
Abstract
Background:
Parasites of Entamoeba and Cryptosporidium genera, prevalent among various vertebrates such as humans and pigs, pose a zoonotic threat as common protozoan pathogens. This study investigated the prevalence and genetic diversity of Entamoeba and Cryptosporidium species in pigs and wild boars across central and southern Vietnam, to ascertain parasite transmission dynamics.
Methods:
A total of 113 independent stool samples from 77 pigs and 36 wild boars were analyzed using PCR-based molecular methodologies to detect the presence of Entamoeba spp. and Cryptosporidium spp. The identified species were further characterized through Sanger sequencing, and phylogenetic relationships were analyzed.
Results:
The study revealed a high prevalence of Entamoeba spp. (62%, n = 70/113) and Cryptosporidium spp. (31%, n = 35/113). Entamoeba suis (57%, n = 40) was predominant, followed by Entamoeba polecki (40%, n = 40) and Entamoeba hartmanni (3%, n = 2). Among Cryptosporidium species, Cryptosporidium scrofarum (89%, n = 31) was the most common, followed by Cryptosporidium suis (11%, n = 4). Wild boars exhibited a higher prevalence of Entamoeba infection compared with domestic pigs (p = 0.019).
Conclusions:
The study highlights a high prevalence of Entamoeba and Cryptosporidium, suggesting a potential for zoonotic transmission in Vietnam. Further investigations are necessary to determine the extent to which these parasites in pigs and wild boars contribute to the burden in the human population.
Introduction
Zoonotic diseases have gained global attention, accounting for 60% of emerging infectious diseases (Jones et al., 2008). Viral and bacterial pathogens are more frequently studied, but zoonotic parasites are often neglected. Swine population are the reservoirs for several emerging zoonotic parasites that pose a potential threat to human health (VanderWaal and Deen, 2018). In addition, the close interaction between humans and pigs can result in zoonotic infections, as humans share a significant proportion of parasites with pig populations (Ledger and Mitchell, 2022). Entamoeba spp. and Cryptosporidium spp. are two widespread parasites that cause diarrhea in domestic and wild pigs, with several pathogenic species also occurring in humans (Wang et al., 2022a; Wang et al., 2022b). Vietnam, renowned for its significant pig production and consumption, stands as a potential hotspot for possible zoonotic parasite transmission.
The genus Entamoeba is an intestinal protist that includes several free-living and parasitic species from all classes of vertebrates and some invertebrates. Entamoeba suis and Entamoeba polecki have been identified as the most common causative agents of diarrhea in pigs (Wang et al., 2022b), while Entamoeba histolytica can cause amoebiasis and lead to severe disease in humans. Although experimental observations have shown E. histolytica infections in pigs, no cases have been reported in farmed animals (Girard-Misguich et al., 2011). Of note, E. polecki observed in pigs are occasionally found in humans, suggesting cross-infection of Entamoeba species between the human and animal reservoirs (Stensvold et al., 2018). In addition, a variety of Entamoeba species have been reported in humans (Entamoeba bangladeshi, E. histolytica, Entamoeba dispar, Entamoeba coli, Entamoeba moshkovskii, Entamoeba hartmanni, and E. polecki) (Dos Santos Zanetti et al., 2021), indicating the importance of studying these protozoa in nonhuman reservoirs to understand their transmission dynamics.
Cryptosporidium, on the other hand, belongs to the genus of Apicomplexan protozoa and is an opportunistic parasite that causes self-limiting diarrhea in both humans and a wide range of animals (Chen et al., 2023). Cryptosporidium is commonly found in the intestines of humans and animals and is transmitted by the fecal–oral route. Interestingly, considerable genetic variation in the genus of Cryptosporidium has been reported worldwide with 44 known species and more than 120 identified genotypes (Ryan et al., 2021). Pigs are recognized as a reservoir of Cryptosporidium with 13 different species/genotypes reported thus far. Of note, the majority of cases of cryptosporidiosis in pigs are caused by Cryptosporidium scrofarum (formerly Cryptosporidium pig genotype II) and Cryptosporidium suis (Wang et al., 2021), which occasionally infect humans, suggesting that these two species may be zoonotic and pose a potential risk to humans (Kvac et al., 2009; Moore et al., 2016).
Detection and monitoring of Entamoeba spp. and Cryptosporidium spp. in pigs have been carried out in several Asian countries as these parasites have an impact on human and animal health (Chen et al., 2023; Dos Santos Zanetti et al., 2021; Matsubayashi et al., 2015). In Vietnam, pig farming stands as a pivotal component of the livestock sector, and the increased interaction between humans and pigs amplifies the potential for zoonotic disease transmission (Coker et al., 2011). However, only limited research has delved into protozoan infections in pigs in this region, particularly on Entamoeba spp. Consequently, monitoring Entamoeba spp. in pigs is imperative not only for enhancing public health safety but also for elucidating their genetic diversity to understand genotypes/subtypes in circulation. Concurrently, there is mounting interest in understanding the distribution and genetic attributes of Cryptosporidium species, given the documented evidence of Cryptosporidium infections associated with pigs in this region (Iwashita et al., 2021). Additional surveillance studies are warranted to ascertain the presence of zoonotic species such as Cryptosporidium hominis or Cryptosporidium parvum in these animals, thus evaluating the potential risk of pathogen transmission.
This study aims to identify Entamoeba and Cryptosporidium parasites in pigs and wild boars in Vietnam, while also subjecting the specific species/genotypes to molecular analysis through sequencing of the small subunit ribosomal RNA (SSU rRNA) gene to elucidate their genetic diversity.
Materials and Methods
Ethical statement
The study was approved by the Ethics Committee of Hue University of Medicine and Pharmacy, Hue University, Vietnam (H2022/020) and the Animal Ethics Committee of the International University (IU)—Vietnam National University—Ho Chi Minh City (VNUHCM, August 2022).
Study design and sampling
Between April and June 2022, a total of 113 fecal samples were collected from 77 pigs and 36 wild boars from various farms and slaughterhouses in southern and central Vietnam. In central Vietnam, 88 samples (77 pigs and 11 wild boars) were collected from eight different farms and slaughterhouses. In southern Vietnam, 25 stool samples were collected from wild boars from two different farms. Fecal samples were collected from various cages in each farm and slaughterhouse to ensure that the samples originated from independent sources.
DNA isolation and PCR amplification
DNA isolation was performed using the QIAamp Fast DNA Stool Mini Kit (Qiagen GmbH, Hilden, Germany). The manufacturer’s protocol was followed with minor modifications. For partial amplification of the SSU rRNA gene of Entamoeba species, conventional PCR with Taq DNA polymerase (Qiagen GmbH) at an annealing temperature of 64°C was performed. Forward and reverse primers that were utilized were sourced from a prior publication, resulting in a PCR product of 580 bp (Verweij et al., 2001). For amplification of the SSU rRNA gene of Cryptosporidium species, nested PCR was performed with Taq DNA polymerase (Qiagen GmbH) at an annealing temperature of 55°C. The outer and inner primer pair was described in the study by Xiao et al. (2001). The inner PCR product was 830 bp. Amplicons were visualized on 1.5% agarose gels stained with c Green. Positive samples underwent an additional round of PCR screening to ensure reproducibility.
Sanger sequencing
All samples positive for PCR, which produced clear amplicons, underwent Sanger sequencing. PCR products were purified using ExoSAP-IT™ PCR Product Cleanup Reagent (Thermo Fisher Scientific, Carlsbad, CA, USA). Amplicons were sequenced using both forward and reverse primers using the BigDye Terminator V.1.1 Sequencing Kit (Applied Biosystems™, Foster City, CA, USA) and were subsequently purified on Sephadex G50 (Cytiva, Uppsala, Sweden). The product templates were sequenced with the Applied Biosystems 3130xl Genetic Analyzer (Applied Biosystems).
Phylogenetic analysis
The obtained SSU rRNA gene sequences were assembled using Lasergene SeqMan II software (Jin and Sun, 2018) and aligned using EditSeq software (DNASTAR, Madison, Wisconsin, USA). The sequences were analyzed together with the representative nucleotide sequences of Entamoeba and Cryptosporidium species and/or respective genotypes using MAFFT version 7 using the G-INS-i model (Katoh et al., 2019). All reference sequences were obtained using the GenBank database (www.ncbi.nlm.nih.gov/GenBank). Phylogenetic trees were reconstructed using MEGA version 11 (www.megasoftware.net) (Tamura et al., 2021), employing the Maximum Likelihood method and the general time reversible plus gamma distribution model. The statistical robustness and reliability of the branching order were confirmed via bootstrapping with 1000 replicateset al. The resulting phylogenetic tree was annotated and visualized using the online tool iTOL v6 (https://itol.embl.de/) (Letunic and Bork, 2019). The representative sequences of Entamoeba spp. and Cryptosporidium spp. obtained in this study have been deposited in the National Center for Biotechnology Information (NCBI) GenBank database and can be retrieved with accession numbers PP735745–PP735814 (Entamoeba spp., n = 70) and PP748483–PP748517 (Cryptosporidium spp., n = 35).
Data analysis
All analyses were conducted using GraphPad Prism (version 9.5.1). A p-value <0.05 was deemed statistically significant. When comparing differences between and among groups, Fisher’s exact or chi-squared tests were used where appropriate.
Results
Demographic and sample characteristics
This study, conducted in central and southern Vietnam, involved the analysis of a total of 113 fecal samples. In central Vietnam, 88 samples were collected from various pig holdings, comprising adult pigs and wild boars sourced from five slaughterhouses (n = 64), one pig farm (n = 13), and two wild boar farms (n = 11). In southern Vietnam, 25 samples were collected from two wild boar farms, including wild boar piglets sampled between 24 and 160 days after birth (mean: 100 days ±54) and weighing between 4 and 35 kg (mean: 17.2 kg ±30.5).
DNA positivity of Entamoeba spp. and Cryptosporidium spp.
PCR analysis targeting the SSU rRNA gene indicated that 62% (n = 70/113) of the tested samples were positive for Entamoeba spp. Among these, 55% (n = 42/77) were from pigs, and 78% (28/36) were from wild boars. A statistically significant difference in the Entamoeba spp. prevalence was observed between pigs and wild boars (p = 0.019) (Table 1). Regarding Cryptosporidium spp., the prevalence was 31% (n = 35/113). The prevalence was marginally lower in wild boar at 29% (n = 8/36) compared with pigs at 35% (n = 27/77), with no statistical difference observed between the groups (p = 0.5) (Table 1). Coinfection with both parasites was detected in 27% of the samples (n = 30/113).
Distribution of Entamoeba spp. and Cryptosporidium spp. in Domestic Pigs and Wild Boars
CI, confidence interval.
Species identification and phylogenetic analysis
Among the Entamoeba-positive samples, E. suis (n = 40/70) predominated with 57%, followed by E. polecki at 40% (n = 28/70) and E. hartmanni at 4% (n = 2/70). Subtyping of E. polecki isolates in this study revealed subtypes ST1 (n = 14, 50%) and ST3 (n = 14, 50%) (Table 1). In addition, two species of Cryptosporidium were detected, with C. scrofarum being the most prevalent at 89% (n = 31/34), followed by C. suis at 11% (n = 4/35) (Table 1). The distribution of all Entamoeba species was uniform across domestic pigs and wild boar, except for the two E. hartmanni samples, which were exclusively found in pig samples from central Vietnam (Fig. 1). This is also the first study reporting on E. hartmanni in the pig population. Sequences of the two E. hartmanni samples in this study (#PP735749 and #PP735767) share >99% homology with the human isolate (#KX618191). The Entamoeba spp. reliably clustered in a distinct branch with a bootstrap value exceeding 95 (Fig. 1). Similarly, the Cryptosporidium spp. clustered with C. scrofarum and C. suis branches with a bootstrap value exceeding 99. Of note, C. suis was found only in pigs from central Vietnam (Fig. 2), which are phylogenetically related to other pathogenic species (C. parvum and C. hominis), suggesting a potential risk of human infection from C. suis.

Reconstructed phylogenetic tree using Entamoeba spp. SSU rRNA gene nucleotide sequences (n = 70, this study) along with 23 reference nucleotide sequences from 18 distinct Entamoeba species retrieved from GenBank. The sequences from this study are highlighted in red (central VN) and blue (southern VN). Only bootstrap values above 70 were shown. SSU rRNA, small subunit ribosomal RNA; VN, Vietnam.
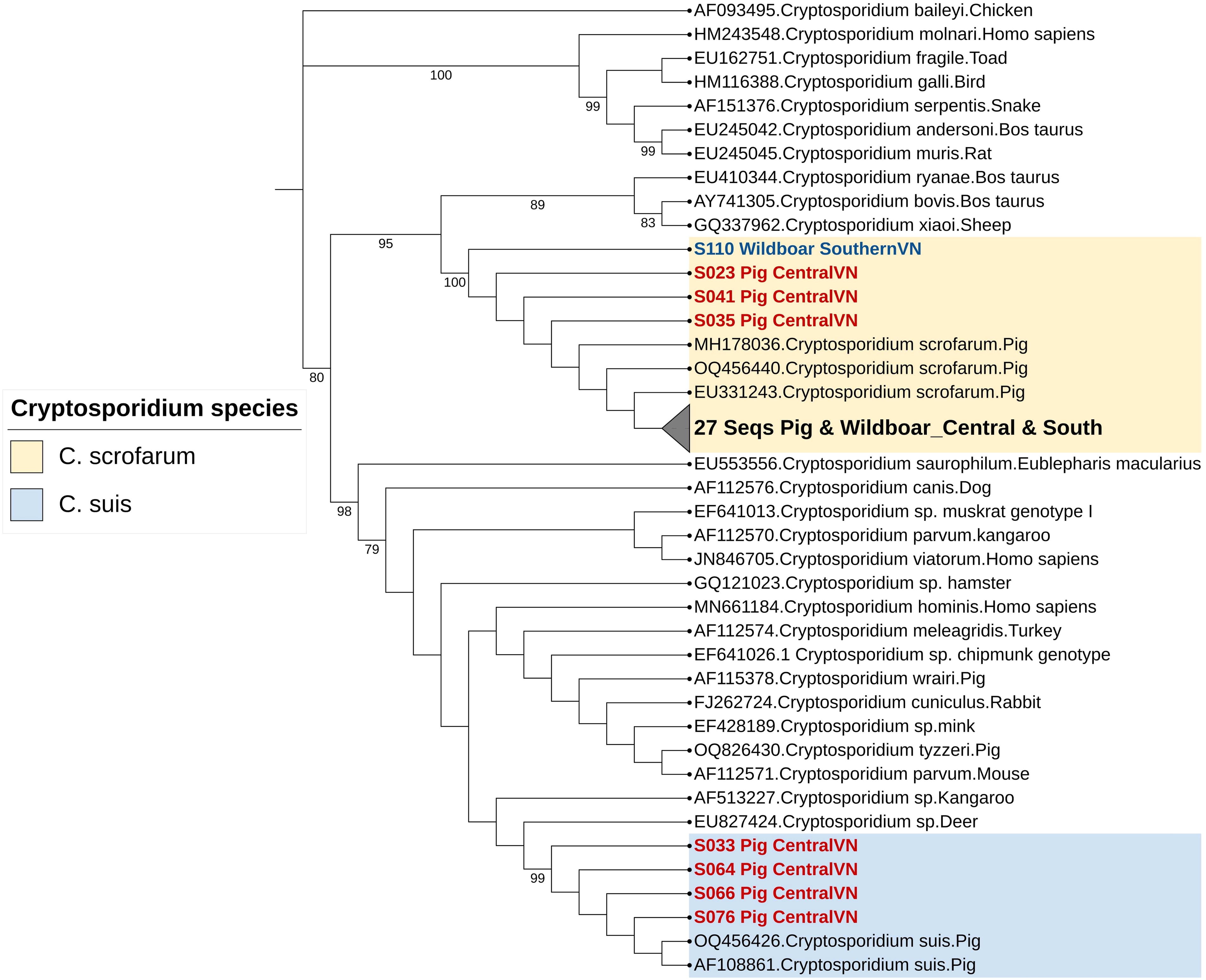
Reconstructed phylogenetic tree using Cryptosporidium spp. SSU rRNA gene nucleotide sequences (n = 35, this study) along with 34 reference sequences from 28 distinct Cryptosporidium spp. retrieved from GenBank. The sequences from this study are highlighted in red (central VN) and blue (southern VN). Only bootstrap values above 70 were shown.
Discussion
This study investigated the prevalence and genetic diversity of Entamoeba and Cryptosporidium species in pigs and wild boars across central and southern Vietnam, offering insights into potential zoonotic risks and parasite transmission dynamics. Notably, this study revealed significant differences in the prevalence of Entamoeba spp. between pigs and wild boars, alongside the exclusive detection of pathogenic Cryptosporidium species in pigs from central Vietnam.
The distribution of Entamoeba and Cryptosporidium in vertebrates varies geographically (Cui et al., 2019). However, the differences in prevalence may stem from varied detection methods. Microscopy, commonly employed, is less sensitive than molecular techniques, as it can misdiagnose parasite morphology due to stool sample artifacts (Stensvold and Nielsen, 2012). This study revealed a high prevalence of Entamoeba spp. at 62%, as observed in Wang’s study in China, where 60% of 1254 pigs were infected (Wang et al., 2022b), with similar rates observed in Nepal at 61% (Adhikari et al., 2021). In our study, determining the age and sex of pigs is a challenge as different pigs live together in the same barn. However, the study samples from pigs were only taken from adult animals. The prevalence of porcine Entamoeba varies depending on the methodology utilized. For instance, microscopy-based studies reported lower prevalence in Korea (4%) (Ismail et al., 2010), Greece (8%) (Symeonidou et al., 2020), and Cambodia (27%) (Schar et al., 2014) compared with molecular screening studies in Indonesia (85%) (Wardhana et al., 2020) and Vietnam (92%) (Jacob et al., 2016), which indicated much higher prevalence rates. Numerous factors, including geographical location, climate, housing conditions, farming practices, feed and water quality, animal density, cohabitation with other livestock, and veterinary care, are likely contributors to such a varying prevalence of parasites in farmed pigs. Entamoeba spp. has been detected across a range of vertebrates, with prevalence rates varying from 0.8% in dogs to around 63% in nonhuman primates and 100% in long-tailed macaques (Cui et al., 2019). Despite its presence in a wide array of vertebrates, studies on the pathogenicity induced by the parasite in animals are limited or rarely documented. This study marks the first reporting of Entamoeba spp. prevalence in the wild boar population in Vietnam, revealing a rate of approximately 78%, notably higher than that documented in an Iranian study (52%) (Yaghoobi et al., 2016). While the prevalence of Entamoeba in wild boar surpasses that in domestic pigs (78% compared with 55%), it is important to note the limitation of our study, which had a small sample size of 36 wild boars.
In this study, the presence of three Entamoeba species was identified as follows: E. suis, E. polecki, and E. hartmanni. Domestic pigs are the primary hosts for E. suis and E. polecki (ST1, ST3) worldwide (Cui et al., 2019), although the distribution of these species and subtypes varies. For instance, Wang et al. showed that E. polecki (82%) was the predominant species, followed by E. suis (18%) (Wang et al., 2022b). Conversely, our findings show that the majority of Entamoeba-positive samples consisted of E. suis (57%) and E. polecki (40%). E. polecki exhibits zoonotic potential and has been detected in several hosts, including humans and pigs. There are four subtypes of E. polecki (ST1–ST4), all of which have been observed in humans. ST1 and ST3 have been identified in pigs, while ST2 has solely been detected in nonhuman primates. Notably, E. suis has not yet been detected in humans (Ji et al., 2019). Interestingly, two E. hartmanni detected from two distinct slaughterhouses in central Vietnam displayed significant sequence homology with human isolates (>99%), suggesting a zoonotic potential in this species. While E. hartmanni is widely distributed among nonhuman primates and humans [21], this study marks the first detection of this species within the pig population. Although E. hartmanni is generally considered nonpathogenic in humans, a recent study from Indonesia reported a significantly higher prevalence of E. hartmanni in loose stools (69%) compared with its prevalence in healthy schoolchildren (31%) aged 7–15 years (Matsumura et al., 2019). This finding suggests a potential role for E. hartmanni in mild diarrhea among children. However, further research is needed to confirm its pathogenicity, as the study by Matsumura et al. did not account for the presence of true pathogens.
Cryptosporidium infections are widespread in pigs, but data on wild boar are limited (Nemejc et al., 2013). The prevalence of Cryptosporidium in domestic pigs varies globally, with rates ranging from 0.4% in Germany to 80% in South Africa (Chen et al., 2023). This study found moderate prevalence in both domestic pigs (35%) and wild boar (29%), surpassing previous findings in central Vietnam (14%) (Nguyen et al., 2013) and China (4.5%) (Wang et al., 2022a). Globally, Cryptosporidium pig infection rates are estimated at 16.3%, with Asia and Africa being a hotspot (Chen et al., 2023). Surveillance studies conducted in Vietnam have revealed varied infection rates, with reports of both C. suis and C. scrofarum species (Iwashita et al., 2021; Nguyen et al., 2013). The rates observed align with the findings of this study. Although no C. suis was observed in the wild boar population in this study, they remain susceptible hosts (Nemejc et al., 2013). Although numerous Cryptosporidium species have been identified in pig populations, over 90% of cases in pigs are attributed to C. scrofarum and C. suis, indicating pigs as the main hosts (Chen et al., 2023). C. scrofarum has been identified as a causative pathogen in patients presenting with diarrhea, including those who are HIV positive (Xiao et al., 2002).
Conclusion
Although our study has limitations regarding sample characteristics, limited sample size, and the need for further confirmation of pathogenicity among animals, the prevalence of Entamoeba and Cryptosporidium species suggests a potential for zoonotic transmission, underscoring the need for additional epidemiological surveillance efforts. In summary, the study highlights a significant prevalence of Entamoeba and Cryptosporidium infections in domestic pigs and farmed wild boars in central and southern Vietnam. The diverse species of Entamoeba identified, including the first detection of E. hartmanni in domestic pigs in Vietnam, underscore the importance of surveillance in animal populations. Further investigations in the human population of these regions are essential to assess whether the parasites found in pigs and wild boars contribute to the human disease burden.
Footnotes
Acknowledgments
The authors thank all study participants for their participation. The authors also thank Ms. Le Thi Kieu Linh for supporting sequencing procedures.
Authors’ Contributions
T.P.V. designed and supervised the study and contributed to the study materials for all experimental investigations. L.C.C., D.M., T.T.G., V.M.T., N.T.M.C., T.N.P.A., L.N.N.H., N.T.T.H., T.N.M., and L.H.S. participated in the study design. L.C.C., T.T.G., V.M.T., and L.N.N.H. collected samples. L.C.C., D.M., N.H., and A.M. performed the experimental procedures. L.C.C. performed statistical and phylogenetic analysis. L.C.C. wrote the first draft, and T.P.V. revised the first draft. All authors have read and approved the article.
Data Availability Statement
All data generated or analyzed during this study are included in this article. A total of 105 successfully sequenced samples were submitted to the NCBI GenBank database, with accession numbers for Entamoeba ranging from PP735745 to PP735814 (n = 70) and for Cryptosporidium ranging from PP748483 to PP748517 (n = 35).
Disclosure Statement
The authors declare no conflicts of interest. The funder has no role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Funding Information
This study was funded by the PAN-ASEAN Coalition for Epidemic and Outbreak Preparedness [PACE-UP; German Academic Exchange Service (DAAD) Project ID: 57592343].