
Abstract
The development of novel antibacterial agents from plant sources is emerging as a successful strategy to combat antibiotic resistance in pathogens. In this study, we systemically investigated the antibacterial activity and underlying mechanisms of baicalin against methicillin-resistant Staphylococcus aureus (MRSA) and Stenotrophomonas maltophilia. Our results showed that baicalin effectively restrained bacterial proliferation, compromised the integrity of cellular membranes, increased membrane permeability, and triggered oxidative stress within bacteria. Transcriptome profiling revealed that baicalin disrupted numerous biological pathways related to antibiotic resistance, biofilm formation, cellular membrane permeability, bacterial virulence, and so on. Furthermore, baicalin demonstrated a synergistic antibacterial effect when combined with ampicillin against both MRSA and S. maltophilia. In conclusion, baicalin proves to be a potent antibacterial agent with significant potential for addressing the challenge of antibiotic resistance in pathogens.
Introduction
Antibiotic resistance presents an urgent problem among pathogens. Natural (Chen et al., 2022; Kong et al., 2010) and chemically synthesized (Wang et al., 2023; Wilson Lucas et al., 2023; Zhang et al., 2018) antibacterial active substances are urgently needed to substitute traditional antibiotics and address the issue of antibiotic resistance. Researchers have paid great interest in natural active substances derived from medicinal plants with remarkable antibacterial activity, which are thought to have great potential to replace conventional antibiotics. Taking chlorogenic acid (Chen et al., 2022), protocatechualdehyde (Tian et al., 2021), and berberine (Kong et al., 2010), for example, they have been widely demonstrated to possess excellent antibacterial activities against common foodborne pathogens. In addition, baicalin is another crucial bioactive compound with excellent antibacterial properties. Baicalin is a natural flavonoid derived from the dried root of Scutellaria baicalensis (Liu et al., 2018). The antibacterial activities of baicalin are mainly concentrated on Escherichia coli, Staphylococcus aureus, S. saprophyticus, and Pseudomonas aeruginosa (Miao et al., 2021; Wang et al., 2021; Zhang et al., 2021).
Methicillin-resistant S. aureus (MRSA), the predominant drug-resistant bacteria, exhibits elevated incidence and fatality rates, with initial documentation dating back to 1961 in the United Kingdom (JEVONS et al., 1962; Jin et al., 2020). MRSA is recognized for its multidrug resistance to conventional antibiotics, including macrolides, linezolid, aminoglycosides, and others, presenting substantial struggles for the cure of MRSA infections (Han et al., 2007; Kali, 2015; Schaefler et al., 1981; Schito, 2006). The primary cause of its antibiotic resistance stems from staphylococcal chromosome cassette mec, a mobile genetic element ranging from 21 to 67 kb, which contains the resistant genes mecA and mecC.
Meanwhile, Stenotrophomonas maltophilia, a Gram-negative rod-shaped bacterium that exhibits resistance to various drugs, especially the broad-spectrum antibiotics of the carbapenem class (Spencer, 1995), is responsible for causing opportunistic infections in patients with weakened immune systems (Brooke, 2012). Many antibiotics are used to treat S. maltophilia infection, such as trimethoprim, sulfamethoxazole, and fluoroquinolone (Cho et al., 2014; Chung et al., 2013). However, the multidrug-resistance pumps and plasmids and gene transfer mechanisms related to antibiotic resistance in S. maltophilia make it difficult to cure its infection (Nakamura et al., 2021).
Nowadays, studies related to the antibacterial activity of baicalin against MRSA and S. maltophilia are limited. Therefore, in this study, we first explored the antibacterial activities and the related antibacterial mechanisms of baicalin on MRSA and S. maltophilia. Specifically, after baicalin treatment, changes in antibiotic resistance, biofilm formation, bacterial morphology, and bacterial membrane permeability of MRSA and S. maltophilia were comprehensively analyzed. Finally, we compared the effects of baicalin on these two bacteria at the transcriptomic level. This study provides new ideas for addressing the antibiotic resistance of MRSA and S. maltophilia.
Materials and Methods
Sample preparation
MRSA (ATCC 43300) used in subsequent experiments was cultured in Luria–Bertani (LB) broth for 12 h at 150 rpm, 37°C. S. maltophilia (ATCC 17666) used in subsequent experiments was cultured in Soya Casein Digest Lecithin Polysorbate (SCDLP) broth for 48 h at 150 rpm, 30°C. Then, cultures were centrifuged for 10 min at 6000 rpm, 4°C. The resulting cell pellets were harvested, washed thrice with PBS, and homogenized in PBS for subsequent use.
Baicalin (98%, CAS 21967-41-9) was dissolved in dimethyl sulfoxide (DMSO) and stored at 4°C for standby use.
Antibacterial activity of baicalin
The antibacterial activity of baicalin was determined based on Xue et al (Xue et al., 2022) protocol with some modifications. Baicalin was mixed with sterilized LB or SCDLP agar in various concentrations. Following solidification, the diluted bacterial solution was spread on each agar plate. Colony count was recorded by Fully Automatic Colony Counter (Beijing Dequan Xingye Commerce and Trade Co. Ltd., China) after incubation.
Biofilm development evaluation
Antibiofilm activity of baicalin on pre-developed biofilm of bacteria
In short, overnight cultures (about 108 colony-forming unit or 108CFU/mL) were seeded into a 96-well plate and incubated statically for 24 h to develop bacterial biofilm. Subsequently, the biofilms underwent a PBS wash and were exposed to different treatments for 30 min. Hereafter, the wells were processed according to Shi et al. (2018), and the absorbance was measured at 595 nm.
Antibiofilm activity of baicalin on biofilm formation of bacteria
Similarly, the effects of baicalin on the biofilm formation of these two bacteria were also conducted (Maury et al., 2019). Equal amounts of bacterial solution and different concentrations of baicalin were mixed and cultured for 24 h. Then, biofilm quantification was conducted by crystal violet staining as above.
Ultrastructural analysis
Scanning electron microscopy
After 30 min of baicalin (6 mg/mL) and DMSO treatment, bacteria were harvested and fixed in 2.5% glutaraldehyde overnight at 4°C (Fang et al., 2018). Then, bacteria were processed by standard procedures and visualized by scanning electron microscopy (SEM) (JSM-7500F; Hitachi, Japan).
Transmission electron microscopy
Bacteria after the same treatment as above were treated according to standard procedures (Li et al., 2019), and the intercellular morphology of bacteria was visualized by transmission electron microscopy (TEM) (JEM-1200EX; JEPL Ltd., Japan).
Permeability analysis of bacterial membrane
Live/dead cell staining
Cell membrane permeability was initially tested by the LIVE/DEAD BacLight Bacterial Viability Kit (L7012; Invitrogen, USA) (Watson et al., 2019). The bacteria solution was treated with 6 mg/mL baicalin at different times. Then, bacteria were stained with mixed dyes of Propidium Iodide (PI) and SYTO-9 and observed using a TCSsp5II laser scanning confocal microscope (Agilent, USA).
Release of intracellular components
Release of components within bacteria, such as potassium ion (K+) and DNA, was also commonly used to reflect cellular membrane permeability. The K+ contents in cell-free supernatants were detected by an atomic absorption spectrophotometer (TAS-990; China) at 766.5 nm (Lou et al., 2011), and the DNA concentrations in cell-free supernatants were determined by spectrophotometry at 260 nm.
Assessment of oxidative stress
Reactive oxygen species (ROS) contents generated in bacteria were measured by the ROS assay kit (Wang et al., 2020). First, bacteria were treated with 6 mg/mL baicalin for 30 min. Then, bacteria were loaded with a 2′,7′-dichlorofluorescein diacetate fluorescent probe for 1 h in the dark. Finally, the fluorescence intensity of bacteria was detected by a fluorescence spectrophotometer (Hitachi HT-7000; Japan).
Transcriptomic profiling
Bacteria treated with DMSO and 6 mg/mL baicalin were transcriptome sequenced by Shanghai Majorbio Bio-pharm Technology Co., Ltd. RNA-seq data for MRSA and S. maltophilia were deposited in the NCBI Sequence Read Archive under accession numbers PRJNA1101445 and PRJNA1101463, respectively. Finally, six and five genes were selected from MRSA and S. maltophilia for quantification using quantitative real-time polymerase chain reaction (qRT-PCR), respectively. All the primers used in qRT-PCR were listed in Supplementary Table S1.
Decrease in bacterial ampicillin resistance
Evaluation of minimum inhibition concentrations
The minimum inhibition concentrations (MICs) of baicalin and ampicillin against bacteria were determined (Qiao and Sun, 2014). Wells treated with DMSO acted as the negative control, while wells without bacteria acted as the blank control. MICs were identified as the lowest concentration that inhibited bacterial growth by over 90% relative to the negative control. The inhibition rate was calculated using the following formula:
Checkerboard assay
The conjunct effects of baicalin and ampicillin against bacteria were evaluated by checkerboard assay (Yu et al., 2005). The fractional inhibitory concentration (FIC) values were calculated using the above MIC values by the formula provided:
The lowest FIC was termed the FIC index (FICI). Synergy, antagonism, additivity, and indifference were defined based on FICI values: ≤0.5 indicated synergy, >2 indicated antagonism, 0.5< FICI ≤1.0 indicated additivity, and 1.0< FICI ≤2.0 indicated indifference.
Statistical analysis
Experiments were conducted at least in triplicate. Data were recorded as mean ± standard deviation. One-way analysis of variance was performed by SPSS 26 (SPSS Inc., USA). Statistical significance was considered at p < 0.05.
Results
Antibacterial effects of baicalin
The results of the in vitro drug sensitivity test of baicalin are shown in Figure 1. As shown in Figure 1A, 2 and 4 mg/mL baicalin notably reduced (p < 0.05) the number of single colonies. Then, when the concentrations of added baicalin were over 6 mg/mL, there were almost no obvious colonies on the plates (Fig. 1A d–f). Likewise, as shown in Figure 1B, compared with the control group (average single colony of S. maltophilia was 947), baicalin treatment could obviously (p < 0.05) inhibit the growth of S. maltophilia, and this effect on S. maltophilia was also concentration dependent (Fig. 1B b–f). In comparison, baicalin with identical concentrations exhibited greater inhibitory effects against MRSA than S. maltophilia. After verification of the antibacterial activity of baicalin in vitro, we finally selected 6 mg/mL of baicalin for subsequent related experiments.
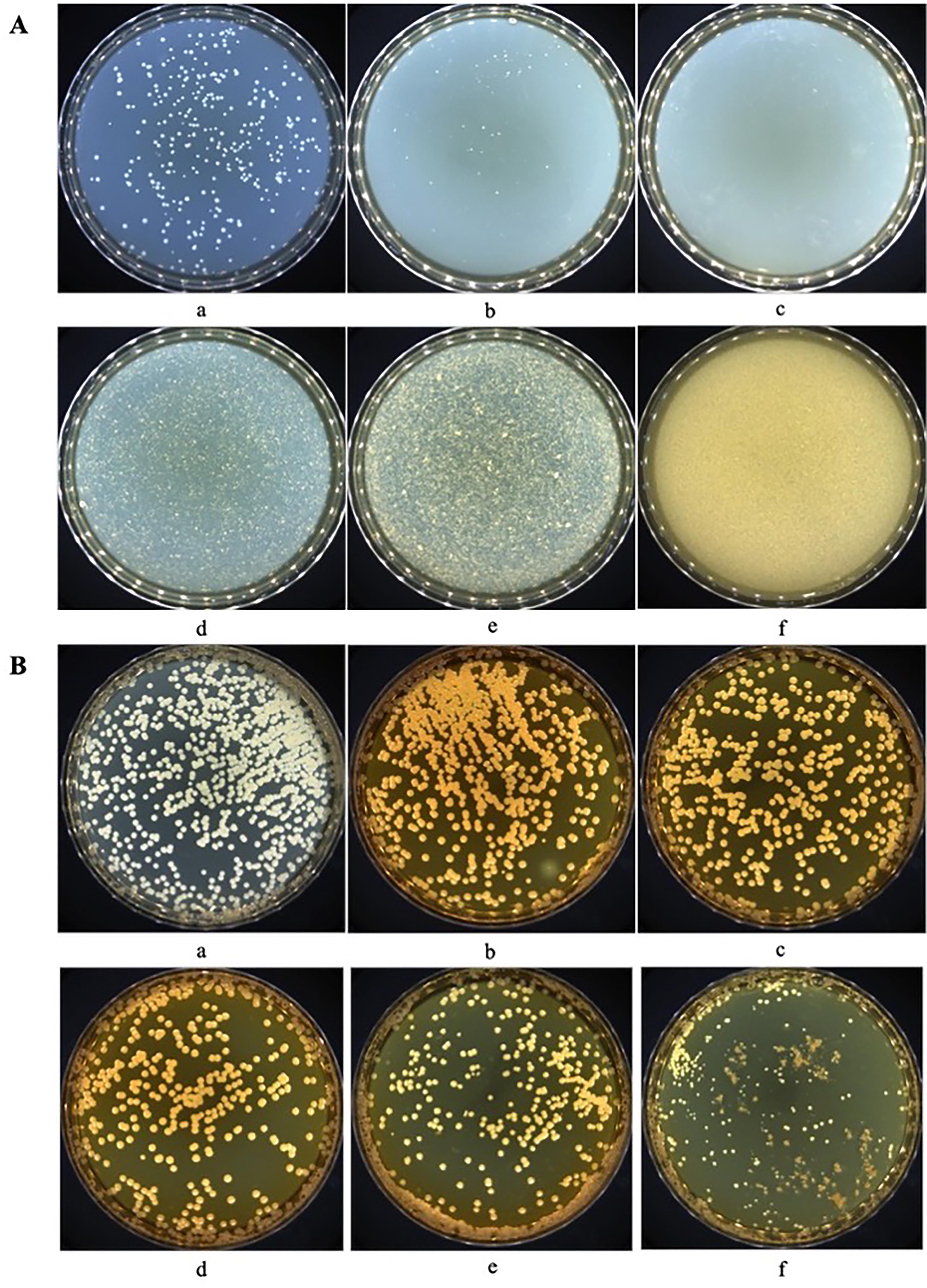
Antibacterial effects of baicalin.
Antibiofilm activity of baicalin on bacteria
The impacts of baicalin on the biofilm of MRSA and S. maltophilia were quantitatively analyzed by a crystal violet staining assay (Fig. 2). For already formed biofilm (Fig. 2A, C), except for the treatment group of 1 mg/mL baicalin against S. maltophilia, all other treatments showed a considerable reducing effect in a dose-dependent manner, as reflected in evident distinctions (p < 0.05) compared with the control. Among them, the inhibition rate of 6 mg/mL baicalin was 52.2% and 19.1%, respectively. Baicalin showed better inhibition on the pre-developed biofilm of MRSA. In addition, there was also a concentration-dependent reduction (p < 0.05) in the biofilm-forming ability of MRSA and S. maltophilia after baicalin treatment (Fig. 2B, D), and the inhibition rate of 6 mg/mL baicalin was 41.7% and 95.2%, respectively. Baicalin exhibited better inhibitory effects on the biofilm formation of S. maltophilia. In summary, baicalin showed effective anti-biofilm activities both on MRSA and S. maltophilia.
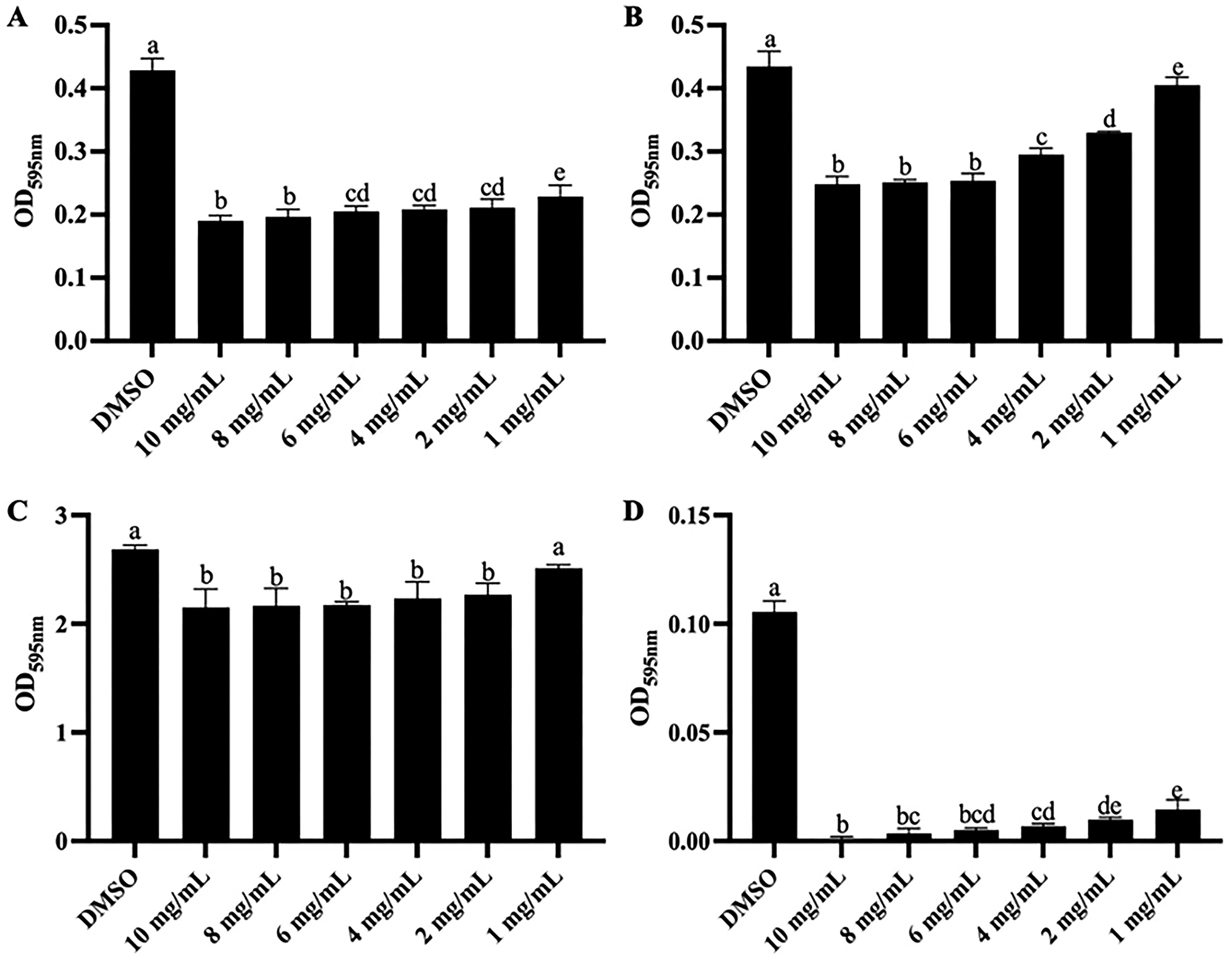
The impacts of baicalin on bacterial biofilm.
Morphology characterization of bacteria
The impacts of baicalin on the ultrastructure of MRSA and S. maltophilia were visualized (Fig. 3). Initially, SEM analysis revealed that MRSA without treatment maintained their characteristic spherical shape with an intact membrane and smooth surface (Rose-Martel et al., 2017), and S. maltophilia displayed a typical rod shape with a complete structure (ElBaradei and Yakout, 2022). In contrast, baicalin treatment resulted in irregular morphology, increased surface roughness, and structural damage in the bacteria. Likewise, further TEM observations revealed destruction of the cell wall, cytoplasmic leakage, and cell lysis. Overall, baicalin treatment could severely compromise the structural integrity of MRSA and S. maltophilia.

SEM and TEM images of bacteria.
Bacterial membrane permeability influenced by baicalin
Live/dead cell staining
The LIVE/DEAD BacLight Bacterial Viability Kit was applied to intuitively reveal the altered cell membrane permeability of bacteria by baicalin (Fig. 4). SYTO-9 can freely penetrate bacteria (both intact and destroyed), making them emit green fluorescence, while PI can only pass through bacteria with ruptured cell membranes, emitting red fluorescence. At the same time, when combined, PI can mask the green fluorescence of SYTO-9, highlighting damaged or dead cells in red.

Investigation of baicalin’s impact on bacterial membrane permeability of bacteria by LSCM.
In Figure 4, the untreated bacteria with intact cell membranes were almost green, while after baicalin treatment, some bacteria showed red fluorescence, and with the extension of baicalin treatment time, red fluorescence intensity obviously increased, and green fluorescence accordingly decreased. The findings indicated that baicalin impacted bacterial membrane permeability in a time-dependent manner.
Release of intracellular components
Measuring extracellular K+ concentration and DNA concentration was also an effective approach to assess bacterial membrane permeability. As shown in Supplementary Figure S1A and 1C, compared with the control group, baicalin treatment markedly increased (p < 0.05) the contents of K+ by 0.58 and 0.88 times in the external environment, respectively. Likewise, in Supplementary Figure S1B and 1D, compared with the control group, baicalin treatment notably increased (p < 0.05) the extracellular concentrations of DNA by 105.38 and 25.14 times, respectively. All these results confirmed that baicalin treatment could obviously increase the cell membrane permeability of bacteria.
Oxidative stress of bacteria induced by baicalin
Cellular ROS levels following baicalin treatment were measured, with findings presented in Figure 5. Relative to the control group, 6 mg/mL baicalin markedly stimulated (p < 0.05) ROS production in bacteria, resulting in fluorescence intensities that were 1.38 and 0.76 times higher, respectively. Previous studies have reported that ROS could interact directly with different macromolecules to induce oxidative stress (Gurrieri et al., 2020). Therefore, it could be deduced that oxidative stress would be reduced in bacteria after baicalin treatment.
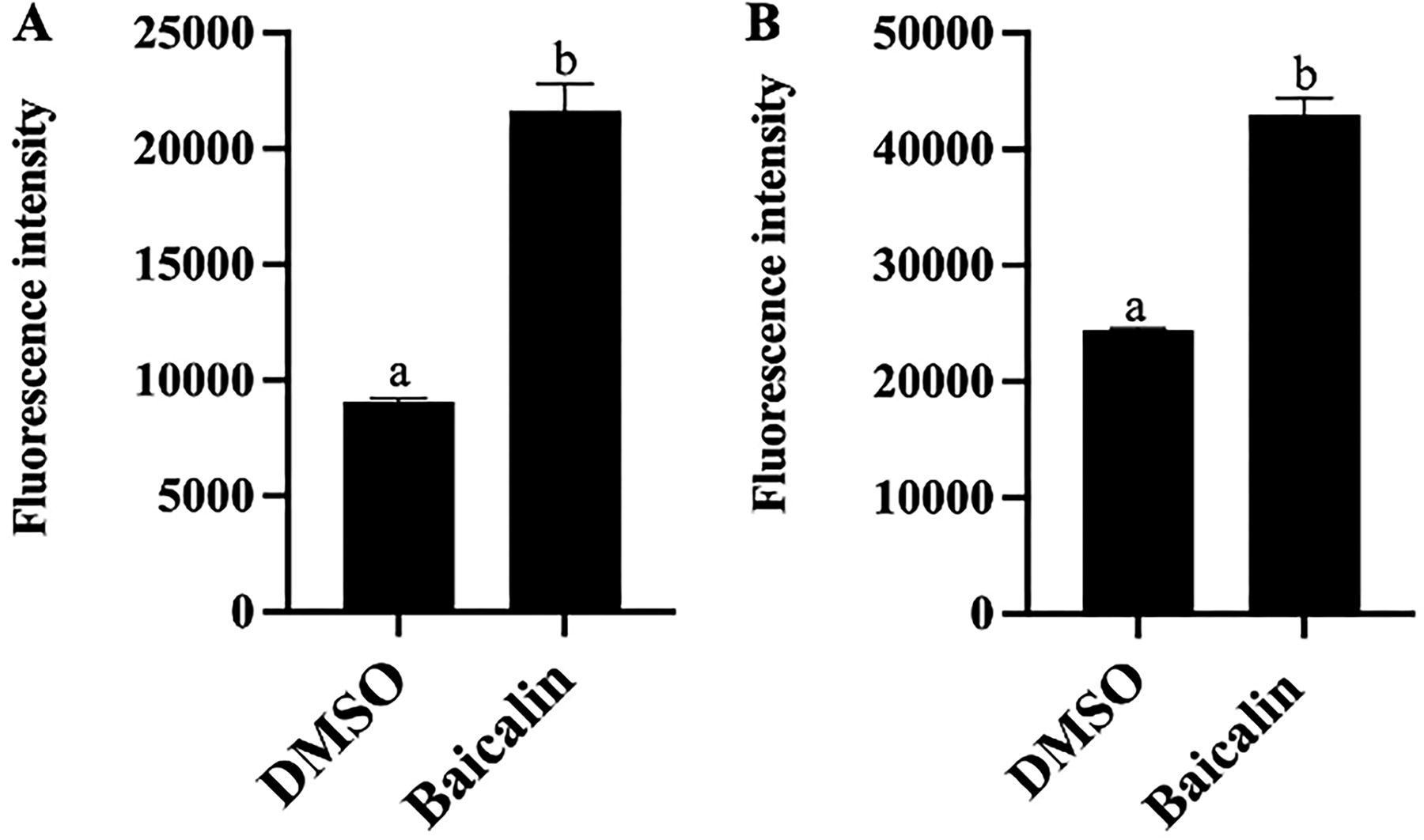
Contents of ROS in bacteria after baicalin treatment.
Transcriptome profiling after baicalin exposure
Overall, we obtained 147.7 and 168.9 million clean reads for MRSA and S. maltophilia, respectively, with mapping rates of at least 92.73% and 62.25% to their reference genomes (Supplementary Table S2). There were good biological reproducibility and clear distinctions in our data (Supplementary Figs. S2 and Figs. S3). Gene expression counts (FPKM > 1) revealed 2721 and 4004 genes expressed in MRSA and S. maltophilia, respectively. Among these, baicalin treatment resulted in 268 differentially expressed genes (DEGs) in MRSA, with 163 markedly upregulated and 105 downregulated (p < 0.05) (Fig. 6A and Supplementary Fig. S4A). In S. maltophilia, 329 DEGs were observed, with 162 obviously upregulated and 167 downregulated (p < 0.05) (Fig. 6C and Supplementary Fig. S4B).
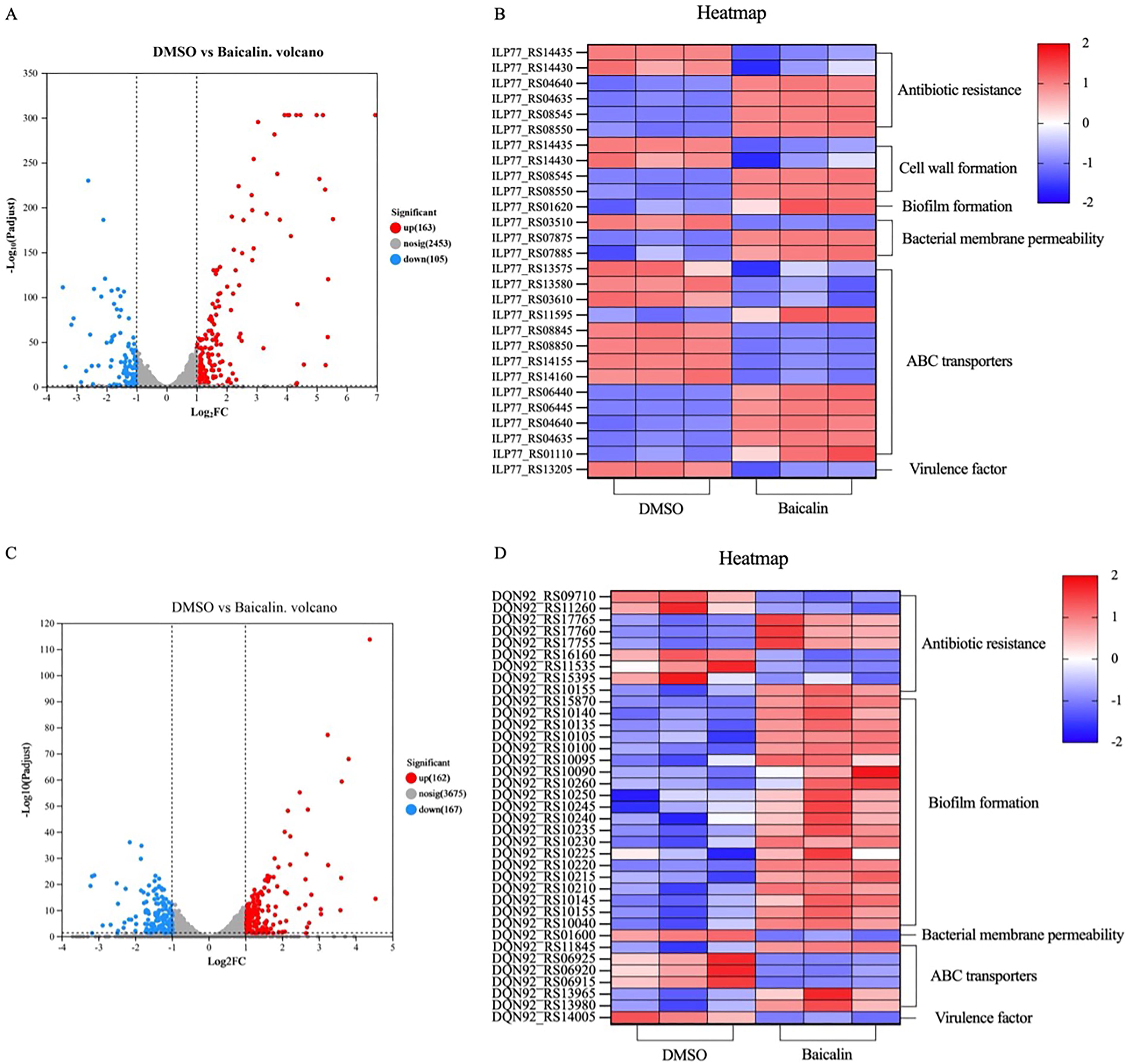
Transcriptomic profiling of MRSA and Stenotrophomonas maltophilia following baicalin exposure.
To deepen our understanding, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichments were conducted using the DEGs as a basis (Supplementary Figs. S5 and Figs. S6). For MRSA, 268 DEGs were associated with 45 GO terms, and for S. maltophilia, 329 DEGs corresponded to 78 GO terms. The top 20 enriched GO terms are shown in Supplementary Figure S5. In KEGG pathway analysis, MRSA-associated DEGs were linked to 86 pathways. Notably, the nitrogen metabolism pathway (map00910) showed the most obvious variation. For S. maltophilia, DEGs were linked to 99 pathways, with flagellar assembly (map02040) and valine, leucine, and isoleucine degradation (map00280) showing notable distinctions. The top 20 enriched KEGG pathways are shown in Supplementary Figure S6.
Specifically, in MRSA (Fig. 6B), among 268 DEGs, key genes were involved in pathways like antibiotic resistance (6 DEGs), cell wall formation (4 DEGs), biofilm formation (1 DEG), bacterial membrane permeability (3 DEGs), ABC transporters (13 DEGs), and virulence factor (1 DEG). In S. maltophilia, of 329 DEGs, significant pathways included antibiotic resistance (9 DEGs), biofilm formation (20 DEGs), bacterial membrane permeability (1 DEG), ABC transporters (6 DEGs), and virulence factor (1 DEG), as shown in Figure 6D.
Finally, qRT-PCR results were consistent with RNA-seq trends, confirming the reliability of the sequencing and validating the transcriptional profiling (Supplementary Fig. S7).
Decrease in bacterial ampicillin resistance
When administered independently, the MICs for baicalin and ampicillin against MRSA were 512 and 4 μg/mL, respectively, and against S. maltophilia were 8192 and 128 μg/mL, respectively. However, their combination markedly lowered the MIC values for both pathogens (Table 1). Specifically, the MIC of baicalin against MRSA decreased to 16 μg/mL and that of ampicillin to 0.5 μg/mL. For S. maltophilia, the MICs were reduced to 1024 μg/mL for baicalin and 16 μg/mL for ampicillin. Additionally, the FICIs for the combination against these two bacteria were less than 0.5, demonstrating a synergistic antibacterial effect. These findings indicated that baicalin could effectively enhance the efficacy of β-lactam antibiotics like ampicillin against these bacteria.
Combined Actions of Baicalin and Ampicillin Against MRSA and Stenotrophomonas maltophilia
MIC, minimum inhibition concentration; MRSA, methicillin-resistant Staphylococcus aureus.
Discussion
Interest in developing new plant-sourced substances with strong antibacterial properties has increased over the years. These natural compounds are being studied as potential alternatives or complements to conventional antibiotics. This research delved into the first-ever exploration of the antibacterial effects and potential mechanisms of baicalin against MRSA and S. maltophilia and predominantly validated its anti-biofilm activity and ability to reduce antibiotic resistance.
Primarily, the antibacterial activity of baicalin against MRSA or S. maltophilia was proved by coating a suitable concentration of bacteria on a series of baicalin-containing agar plates. In contrast, the poured-plate method and growth curve were not suitable for antibacterial experiments in our studies due to the solubility and color interference of baicalin, respectively. The growth of MRSA on agar plates was completely inhibited when baicalin was added at 6 mg/mL, while there were still S. maltophilia colonies at the same concentration, indicating that baicalin had a better antibacterial effect on MRSA (Fig. 1). Meanwhile, in transcriptome analysis, the expression of the eta (ILP77_RS13205) gene in the S. aureus pathway (map05150) was significantly suppressed (p < 0.05) after baicalin treatment. This gene for the virulence factors was present only in MRSA, and its reduction indicated the removal of MRSA toxicity (Changchien et al., 2011). The expression of the pagA (DQN92_RS14005) gene in S. maltophilia was also significantly suppressed (p < 0.05), which contributed to its virulence (Abirami et al., 2021). Therefore, baicalin treatment not only obviously inhibited the growth of bacteria, but also effectively reduced their virulence.
An intricate relationship might exist between bacterial virulence and antibiotic resistance. For sure, Dahdouh et al. (Dahdouh et al., 2017) proved that the predominance of multidrug-resistant factors in Acinetobacter baumannii was closely associated with enhanced expression of its virulence determinants. Therefore, based on the downregulation of the bacterial virulence-related genes, we further explored the impact of baicalin on bacterial antibiotic resistance. Indeed, findings showed that baicalin treatment demonstrated a reduction in bacterial antibiotic resistance (Table 1).
As we all know, the resistance of bacteria to antibiotics is linked to the resistance genes they possess (Xia et al., 2020). Simultaneously, six key genes of MRSA, associated with antibiotic resistance, in a two-component system (map02020) were obviously affected by baicalin treatment. For one thing, there was a marked decrease (p < 0.05) in the expression of dltB (ILP77_RS14435) and dltC (ILP77_RS14430). The Dlt operon consisted of these two genes and was accountable for the d-alanylation of cell wall teichoic acid. Studies have reported that increased d-alanylation of teichoic acid was closely related to the antibiotic resistance of bacteria, such as resistance to cationic antimicrobial peptides (Jeong et al., 2019; O’Shea et al., 2013). This finding was the same as the results of reduced antibiotic resistance in MRSA. For another, bacitracin efflux system-related genes, such as bceA (ILP77_RS04640) and bceB (ILP77_RS04635), were significantly upregulated (p < 0.05). This system could pump bacitracin, a well-known antibiotic peptide, out of bacteria (Benjdia et al., 2017; Ohki et al., 2003). Similarly, the expression of vraS (ILP77_RS08545) and vraR (ILP77_RS08550) was also significantly upregulated (p < 0.05). Boyle-Vavra et al. (Boyle-Vavra et al., 2006) illustrated that the VraS/VraR two-component system was required for the regulation of antibiotic resistance in MRSA and was a potential target for combination antimicrobial therapy. The overexpression of these four genes illustrated the self-protection of MRSA in the face of environmental stress. For S. maltophilia, there were many significantly changed genes related to antibiotic resistance after baicalin treatment. For example, on the one hand, the expression of some related antibiotic resistance genes, such as eptA (DQN92_RS15395) (Cervoni et al., 2023), rpoN (DQN92_RS10155) (Viducic et al., 2007), parR (DQN92_RS16160) (Lee et al., 2014), and so on, was significantly influenced (p < 0.05). On the other hand, more genes associated with multidrug efflux system were obviously changed (p < 0.05), such as oprM (DQN92_RS17755) (Pan et al., 2016), adeA (DQN92_RS09710) (Terkuran et al., 2022), acrB (DQN92_RS17760) (Hayashi et al., 2016), and so on. In comparison, baicalin caused more severe effects on the antibiotic resistance of S. maltophilia than MRSA, including but not limited to β-lactam antibiotics.
Additionally, the structure of the cell wall and cell membrane (Watkins et al., 2012; Xie et al., 2017), and bacterial membrane permeability (Muntean et al., 2018) both contributed to the antibiotic resistance of bacteria. Thus, this research examined how baicalin affected structure and integrity using various assays. First, our experiments proved that baicalin had significant (p < 0.05) anti-biofilm activities (Fig. 2). Next, electron microscope observation indicated that baicalin treatment compromised the cell wall and cell membrane of bacteria (Fig. 3). Then, the increase in the number of bacteria with red fluorescence reflected the increased bacterial membrane permeability after baicalin treatment (Fig. 4). Moreover, the significant leakage of K+ and DNA (p < 0.05) also reflected the increase of bacterial membrane permeability of bacteria (Supplementary Fig. S1).
Interestingly, all these in vitro changes in bacteria had corresponding gene expression alternations. For MRSA, after baicalin treatment, there was a distinct (p < 0.05) increase in the expression of vraS (ILP77_RS08545) and vraR (ILP77_RS08550). These genes were part of the VraS/VraR two-component system, which governed the expression of numerous genes associated with cell wall synthesis (Boyle-Vavra et al., 2006). The increase of these two genes indicated a reparative response in the cell wall structure of MRSA compromised by baicalin. Meanwhile, the expression of dltB (ILP77_RS14435) and dltC (ILP77_RS14430), which were the important composition of the Dlt operon and were the cause of the d-alanylation of cell wall teichoic acid, an extremely important component in the cell wall, in Gram-positive bacteria, was significantly (p < 0.05) suppressed after baicalin treatment (Asciolla et al., 2020; Brown et al., 2013; O’Shea et al., 2013; Poyart et al., 2001). Thus, decreased expression of them was consistent with baicalin-induced cell wall destruction. However, there were no enriched genes associated with cell wall formation in S. maltophilia. Thus, we could find from Figure 3 that baicalin induced more severe damage to the cell wall of MRSA than S. maltophilia.
Then, ILP77_RS01620 was significantly (p < 0.05) upregulated after baicalin treatment and closely related to mevalonate kinase, which was a peroxisomal enzyme that participated in cholesterol biosynthesis (Cader et al., 2019). Cholesterol was a crucial component in the maintenance of cell membrane construction (Wang et al., 2017). Therefore, its overexpression illustrated the self-protection of MRSA against the destructive effects of baicalin on biofilm. For S. maltophilia, after baicalin treatment, the expression of 19 genes in the flagellar assembly pathway was markedly upregulated (p < 0.05), and the formation of flagella contributed to the production of the bacterial cell membrane (Li et al., 2022). In addition, pilJ (DQN92_RS15870) was also significantly (p < 0.05) upregulated after baicalin treatment, which could encode the subunit of type IV pili to promote biofilm formation through binding to the DNA of the extracellular matrix (Ronish et al., 2022). All these alternations suggested that bacterial biofilm was destroyed after baicalin treatment, which was compatible with the findings in Figures 2 and 3.
In addition, kdpA (ILP77_RS07875) and kdpC (ILP77_RS07885) genes in a two-component system (map02020) of MRSA were significantly (p < 0.05) upregulated after baicalin treatment. These two genes could encode inner membrane proteins, KdpA and KpdC, respectively, and they were the critical part of the Kdp-ATPase, which was encoded by the KdpFABC operon and functioned as an ATP-dependent high-affinity K+ transport system (Altendorf et al., 1998; Laimins et al., 1978; Liu et al., 2020). It was reported that this system was induced under conditions of low K+ concentrations and repressed under conditions of high K+ concentrations (Ballal et al., 2007; Hofreuter et al., 2006; Salvetti et al., 2011). In our studies, after baicalin treatment, the extracellular K+ concentration significantly (p < 0.05) increased, which reflected the lack of K+ in bacteria. Thus, kdpA and kdpC genes were significantly (p < 0.05) expressed to absorb extracellular K+ and maintain bacterial homeostasis. On the contrary, the expression of kdpA (DQN92_RS01600) in S. maltophilia was significantly downregulated (p < 0.05), which exhibited that baicalin caused more severe effects on S. maltophilia in terms of K+ leakage.
Finally, previous studies have illustrated that ABC transporters contributed to bacterial resistance by expelling antibiotics through the cell membrane (Zhi et al., 2017). In this study, there were 13 DEGs and 6 DEGs enriched in the ABC transporter system (map02010) of MRSA and S. maltophilia, respectively, suggesting that ABC transporters might also participate in the process of decreased antibiotic resistance of bacteria.
Conclusion
In this study, we systematically explored the antibacterial effects of baicalin against MRSA and S. maltophilia, elucidating the underlying antibacterial mechanisms. Baicalin effectively hindered bacterial proliferation by disrupting the overall cellular structure, increasing membrane permeability, and inducing significant oxidative stress within the bacteria. Baicalin also had both scavenging and inhibiting effects on the two bacterial biofilms. Transcriptomics analysis revealed that baicalin negatively impacted key bacterial physiological pathways, including the two-component system, ABC transport system, biofilm formation, and antibiotic resistance. Moreover, when used in conjunction with ampicillin, baicalin exhibited a synergistic antibacterial effect, suggesting that baicalin could reduce resistance to β-lactam antibiotics. Conclusively, the antibacterial mechanism of baicalin against the two bacteria is multifaceted and needs to be further studied, and the correlation between the antibiofilm activity of baicalin and its decreasing antibiotic resistance effect is worth exploring in the future. This research not only underscores the therapeutic promise of baicalin but also opens new avenues for future studies aimed at developing novel antibacterial strategies and enhancing the efficacy of existing antibiotic treatments.
Footnotes
Acknowledgment
This work was financially supported by the Natural Science Foundation of Shandong Province (ZR2020MC217) and the Special Foundation for Distinguished Taishan Scholars of Shandong Province (tstp20230630).
Authors’ Contributions
K.C.: Conceptualization, investigation, methodology, writing—original draft. X.L.: Formal analysis, investigation, and writing—review and editing. L.S.: Methodology and writing—review and editing. Y.W.: Conceptualization, methodology, and writing—review and editing. J. Zhang: Data curation, methodology, and writing—review and editing. Y.S.: Data curation, methodology, and writing—review and editing. H.Z.: Data curation, methodology, and writing—review and editing. J.S.: Data curation, formal analysis, and writing—review and editing. J.Y.: Data curation, formal analysis, and writing—review and editing. C.P.: Data curation, formal analysis, and writing—review and editing. J. Zang: Data curation, formal analysis, and writing—review and editing. Q.Y.: Funding acquisition, project administration, and writing—review and editing. D.L.: Data curation, formal analysis, and writing—review and editing. T.B.G.: Conceptualization, resources, supervision, and writing—review and editing. D.G.: Conceptualization, resources, supervision, and writing—review and editing. Z.L.: Conceptualization, resources, supervision, and writing—review and editing.
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Funding Information
This work was financially supported by the Natural Science Foundation of Shandong Province (ZR2020MC217) and the Special Foundation for Distinguished Taishan Scholars of Shandong Province (tstp20230630).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.