variance (ANOVA). Data comparison was performed using the Tukey test with a significance level of 1%.
Results and Discussion
Predictive modeling of açaí pulp
Predictive microbiology in the present work aimed to describe the behavior of potentially pathogenic and spoilage microorganisms in unpasteurized frozen açaí pulp subjected to abusive thaw-freeze cycles, simulating thawing points along the value chain. All microorganisms showed inactivation behavior, with the exception of that observed in açaí C pulp contaminated with mesophilic aerobic bacteria. In this case, the spoilage bacteria showed a tendency toward stability at the two temperatures studied (data not shown). From the experimental data, the model that best fit the data to describe the microbial inactivation curves was the nonlinear Weibull model (Mafart et al., 2002).
Table 1 shows the kinetic parameters of the pathogenic and spoilage microorganisms studied in three batches of açaí pulp, which were subjected to freeze-thaw cycles. The data that best fit the model were obtained from pulp samples containing mesophilic bacteria (0.9999), L. monocytogenes (0.9993 and 0.9991), and E. coli (0.9990) at 5°C with R 2 close to 1.
Kinetic Parameters of Potentially Pathogenic and Spoilage Microorganisms in Three Batches of Unpasteurized Frozen Açaí Pulp Subjected to Successive Abusive Storage Conditions, a Adjusted to the Weibull Model
Thawing in two simulated conditions, under refrigeration at 5°C and at room temperature of 30°C, followed by three refreezes (−20 ± 2°C) at successive intervals of 1 month.
δ: Time (day−1) required for first decimal reduction. p: Parameter related to the inactivation curvature. t 4D: Time (day) for inactivation of 4 logarithmic cycles.
Parameter values are represented as mean ± standard deviation.
The time required to reduce the initial population (δ) in açaí pulp at 5°C can be compared between pathogenic microorganisms. L. monocytogenes presented the highest delta values, ranging from 31.99 to 46.05 days (Table 1). The δ of L. monocytogenes was about 3.5 times higher than that of Salmonella spp., and reached 6 times that of E. coli, on average. The delta values of L. monocytogenes in açaí pulp also revealed that the time required to reduce the initial population at 5°C was 3 to 4 times longer than at 30°C. The psychrotrophic nature of L. monocytogenes can be correlated with its ability to develop at refrigeration temperatures. It also took longer for the first decimal reduction of L. monocytogenes to occur at 5°C than for E. coli and Salmonella spp. Compared with the spoilage microorganisms, yeasts and molds in açaí pulp showed the highest δ (Table 1), as they required 72.06 to 126.34 days for the first decimal reduction to occur, showing population stability in the face of freeze-thaw cycles at 5°C and 30°C.
The downward curvature (p > 1) of L. monocytogenes in açaí pulps at 5°C (Fig. 1) showed that the remaining cells were damaged, and, in such a situation, less time would be needed to inactivate the same population in relation to the cells still surviving. When the samples reached 30°C, L. monocytogenes showed the opposite behavior (p < 1), according to Table 1, with an upward curvature. In this case, the most sensitive cells were promptly destroyed, while the survivors showed the ability to adapt to the stressful environment. This demonstrates that açaí pulp contaminated by L. monocytogenes after thawed-refrozen at a lower temperature presented a progressive increase in the inactivation of the remaining population. Regarding the behavior of the other pathogens and deteriorants, all showed an upward curvature where p < 1, also considering the standard deviations of the data obtained (Table 1).

Listeria monocytogenes (LM) inactivation curves in açaí pulps A (5°C: ◊ and black line; 30°C: □ and red line), B (5°C: ▽ and yellow line; 30°C: ∗ and blue line), and C (5°C: + and orange line; 30°C: ○ and green line) after thawing at 5°C and 30°C adjusted by the Weibull model.
In a study of inactivation and viability of pathogens in pressurized açaí juice, Gouvea et al. (2020) found that in the control sample (nonpressurized açaí juice, pH 4.3 and 2.9°Brix) L. monocytogenes and Salmonella spp. remained viable at concentrations of 7.33 ± 0.22 and 7.43 ± 0.17 log CFU.mL−1 (no significant difference in relation to the initial day, p > 0.05), respectively, after 42 days at 7°C. During this period, the logarithmic reductions for L. monocytogenes were 0.21 CFU.mL−1 and 0.49 CFU.mL−1 for Salmonella spp. These data demonstrate the survival capacity of pathogenic microorganisms in açaí under refrigerated conditions for almost 2 months of storage. In comparison with the present study, when subjecting L. monocytogenes and Salmonella spp. to thaw-freeze cycles, unlike Gouvea et al. (2020) have observed, the cells end up suffering injuries that promote microbial inhibition over time, and also difficult cell recovery, as it can be seen by the value of p > 1 for L. monocytogenes at 5°C.
The time required to reduce 4 logarithmic cycles (t 4D) cannot be estimated in all cases, as it would be too long to be calculated (Table 1). However, it can be seen that the shortest and longest t 4D times of E. coli in açaí pulp were 1.8 (C pulp/5°C) and 47.4 days (A pulp/30°C), respectively. For Salmonella spp., the shortest t 4D time was 54.0 days (B pulp/30°C), while the longest time was 86.4 days (A pulp/5°C). The inactivation time can only be observed in L. monocytogenes at 30°C, ranging from 68.4 days in C pulp to 81.9 days in A pulp to reduce 4 log CFU.g−1. Among the pathogens, the average of the data revealed that in the thaw-freeze cycles (5°C and 30°C) of açaí pulp, E. coli was more influenced than Salmonella spp., followed by L. monocytogenes, which was the most stable pathogen, regarding t 4D values.
At 90 days, after thaw-freeze cycles, there was a similar average reduction of L. monocytogenes in the açaí pulp at 5°C and 30°C, being 3.44 and 3.74 log CFU.g−1, respectively (Fig. 1). In 60 days, E. coli decreased 5.30 log CFU.g−1, on average, relative to the initial inoculum at 5°C and 30°C, with the exception of the C pulp at 5°C which had an estimated value <1.00 log CFU.g−1 after the first thawing (Fig. 2). E. coli suffered the greatest log inactivation in the shortest time. Salmonella spp. reduced 4.49 log CFU.g−1 after three thaws (60 days) at 30°C, while the reduction at 5°C was 4.35 log CFU.g−1 after four thaws (90 days) (Fig. 3). To evaluate the adherence, survival, and recovery of Salmonella spp. in açaí fruits, Bezerra et al. (2015) inoculated a concentration of 10 log CFU.mL−1 and observed an inactivation of 4 log CFU.mL−1 using the inoculum dripping method on fruit with drying in laminar flow.
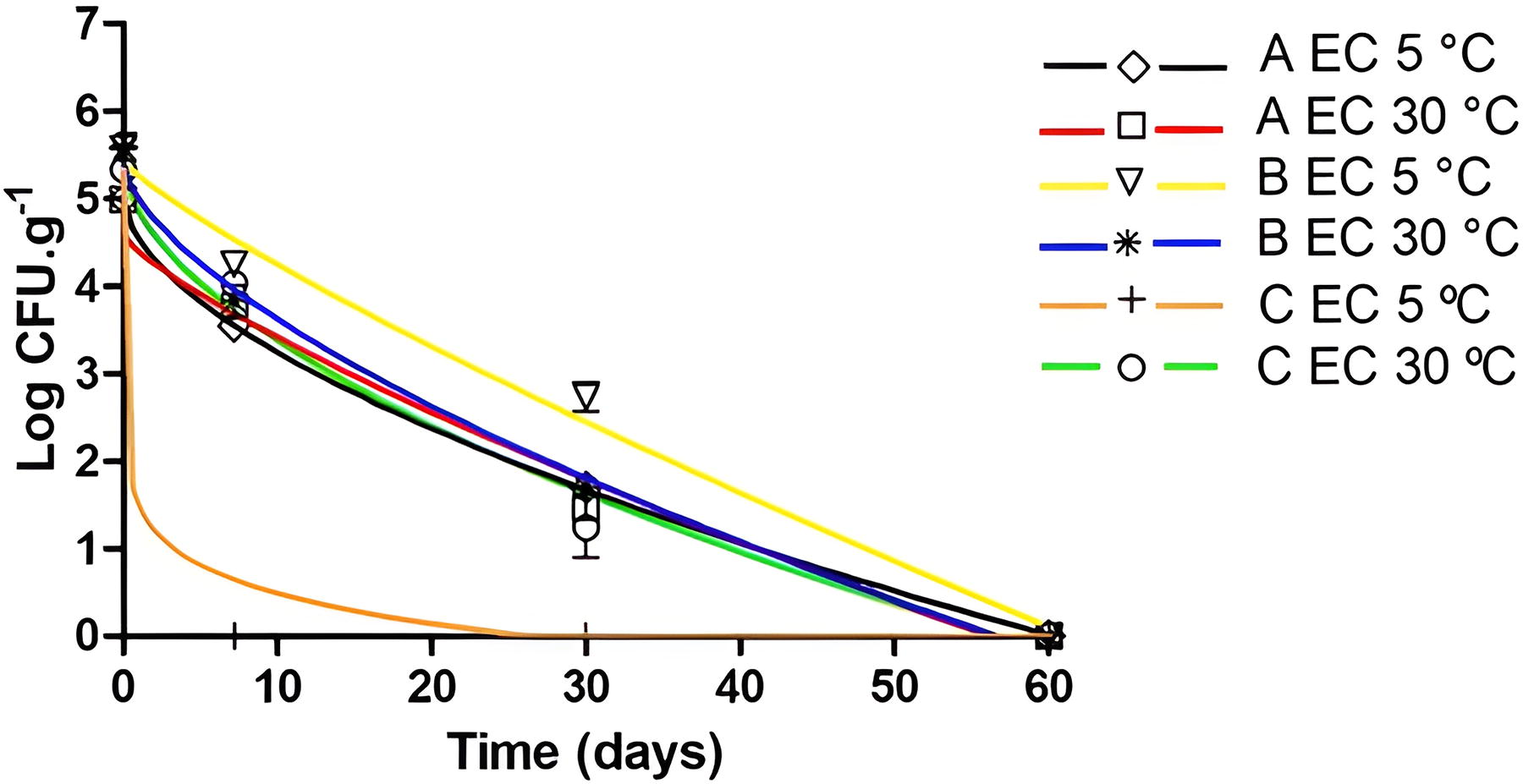
Escherichia coli (EC) inactivation curves in açaí pulps A (5°C: ◊ and black line; 30°C: □ and red line), B (5°C: ▽ and yellow line; 30°C: ∗ and blue line), and C (5°C: + and orange line; 30°C: ○ and green line) after thawing at 5°C and 30°C adjusted by the Weibull model.
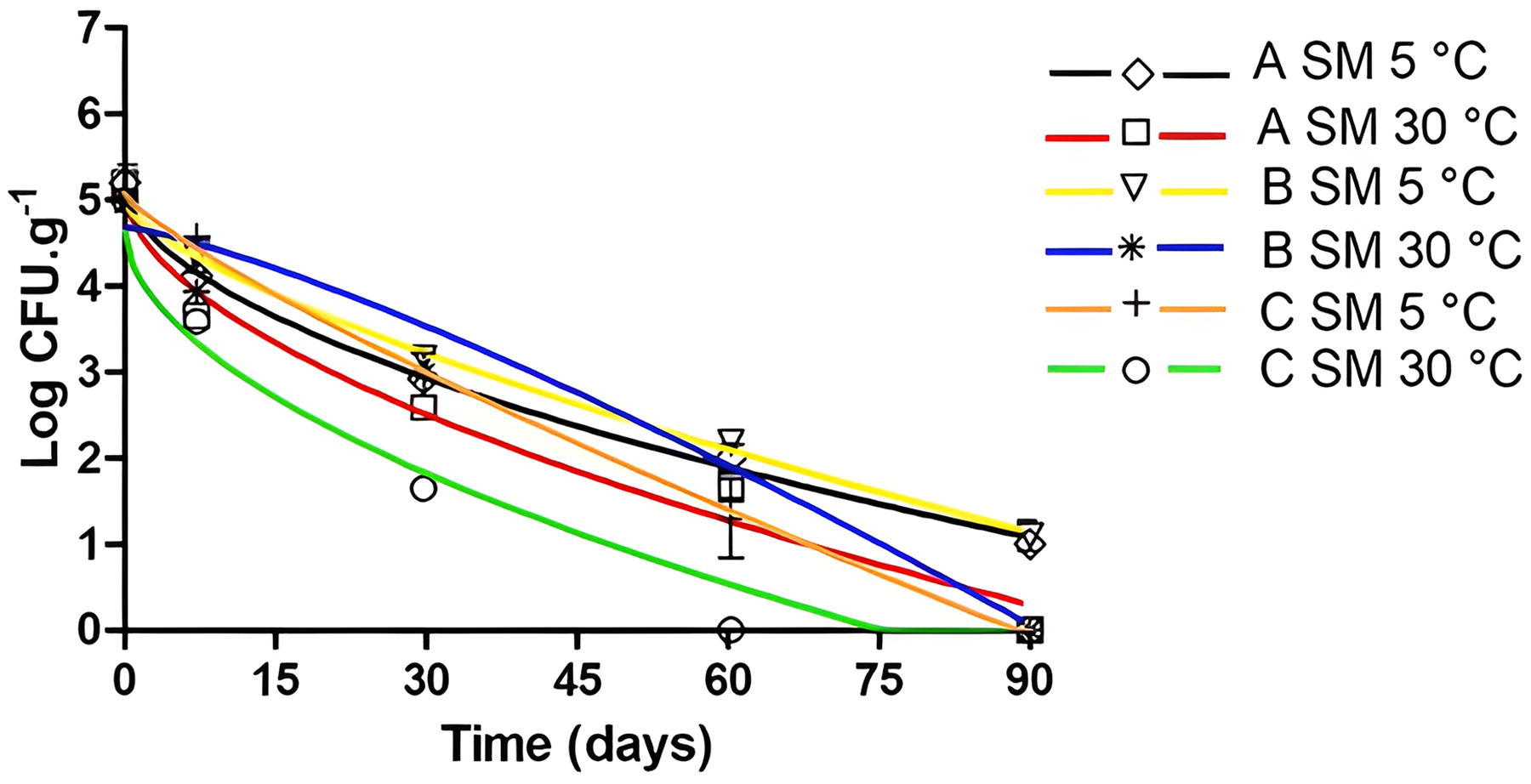
Salmonella spp. (SM) inactivation curves in açaí pulps A (5°C: ◊ and black line; 30°C: □ and red line), B (5°C: ▽ and yellow line; 30°C: ∗ and blue line), and C (5°C: + and orange line; 30°C: ○ and green line) after thawing at 5°C and 30°C adjusted by the Weibull model.
The frozen açaí pulps, when obtained, presented endogenous contamination by mesophilic aerobic bacteria (3.10 log CFU.g−1 in C pulp − 5.48 log CFU.g−1 in A pulp), and yeasts and molds (3.56 log CFU.g−1 in B pulp — 3.70 log CFU.g−1 in C pulp). In commercial açaí pulps analyzed by Bandeira et al. (2023), the mesophilic aerobic bacteria counts determined were close to those seen here, being 1.4 × 102 CFU.g−1 and 7.7 × 104 CFU.g−1. In 90 days of storage, the observed logarithmic reduction of mesophilic aerobic bacteria was, on average, 1.74 log CFU.g−1 at 5°C and 2.36 log CFU.g−1 at 30°C (Fig. 4), while for yeasts and molds, it was 0.75 log CFU.g−1 at 5°C and 0.98 log CFU.g−1 at 30°C (Fig. 5). These data show that the most persistent microorganisms, even compared with the pathogens tested, are the yeasts and molds that showed a logarithmic reduction of <1 Log throughout the 90-day thaw-freeze cycles of the açaí pulp at 5°C and 30°C. Sousa et al. (2006) also observed the survival of yeasts and molds in frozen açaí juice (−18°C) for 90 days, verifying the same initial count (>4.2 × 104 CFU.mL−1) after storage.
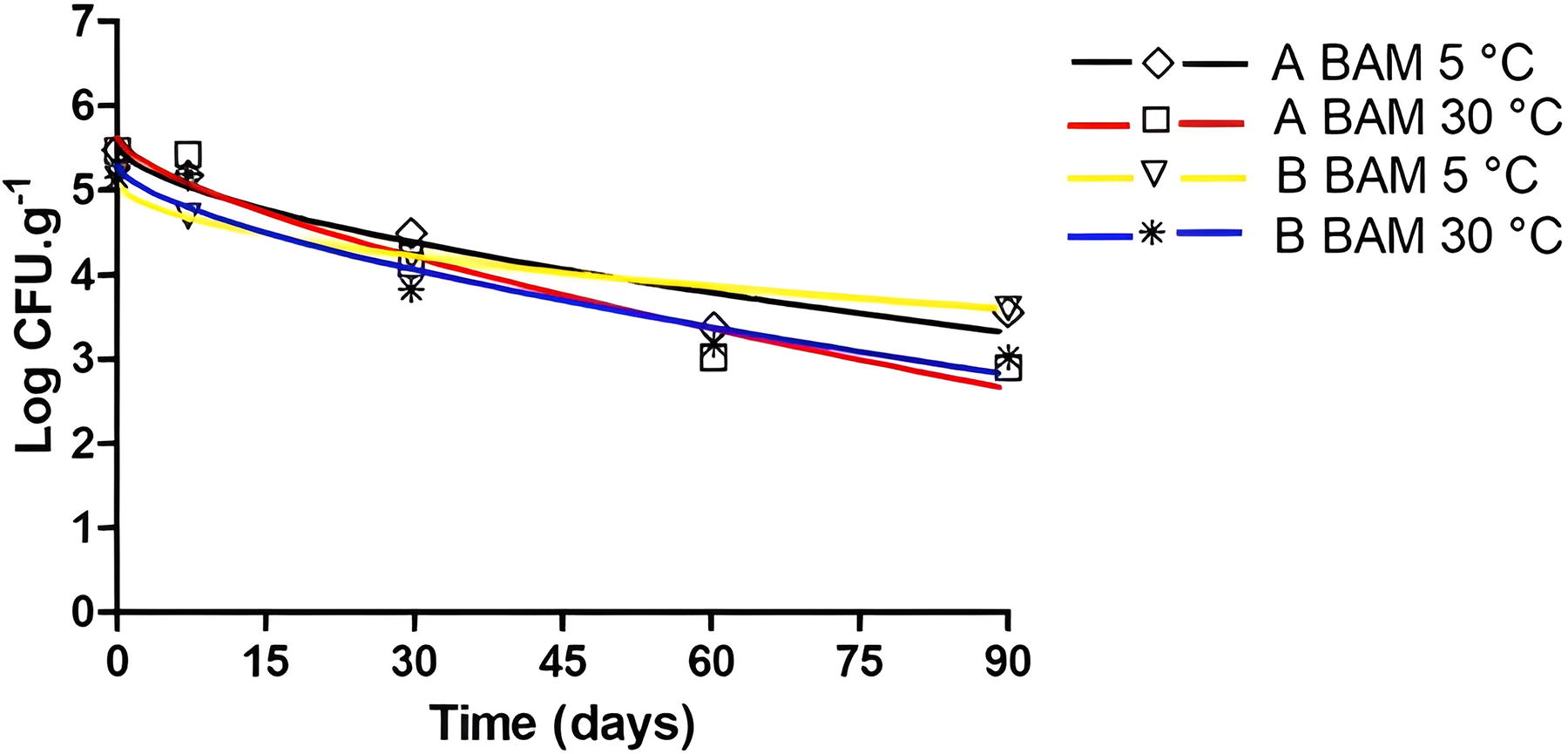
Mesophilic aerobic bacteria (BAM) inactivation curves in açaí pulps A (5°C: ◊ and black line; 30°C: □ and red line) and B (5°C: ▽ and yellow line; 30°C: ∗ and blue line) after thawing at 5°C and 30°C adjusted by the Weibull model.
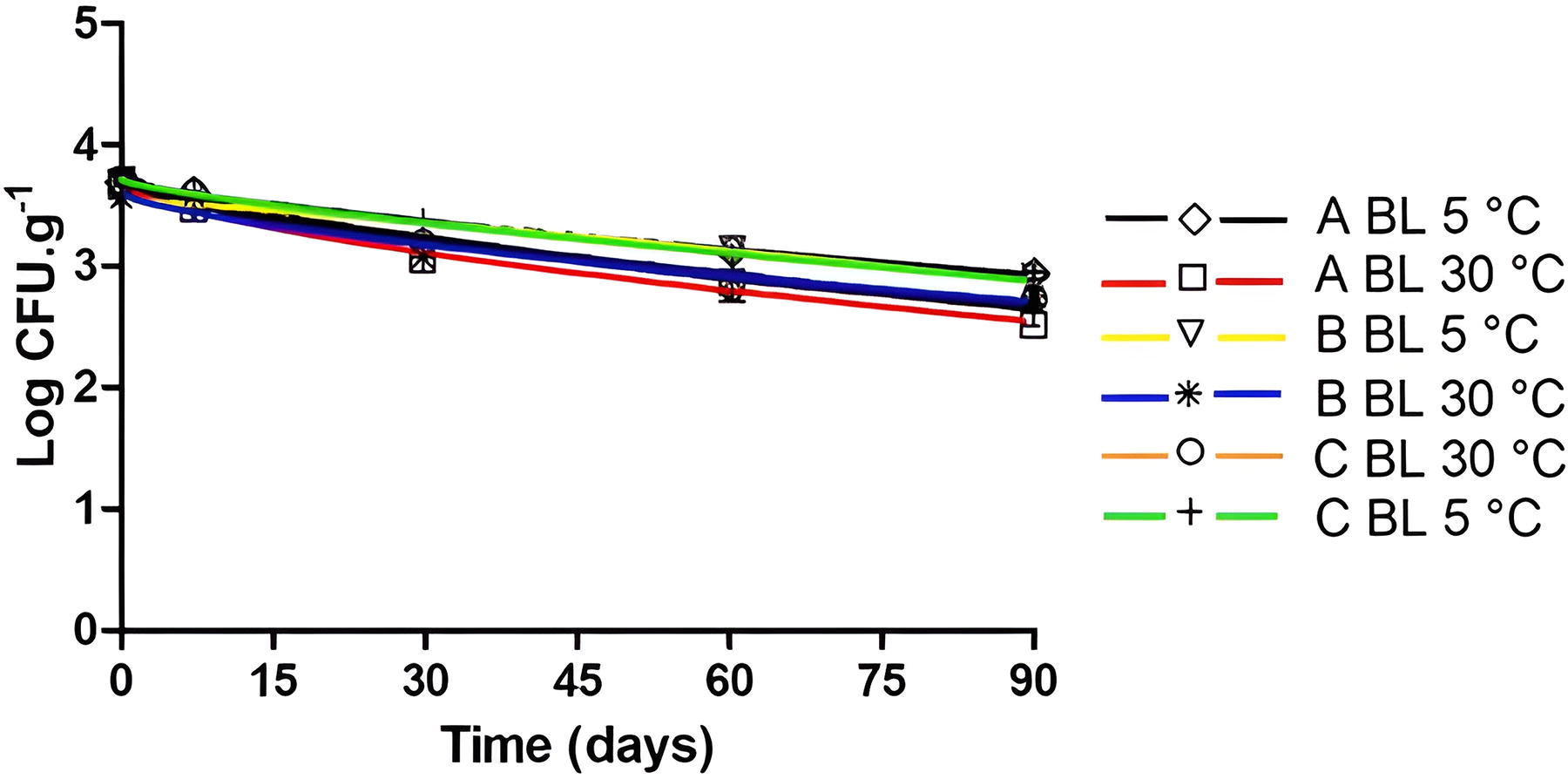
Yeasts and molds (BL) inactivation curves in açaí pulps A (5°C: ◊ and black line; 30°C: □ and red line), B (5°C: ▽ and yellow line; 30°C: ∗ and blue line), and C (5°C: + and green line; 30°C: ○ and orange line) after thawing at 5°C and 30°C adjusted by the Weibull model.
Microbial concentration in açaí pulp: Samples subjected to abusive conditions × control
At the end of the study, the microbial concentration of açaí pulps after thaw-freeze cycles was compared with the counts of samples that remained frozen at −20°C for the same period (control), as shown in Fig. 6. After 60 days, E. coli showed no count (0.00 log CFU.g−1) in the pulp samples thawed to 5°C and 30°C, while in control pulp, the average was 0.55 ± 0.02 log CFU.g−1, with no significant difference between the different conditions.
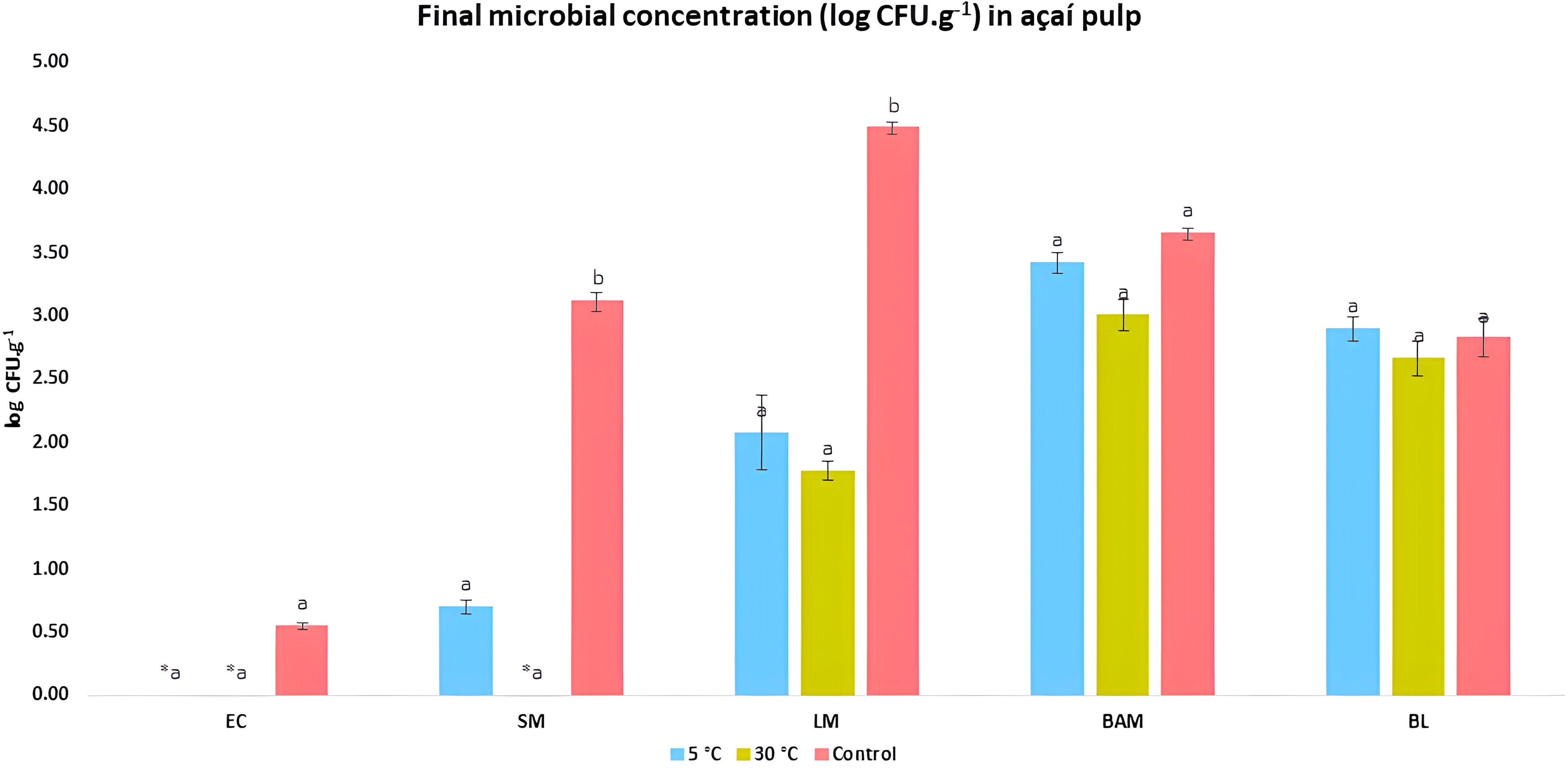
Comparison of the final microbial concentration (log CFU.g−1) of EC, SM, LM, BAM, and BL in açaí pulp samples subjected to successive thawing at 5°C and 30°C, and control açaí pulp samples, which were not subjected to thawing for up to 90 days. *0.00 log CFU.g−1. **Lowercase letters represent comparison between bars with a significance level of 1% using the Tukey test. Analyzes of each batch were performed in triplicate, and the data were presented as the average of the batches.
The control samples with Salmonella spp. and L. monocytogenes showed a significant difference (p < 0.01) with higher concentrations than those of the samples subjected to abusive conditions, after 90 days of freezing. In the açaí pulp with Salmonella spp. there was an average of 0.70 ± 0.06 log CFU.g−1 at 5°C and no count at 30°C, while in the control pulp there was 3.12 ± 0.08 log CFU.g−1. Likewise, it can be observed that the pulp containing L. monocytogenes presented an average of 2.08 ± 0.30 log CFU.g−1 at 5°C, 1.78 ± 0.07 log CFU.g−1 in the pulp at 30°C, while in the control pulp the count was 4.49 ± 0.04 log CFU.g−1. These data show that keeping açaí pulp frozen, without thaw-freeze cycles in the production chain, for at least 90 days increases the chances of survival of the Salmonella spp. and L. monocytogenes.
Archer (2004) stated that freezing is generally an excellent way to preserve the viability of microorganisms. However, repeated freeze-thaw cycles can disrupt and destroy bacteria. Zhang et al. (2021) analyzed the recovery of E. coli O157:H7 in TSB after 12 freeze-thaw (−20 and 25°C) cycles. After the first thaw, the recovery was 58.8% of the initial inoculum, while at the end of all cycles the recovery of E. coli O157:H7 was 11.4%.
Freezing technology immobilizes water molecules to preserve food at low temperatures. During the freezing, water molecules pass into a solid state, forming ice crystals, while solutes concentrate in the still-thawed fraction and dehydrate the microbial cell. Chemical and enzymatic processes also begin to be inhibited. By osmosis, the water still present tends to migrate to the more concentrated medium, compromising membrane permeability and causing irreparable damage (Archer, 2004; El-Kest and Marth, 1992). In addition, slow freezing results in the appearance of larger ice crystals, increasing damage to microbial cells (Jeremiah, 2019). Thawing generally occurs at a slower rate than freezing (food remains in the zone below 0°C longer) favoring the process of recrystallization, where smaller ice crystals melt and refreeze to form larger ice crystals (Potter, 1986). All these effects of freeze-thaw cycles contribute to cell death, as seen here.
Regarding the spoilage groups, there were no significant differences between the samples subjected to freeze-thaw cycles at 5°C and 30°C compared with the control samples at the end of the 90-day study period. The average count verified for mesophilic aerobic bacteria at 5°C, 30°C, and control was, respectively, 3.42 ± 0.08 log CFU.g−1, 3.01 ± 0.13 log CFU.g−1, and 3.65 ± 0.05 log CFU.g−1. For yeasts and molds, the average count was 2.90 ± 0.10 log CFU.g−1 at 5°C, 2.67 ± 0.14 log CFU.g−1 at 30°C, and 2.82 ± 0.15 log CFU.g−1 in the control pulp.
Gram-negative bacteria are more influenced by freezing than Gram-positive bacteria (Rahman and Velez-Ruiz, 2020), as observed in the results of E. coli and Salmonella spp. in contrast to L. monocytogenes (Fig. 6). Gene activation and repression mechanisms are initiated during the change in the physical state of microorganisms. Freeze-thaw makes L. monocytogenes more susceptible to the action of lipase and lysozyme enzymes (El-Kest and Marth, 1992). Yeast resistant to freeze-thaw cycles produces high levels of trehalose, which performs a cryoprotective function (Rahman and Velez-Ruiz, 2020). Missous et al. (2010) observed the sensitivity of yeasts and molds to freezing (−10°C) and verified the resistance (56–75% survival) of yeasts and a mold strain by the production of arthrospores. By activating defense mechanisms, even with sublethal lesions, microorganisms can resist adverse environmental conditions, adapt, repair lesions, and extend freeze-thaw survival.
Throughout the açaí pulp production chain, the risks of microbial contamination are eminent and often unavoidable by actual practices. Based on the results obtained, pasteurization of the açaí pulp is recommended to reduce any microbial hazards and maintain product safety. In addition, pre-harvest, post-harvest, and processing of açaí pulp (artisanal and industrial) must be carried out applying good practices to prevent possible contamination. Prevention and control recommendations were documented for the application of HACCP (Hazard Analysis and Critical Control Points) of the açaí pulp (SEBRAE, 2013).
Conclusions
Over 90 days, abusive conditions promoted a greater reduction in microbial hazards compared to non-abusive conditions. The results reinforce the importance of pasteurization, as freezing or abusive storage conditions do not eliminate all potentially pathogenic microorganisms studied to safe levels. In this work, only in samples with E. coli subjected to abusive conditions at 5°C and 30°C, it was possible to observe inhibition at safe levels, with a reduction higher than 5.00 log CFU.g−1.
Footnotes
Acknowledgment
The authors acknowledge support from the Coordination for the Improvement of Higher Education Personnel (CAPES) for the scholarship and the Brazilian Agricultural Research Corporation (Embrapa) for their financial support to the project.
Authors’ Contributions
Conceptualization: L.M.C.C., E.H.M.W., J.C.A.S.S., and V.M.D.M. Formal analysis: J.C.A.S.S. and G.L.D.P.A.R. Investigation: J.C.A.S.S. Methodology: J.C.A.S.S., G.L.D.P.A.R., and E.H.M.W. Project administration: J.C.A.S.S., L.M.C.C., E.H.M.W., and V.M.D.M. Supervision: L.M.C.C., E.H.M.W., and V.M.D.M. Validation: J.C.A.S.S. Writing (original draft): J.C.A.S.S. Writing (review and editing): L.M.C.C., E.H.M.W., G.L.D.P.A.R., J.C.A.S.S., and V.M.D.M.
Disclosure Statement
All authors of this publication reported having no conflicts of interest.
Funding Information
This work was supported by CAPES and Embrapa.