
Abstract
The present study evaluated the occurrence, antibiogram profile, and sequence types (STs) of multidrug resistant (MDR) Escherichia coli from freshly laid eggs (n = 480), feed (n = 24), water (n = 24), poultry droppings (n = 24), and hand swab samples (n = 10) collected from 24 deep litter (DL) and caged poultry layer farms (12 per category) across Punjab, India. The overall E. coli contamination rate in DL and cage farms was 32% (95% confidence intervals [CI], 26.6–37.8%) and 16.7% (95% CI, 12.6–21.6%), respectively. The logistic regression analysis revealed that the DL system had higher odds of occurrence (odds ratio [OR]) of extended-spectrum beta-lactamase (ESBL) (2.195, 95% CI, 1.065, 4.522) and ESBL/AmpC coproducers (2.69, 95% CI, 1.122, 6.45) compared to the cage system. Additionally, isolates from the DL were 4.065 (95% CI, 1.477, 11.188) times more tetracycline resistant compared to the latter; however, resistance to amoxyclavulanate (OR, 0.437; 95% CI, 0.209, 0.912), and ampicillin (OR, 0.343; 95% CI, 0.163, 0.720) was lesser in DL system. Notably, around 97.7% and 87.2% of the isolates from the DL and cage system were MDR, with the DL system having 6.439 (95% CI, 1.246, 33.283) times more chances of harboring MDR E. coli. Additionally, among the resistance genes, the DL system demonstrated significantly high presence of blaAmpC (56%), qnrA/B/S (42.3%), and tetA/B (30.6%). Furthermore, multilocus sequence typing of 11 MDR isolates (n = 5, DL, and 6, cage) revealed the presence of 10 STs, of which ST10, ST155, and ST156 were found to be of public health importance. Therefore, the present study highlights the burden of MDR, ESBL, and AmpC-producing E. coli on poultry eggs and farm environment, which could be carried over to human handlers and consumers upon direct contact during handling and processing.
Introduction
Antimicrobial resistance has emerged as a global public health concern and the silent pandemic of the 21st century (Crowther-Gibson et al., 2011; Founou et al., 2021). The contributing factors include the injudicious use of antibiotics in the clinical treatment of humans and animals and applications in food production (Llor and Bjerrum, 2014). The accretion of resistance against different classes of antibiotics has resulted in the emergence of multidrug-resistant (MDR) phenotypes, of which resistance against priority pathogens is of prime importance.
The World Health Organization (WHO) categorized members of Enterobacteriaceae especially extended-spectrum beta-lactamase (ESBL) producing Escherichia coli as a critical pathogen (WHO, 2024), owing to the associated clinical implications and urgent need for new antimicrobials. Due to their clinical significance in Indian healthcare settings, ESBL and AmpC-producing E. coli achieved the status of high-priority pathogens (DBT, 2021). While these bacteria were earlier associated with human hospital settings and institutional care centers, their increasing incidence in food-producing animals and retail meat suggests a possible gene exchange between different reservoirs (Wu et al., 2018).
India is the second largest poultry egg producer worldwide with 4.63% of production from Punjab (FAOSTAT 2021, http://www.fao.org/faostat/en/#data/QC) with the state of Punjab contributing to 4.63% of the nation’s total egg (FAOSTAT, 2021). The layer farms in Punjab practice free range/deep litter (DL) and commercial cage farming. The management practices under these systems differ with respect to exposure with the outer surroundings, veterinary supervision, farm biosecurity (BS), and hygienic practices at farm. While multiple studies have focused on ESBL, AmpC E. coli from poultry cloacal swabs (Brower et al., 2017; Tewari et al., 2019), feces (Mondal et al., 2023; Senthamilselvan et al., 2024; Sharma et al., 2021), and caeca (Sakthikarthikeyan et al., 2024), similar studies on eggs and farm environment are scarce in number. Further, the absence of continuous surveillance of eggs for ESBL/AmpC E. coli, in addition to incompliance with hygiene and BS measures from farm to fork augments the risk of cross-contamination from eggs to clean food items, surfaces, and human handlers with these resistant strains. Therefore, the present study addresses this critical gap and investigates farmed eggs, environmental, and hand swab samples for contamination with ESBL, AmpC E. coli, MDR, molecular characterization, and multilocus sequence typing of resistant isolates in DL and caged poultry layer farms across Punjab, India.
Materials and Methods
Sample collection
Twenty-four poultry layer farms distributed equally between DL and cage system (Cg) from Punjab were randomly selected using convenient and purposive sampling. Twenty freshly laid eggs from each farm (n = 480) were aseptically collected in disposable sampling bags (HiMedia Lab, Mumbai). Around five different feed samples from different locations inside the farm were collected and pooled to form one feed sample representative of one farm. Similarly, water and poultry dropping samples were collected; a total of 72 environmental samples were collected. The hand swab samples from farm handlers were collected after informed consent; however, samples were limited (n = 10) due to the unwillingness of farmers from all the farms. All the samples were brought to the laboratory on ice coolers maintained at 4°C and either processed the same day or kept at 4°C (eggs) or −20°C (environmental samples) for further analysis.
Post sample collection, the farmers were enquired about the management practices, usage of antibiotics, hygiene, and BS measures using a well-structured and informed questionnaire (Supplemental Data S1).
Isolation and identification of E. coli
Each egg was cracked open on the air-sac end using a sterile glass rod to separate and empty the inner content in an aseptic disposable sampling bag. The remaining eggshell was put in another sterile disposable bag containing 20 mL of buffered peptone water and hand-crushed. Around 5 mL each of eggshell wash and inner content were enriched separately in 45 mL brain–heart infusion broth (BHI); however, water (5 mL), feed (12.5 g), and droppings (12.5 g) were enriched in 45 mL, 112.5 mL, and 112.5 mL of BHI broth, respectively. The swab tips along with their diluent were enriched in 9 mL of BHI broth. All the samples were incubated overnight at 37°C.
Following enrichment, a loopful of samples was plated on eosin methylene blue agar and incubated at 37°C for 24–48 h. The round colonies with lustrous metallic sheen were picked, and a single colony was inoculated on BHI agar for purification. The purified colonies were subjected to Gram’s staining, followed by matrix-assisted laser desorption/ionization-time of flight mass spectrophotometry (Bruker Daltonik, Germany) for confirmation of E. coli.
Antimicrobial sensitivity testing of E. coli isolates
The antibiotic sensitivity of E. coli isolates was performed using the Kirby-Bauer disc diffusion method, following the Clinical and Laboratory Standards Institute (CLSI, 2020) guidelines. The sensitivity of isolates was tested against 15 antibiotics; ceftazidime (CAZ, 30 μg), ciprofloxacin (5 μg), levofloxacin (5 μg), amikacin (30 μg), gentamicin (10 μg), tobramycin (10 μg), amoxyclav (20/10 μg), ampicillin (10 μg), meropenem (10 μg), trimethoprim (5 μg), cefazolin (30 μg), cotrimaxazole (25 μg), chloramphenicol (30 μg), and tetracycline (30 μg). E. coli strain ATCC 25923 was used as a quality control strain. The isolates were marked as sensitive, intermediate, and resistant per the CLSI (2020) guidelines; however, those depicting intermediate resistance were considered resistant for the analyses. The isolates exhibiting resistance against >3 antimicrobials belonging to at least two classes were categorized as MDR. Furthermore, the multiple antibiotic resistance index (MAR) was calculated using the given formula (Krumperman 1983).
Here, b = 15.
Detection of ESBL production (modified disc diffusion test)
A sterile MHA plate was swabbed with test culture and dried. Subsequently, cefotaxime (CTX 30 μg), CAZ (30 μg), CTX with clavulanic acid (CEC 30/10 μg), and CAZ with clavulanic acid (CAC 30/10 μg) discs were placed at a distance of 24 mm from each other. All the discs were inoculated with phenylboronic acid (PBA) (400 μg) and 0.1M EDTA (292 μg) and incubated at 37°C for 18–24 h. Following incubation, the difference in the zone of inhibition (ZOI) around CTX, CEC and CAZ, CAC was calculated; a difference ≥5 mm indicated ESBL producer (Poulou et al., 2014).
Detection of AmpC production
A sterile MHA plate was swabbed with cefoxitin-sensitive E. coli ATCC 25923 strain and dried. Subsequently, a cefoxitin disc (FOX) was gently placed on an MHA plate using sterile forceps. Simultaneously, sterile filter discs impregnated with tris-EDTA (AmpC disc) were inoculated with the test culture and placed touching the cefoxitin disc. The plate was incubated at 37°C for 18–24 h. Following incubation, the plate was observed for any indentation/flattening of ZOI around FOX near the inoculated AmpC disc; flattening/indentation indicated AmpC production (Poulou et al., 2014).
Detection of Klebsiella pneumoniae carbapenemase production
A sterile MHA plate was swabbed with the test culture, followed by placement of a plain meropenem disc (MRP 10 μg) and meropenem disc (10 μg) with phenylboronic acid (PBA) (400 μg), and incubation at 37°C for 18–24 h. Following incubation, the difference between the ZOI of MRP and MRP-PBA combination was calculated; a difference ≥5 mm indicated Klebsiella pneumoniae carbapenemase (KPC) production (Poulou et al., 2014).
Detection of metallo-beta-lactamase production
The test culture was swabbed over a sterile MHA plate, followed by the placement of two MRP discs (10 μg). 0.5M EDTA was added to one of the discs, and the plate was incubated overnight at 37°C. The difference in ZOI between the MRP-EDTA combination and MRP alone was calculated; a difference ≥5 mm indicated metallo-beta-lactamase (MBL) producer (Poulou et al., 2014).
Molecular characterization of antibiotic resistance genes in E. coli isolates
The isolation of genomic DNA from confirmed isolates was performed using the Qiagen DNA extraction kit using the manufacturer’s protocol. The single-plex PCR reaction for all antibiotic resistance genes in the test E. coli isolates was performed in Mastercycler Gradient Thermocycler (Eppendorf, Germany) using a predefined set of primers and cycling conditions (Supplemental Data S1). Further DNA templates from the previously identified isolates were used as positive controls: bla TEM (GenBank:OL629040.1), bla SHV (GenBank:OL587692.1), and blaAmp C (GenBank:OL587697.1). Furthermore, the field isolates from the present study were used as positive controls for other antibiotic resistance genes.
Hierarchical clustering of AmpC-producing E. coli
The AmpC-producing E. coli were clustered on the basis of their phenotypic resistance against penicillins, cephalosporins, and sulfamethoxazole-trimethoprim combination using Ward’s linkage and squared Euclidean distance measure.
Multilocus sequence typing of MDR E. coli isolates
A total of 11 MDR, ESBL-producing E. coli isolates from eggs and poultry droppings from DL and cage farms were randomly selected and processed for whole genome sequencing (WGS). Thereby sequences of seven housekeeping genes (HKGs); adk, fumC, icd, purA, gyrB, recA, and mdh were blasted in PubMLST database to deduce the allelic profile and sequence types (STs). The data of whole genome were submitted to GenBank database and accession numbers were granted.
Statistical analysis
The statistical analysis was conducted using IBM SPSS Statistics v.27.0.1 (IBM Corporation, Armonk, NY). The occurrence of E. coli from different sample sources, phenotypic and genotypic AMR profile, and MDR% between different systems of rearing was compared using chi-square statistic, where p < 0.05 was taken as significant. Further, the modeling of resistance profiles of isolates; phenotypic/genotypic/MDR (dependent variable) versus system of rearing (independent variable) was done using binary logistic regression analysis and odds ratio (OR) was calculated and plotted using forest plot.
Results
Occurrence of E. coli
The sample-wise E. coli contamination from different systems of rearing is tabulated in Table 1. The analysis revealed that the E. coli contamination was significantly affected by the system of poultry rearing (p < 0.05), with the eggs from DL being 1.97 (95% CI, 1.289, 3.009) times more contaminated than those in Cg. Additionally, the overall occurrence of E. coli from all samples in DL was higher (OR, 2.125; 95% CI, 1.420, 3.181) than Cg.
Sample-Wise Occurrence of Escherichia coli, ESBL, AmpC, ESBL/AmpC, and MDR E. coli
Different superscripts indicate a significant difference within the same sample type between two different systems of rearing (p < 0.05).
ESBL, extended-spectrum beta-lactamase; MDR, multidrug resistant.
Occurrence of ESBL, AmpC, and ESBL/AmpC coproducing E. coli
DL harbored significantly more ESBL and ESBL/AmpC coproducing E. coli (p < 0.05) (Table 1), while no difference in AmpC E. coli was noted between the systems (p > 0.05). The eggs in DL harbored more ESBL E. coli (OR, 2.429; 95% CI, 1.135, 5.196) and coproducers (OR, 3.449; 95% CI, 1.1291, 9.211) compared to those in Cg. Furthermore, the overall occurrence of ESBL E. coli and ESBL/AmpC coproducers was 2.195 (95% CI, 1.065, 4.522) and 2.69 (95% CI, 1.122, 6.45) times more in the DL system than Cg (Fig. 1).
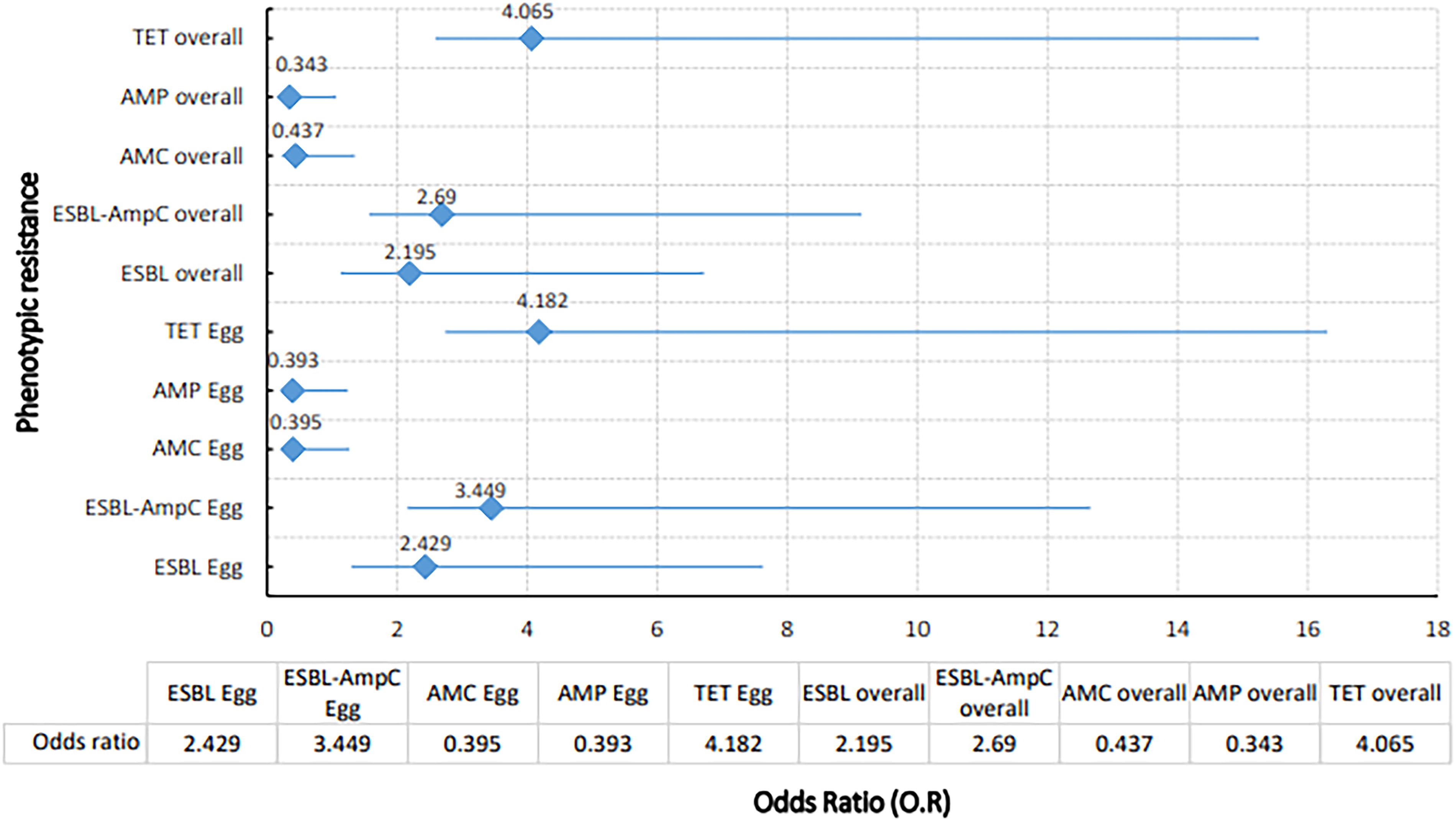
Results from logistic regression modeling the risk of occurrence of phenotypic resistance (egg and overall) against poultry farming system, with cage system kept as the reference category. The X-axis represents the odds ratio on an exponential scale. The horizontal lines represent 95% confidence intervals (CI) for the odds ratio. AMC, amoxyclavulanate; AMP, ampicillin; ESBL, extended-spectrum beta-lactamase-producing Escherichia coli; TET, tetracycline.
Phenotypic antibiogram profile of E. coli isolates
The isolates from eggs in DL exhibited maximum resistance against tetracycline (92%), cefazolin (76%), and CAZ (68%). Likewise, in Cg, egg isolates were majorly resistant to tetracycline (73.3%), cefazolin (66.6%), and trimethoprim (66.6%) (Table 2). However, the overall tetracycline resistance was 4.065 (95 CI, 1.477, 11.188) times more in DL. Unlikely, the odds of resistance against amoxyclavulanate (OR, 0.437; 95% CI, 0.209, 0.912) and ampicillin (OR, 0.343; 95% CI, 0.163, 0.720) were still low in DL (Fig. 1).
Phenotypic Antibiogram Profile of Escherichia coli Isolates Recovered from Different Systems of Rearing
Different superscripts indicate a significant difference in resistance % in the same sample type between two different rearing systems (p < 0.05).
Different superscripts indicate a significant difference in overall resistance % against the same antibiotic between two different rearing systems (p < 0.05).
AK, amikacin; AMC, amoxyclavulanate; AMP, ampicillin; C, chloramphenicol; CIAs, critically important antimicrobials, antibiotics; CAZ, ceftazidime; COT, cotrimoxazole; CTR, ceftriaxone; CTX, cefotaxime; CIP, ciprofloxacin; CZ, cefazolin; GEN, gentamicin; LE, levofloxacin; MRP, meropenem; TE, tetracycline; TOB, tobramycin; TR, trimethoprim.
Occurrence of KPC and MBL
The meropenem-resistant isolates from DL (13.3%; 10/75) and Cg (22.2%; 10/45) were subjected to KPC and MBL production tests. However, none of the isolates tested positive for KPC and/or MBL production.
MDR and MAR
MDR% varied significantly among the systems (p < 0.05), with DL harboring 6.439 (95% CI, 1.246, 33.283) times more MDR E. coli compared to Cg (Table 1). Further, majority of isolates from DL (97.7%; 88/90) and Cg (87.2%; 41/47) had MAR > 0.2, implying a high-risk source of contamination (Supplemental Data S1).
Hierarchical clustering of AmpC-producing E. coli
The dendogram analysis of AmpC producers revealed the formation of seven major clusters, based on the predominance of resistance against the following antimicrobials; cluster 1, CTR, CTX, and TR; cluster 2, CTR, AMP, and CZ; cluster3, CTX; cluster 4, CAZ; cluster 5, CTR, CAZ, CZ, and TR; cluster 6, TR; and cluster 7, AMC; respectively (Fig. 2).

Hierarchical clustering of AmpC-producing Escherichia coli based on phenotypic resistance against CTR, CAZ, CTX, CZ, AMP, AMC, and TR, using Ward’s method and squared Euclidean distance measure. AMC, amoxyclavulanate; AMP, ampicillin; CAZ, ceftazidime; CTR, ceftriaxone; CTX, cefotaxime; CZ, cefazolin; TR, trimethoprim.
Molecular characterization of antibiotic resistance genes in E. coli isolates
The study revealed a significantly higher presence of bla Amp C, qnrA/B/S, tetA/B, and aac(3)-IIa and aph(3)-Ia genes in DL compared to Cg (p < 0.05) (Table 3). This was evident by higher odds of occurrence of bla Amp C (5.046; 95% CI, 2.186, 11.65), qnrA/B/S (2.96; 95% CI, 1.313, 6.673), tetA/B (6.973; 95% CI, 1.997, 24.344), and aac(3)-IIa and aph(3)-Ia (2.776; 95% CI, 1.052, 7.326) in isolates from DL (Fig. 3). Furthermore, isolates carrying multiple antibiotic-resistant genes were also reported in the study (Supplemental Data S1).

Results from logistic regression modeling the risk of occurrence of resistance genes (egg and overall) versus poultry farming system, with cage system kept as the reference category. The X-axis represents the odds ratio on an exponential scale. The horizontal lines represent 95% confidence intervals (CI) for the odds ratio.
Molecular Characterization of Escherichia coli Isolates from Different Systems of Poultry Rearing
Different superscript in the same sample source and AMR genes between different systems of rearing indicate a significant difference (p < 0.05).
Multilocus sequence typing of E. coli isolates
The present study reported ten distinct STs, viz., ST155, ST10, ST156, ST942, ST165, ST295, ST1286, ST2544, ST4162, and ST90 (Table 4). No clonal relationships were observed between the farms, and no new ST was reported. However, among the reported STs, ST10, 155, and 156 were found to be of public health importance, given their wide host spectrum and association with resistant gene determinants such as mcr-1 and blaNDM . in previous studies.
Multilocus Sequence Typing (MLST) Profile of MDR Escherichia coli Isolates
AK, amikacin; AMC, amoxyclavulanate; AMP, ampicillin; C, chloramphenicol; COT, co-trimoxazole; CTR, ceftriaxone; CIP, ciprofloxacin; CZ, cefazolin; GEN, gentamicin; LE, levofloxacin; MRP, meropenem; TE, tetracycline; TOB, tobramycin; TR, trimethoprim.
Discussion
The contamination of farmed eggs varies depending on the poultry rearing conditions and management practices, particularly the BS measures implemented on farms. These measures are designed to prevent the introduction of external infections (external BS) and to limit the spread of pathogens within the farm (internal BS). Our study found significant differences between DL and caged (Cg) farms in terms of rearing practices. The majority of DL farms (58.3%) lacked on-farm BS measures (both external and internal). Additionally, the birds had unrestricted access to the outdoors, which led to eggs being laid near farm dust, dirt, feathers, and bird droppings—potential sources of bacterial contamination. Moreover, none of the farms cleaned either visibly clean or dirty eggs after laying, allowing any contamination on or within the eggs to persist from the farm to the consumer. In contrast, all Cg farms adhered to strict BS protocols (both external and internal) and housed birds in clean cages, with physical separation of eggs from the birds (i.e., cloaca, feathers, and droppings). Eggs were laid directly onto clean trays, minimizing contamination risks. These factors contributed to a higher presence of E. coli on eggs from DL farms compared to Cg systems. This observation aligns with findings from studies by De Reu et al. (2008) and Fahim et al. (2021), which reported higher bacterial loads on eggs from DL compared to Cg system.
Animal food products are subjected to risk of contamination with resistant bacteria and resistance genes at nearly all the stages of processing (Cabello, 2006; Dierikx et al., 2013) and pose a public health threat upon cross-contamination to clean surfaces, foods, and handlers (Cabello, 2006; Perreten et al., 2005). AMR in Gram-negative bacteria (GNB) like E. coli is largely driven by the production of ESBLs, AmpC enzymes, and carbapenemases. Beta-lactam resistance is a leading cause of treatment failure in resistant GNB infections (Laube et al., 2013; Rawat and Nair, 2010), necessitating the use of last resort antibiotics, viz., colistin, and carbapenems, thus promoting resistance to these drugs as well (Cantón et al., 2012). Our study observed that the occurrence of ESBL and ESBL/AmpC coproducers was 2.195 and 2.69 folds higher in eggs from DL. These findings corroborated with previous studies on poultry fecal and cloacal swab samples from India (Brower et al., 2017; Mondal et al., 2023; Senthamilselvan et al., 2024); however, our reported occurrence was higher. Further, meropenem resistance (DL, 13.3%; Cg, 22.2%), and AmpC production (DL, 45.3%; Cg, 40%) were high in both systems, however, the variations were insignificant (p > 0.05). Our results indicate toward misuse of beta-lactams, especially third-generation cephalosporins in both systems, which could be higher in DL than Cg. This was supported by the fact that 58.3% of DL farms report self-prescribing antibiotics. Further free access of birds to outer surroundings in DL predisposes eggs to environmental ESBL and ESBL/AmpC coproducers as opposed to the Cg system.
Besides ESBLs, high resistance against tetracycline, fluoroquinolone, penicillin, sulfa drug, and aminoglycoside groups was evident in both systems; however, significant variation was noted only for tetracycline and penicillins. The resistance levels were lower compared to hatchery eggs, which showed 100% tetracycline and 93.47% ampicillin resistance, as reported by Sharan et al. (2023). We sought to establish a link between the reported on-farm antimicrobial usage (AMU) and the resistance profiles across farms. DL farms reported using cephalexin (83.3%), neomycin (75%), fluoroquinolones (66.6%), tetracycline (66.6%), and sulfonamides (66.6%). In contrast, Cg farms used cephalexin (16.6%) and fluoroquinolones (41.6%). However, the reported AMU does not fully explain the differences in resistance profiles between the two systems. Notably, none of the farms reported using third-gen. cephalosporins, carbapenems, or amphenicols, yet resistance to these drug classes was still high.
Additionally, the MDR status of both systems was fairly high: 97.7%, DL; 87.2%, Cg, with 100% MDR from poultry droppings in both systems. Since all farms used droppings as a soil amendment on agricultural farms, the risk of transmission of these MDR E. coli to the environment was high. Additionally, the human handlers and consumers were at an escalated risk of counteracting ESBL, ESBL/AmpC, and MDR E. coli following improper handling, processing, and consumption of contaminated eggs. Interestingly, despite high overall CAZ resistance (67.7%, DL; 48.9%, Cg), the reported occurrence of CTX-M-15 gene was observed to be low (2.6%, DL; 2.2%, Cg). This could be due to the presence of another CTX-M group 1 variant, CTX-M 27, which is also responsible for CAZ hydrolysis (Castanheira et al., 2021).
The multilocus sequence typing (MLST) analysis of MDR isolates from both systems revealed genetic diversity, supported by distinct STs, with no clonal relationships. However, STs from poultry droppings in DL, and Cg farms shared the same clonal complex, that is, ST155. ST155 was previously reported from live bird markets, farm environments, and handlers in Nigeria (Aworh et al., 2019); as a clonal ST in cattle and pigs with MBL determinants (Tewari et al., 2019) in India; food animals across Europe (Ewers et al., 2011); and humans (Matamoros et al., 2017) worldwide. Furthermore, ST10 and ST155 were associated with colistin-resistant E. coli in chicken fecal samples from Lebanon (Al-Mir et al., 2021). Further, we found ST156 from Cg egg, which has been associated with bla NDM-1 and bla CTX-M-15 E. coli in humans and poultry (Giufre et al., 2012; Mushtaq, et al., 2011), bla NDM-5 in layer feces (Tang et al., 2019) and mcr-1 and bla NDM-5 in Muscovy duck (Yang et al., 2016). The presence of such STs from egg and farm environment in our study emphasizes on need of implementation of farm BS, continuous surveillance of eggs, and advisory over safe handling/processing of eggs to break the chain of transmission.
Conclusion
The present study reported high MDR, ESBL, and ESBL/AmpC-producing E. coli from DL system, in addition to high tetracycline resistance. Additionally, DL system was predominated by bla AmpC, tetA/B, qnrA/B/S, and aac(3)-IIa -aph(3)-Ia genes. Further AmpC E. coli clustered together based on resistance against cephalosporins, penicillins, and sulfa drugs. The MLST profile of MDR E. coli isolates marked the presence of STs 10, 155, and 156, which were associated with colistin and metallo-beta-lactamase determinants in previous studies. Therefore, the present study addresses the potential of eggs and farm environment as vehicles of MDR to human handlers upon cross-contamination at farm and household, and therefore highlights the need for implementation of strict BS and hygienic measures while production, collection and handling of eggs.
Footnotes
Acknowledgment
The authors are thankful to Guru Angad Dev Veterinary and Animal Sciences University, Ludhiana, for providing necessary support throughout the course of study.
Authors’ Contributions
Conceptualization: S.T., R.S., J.S.B., and A.K.A. Methodology: S.T., R.S., J.S.B., A.K.A., R.S.A., and J.S. Experimentation: S.T. Writing—original draft preparation: S.T. Writing—reviewing and editing: R.S. and J.S.B. Sampling: S.T. and J.S. Statistical analysis: S.T. Supervision: R.S. and A.K.A. Project administration: A.K.A.
Disclosure Statement
The authors declare no conflict of interest.
Funding Information
The research was conducted and funded by the Indian Council of Agricultural Research, Niche Area of Excellence project on “Antibiotic Resistance: Animal Human Interface” [ICAR/Edn.10 (8)/2016-EP&HS]. S.T. was supported by the University Grants Commission (UGC)-National Talent Agency (NTA-JRF).
Supplementary Material
Supplemental Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.