
Abstract
This study aimed to investigate the influence of garlic metabolites on the quorum sensing (QS) of Bacillus cereus, a foodborne pathogen that controls its main virulence factor through QS. The QS signal receptor PlcR of B. cereus was targeted by molecular docking with 82 garlic metabolites to identify the most potent QS inhibitors. Five metabolites, quercetin, kaempferol, luteolin, flavone, and rutin, were selected for further evaluation of their impacts on the growth, toxin production, and virulence of B. cereus in vitro. The expression levels of key QS genes were also measured to verify their anti-QS ability. The results revealed that quercetin reduced enterotoxin production by B. cereus but did not affect the QS process at the transcriptional level; flavone and rutin in garlic interfered with the QS of B. cereus by competing with the autoinducing peptide (AIP) PapR7 for the PlcR binding site, resulting in decreased enterotoxin secretion and hemolysis without altering the bacterial growth. Interestingly, luteolin and kaempferol in garlic acted as AIP analogs and bound to PlcR to stimulate the QS process and virulence. Furthermore, kaempferol, luteolin, flavone, and rutin had distinct or opposite interactions with PapR7 at the Gln237 or Tyr275 residues of PlcR, which determined the suppression or enhancement of the QS process. The findings suggested that flavone and rutin were effective compounds to inhibit the QS process in garlic and could be used as alternative methods to control B. cereus.
Introduction
Bacillus cereus, a foodborne pathogen, is known for causing gastrointestinal distress in humans through various toxins and virulence factors, including enterotoxins (Hbl, Nhe, CytK) and emetic toxin (cereulide). These toxins lead to symptoms like diarrhea and vomiting upon ingestion of contaminated food. The bacterium’s resilience is enhanced by its ability to form adhesive spores and biofilms, enabling it to survive harsh conditions and adhere to diverse surfaces, thus contaminating a wide range of foods and food contact areas (Ciofu et al., 2022; Huang et al., 2020; Lin et al., 2022; Yu et al., 2019).
The use of chemical antibacterials and antibiotics to combat B. cereus is prevalent but raises concerns over the development of resistant strains. An alternative approach involves disrupting the bacterium’s quorum sensing (QS) process, a cell-density-dependent communication mechanism, that controls gene expression and behaviors like sporulation and toxin production (Mukherjee and Bassler, 2019; Rutherford and Bassler, 2012). By disrupting the QS process of B. cereus, its growth, toxin production, and spread can be effectively controlled without directly killing the bacteria (Defoirdt, 2018; Yi et al., 2021).
Several studies have reported that some plant extracts or their active compounds can inhibit the QS process of different bacteria by interfering with the signal–receptor interactions (Ding et al., 2017; Li et al., 2022; Li et al., 2018; Yu et al., 2022). However, most research on the inhibition of bacterial QS by plant extracts has focused on Gram-negative bacteria, which use small molecules such as acyl homoserine lactones or their derivatives as QS signal molecules. In contrast, Gram-positive bacteria, including B. cereus, use strain-specific peptides known as autoinducing peptide (AIP) as communication signal molecules, and their QS process differs significantly from that of Gram-negative bacteria. There are few reports on the interference of plant extracts with the QS process of B. cereus. In a previous study, garlic extract has been proved to have a potent inhibitory effect on the B. cereus QS process (Jin et al., 2021; Jin et al., 2020). It can significantly inhibit the production of bacterial virulence factors and biofilms at subinhibitory concentrations, reducing the viability, spreadability, and virulence properties of B. cereus. Therefore, garlic extract may be a potential source of B. cereus QS inhibitors, but the inhibitory components and mechanisms in garlic extract are still unclear and need further investigation.
Molecular docking technology can visually reveal details of molecular interactions that are inaccessible to in vitro experiments and can rapidly screen bioactive compounds from large databases with millions of molecules (Yu et al., 2023). In this study, we docked 82 garlic metabolites with the B. cereus QS signal receptor PlcR and selected the most promising QS inhibitor candidate. However, molecular docking-based screening methods may suffer from significant false positives and false negatives, so adequate phenotypic experiments are required to further validate the inhibitory effect of candidate compounds on QS (Defoirdt, 2018). Therefore, we tested the effect of those candidate compounds on the growth, toxin production, and virulence of B. cereus in vitro and measured the expression of key QS genes to verify their anti-QS ability at the molecular level. Our study aimed to identify the main components in garlic that interfere with the QS process and may provide new insights and a theoretical basis for developing novel QS inhibitors.
Materials and Methods
Chemicals and strains
Quercetin (97%, CAS 117-39-5), luteolin (≥98%, CAS 491-70-3), kaempferol (97%, CAS 520-18-3), flavone (≥99%, CAS 525-82-6), and rutin (95%, CAS 153-18-4) were purchased from Shanghai Macklin Biochemical Technology Co., Ltd. B. cereus ATCC14579 was kindly provided by Dr. Li Li, South China University of Technology. Other solvents and reagents were of analytical grade.
Construction of garlic metabolites 3D database
A 3D database of 82 garlic-derived metabolites (Supplementary Table S1) was compiled using structures from PubChem (https://pubchem.ncbi.nlm.nih.gov), representing compounds from garlic alcohol extract, water extract, and essential oil (Abe et al., 2020; Jin et al., 2020; Wang et al., 2016).
Virtual screening of potential QS inhibitors
PlcR protein (3U3W.pdb, 2.4 Å resolution) from the PDB (https://www.rcsb.org/) was employed to identify potential binders via structure-based screening using the molecular docking tool QuickVina-W (Hassan et al., 2017). A sufficiently large search box was constructed to encompass the entire PlcR surface, and multiple iterations were run to explore various binding orientations and affinities. The lowest energy garlic metabolite-PlcR complex was analyzed further, with intermolecular bonds visualized by LigPlot+. The autoinducer peptide PapR7 (ADLPFGF) served as the reference.
Detection of enterotoxins
B. cereus enterotoxins Nhe and Hbl were detected using the GLISA Duopath® Cereus Enterotoxins Immunoassay Kit (Merck KGaA), with modifications from a previous report (Jin et al., 2021). The metabolites quercetin, kaempferol, luteolin, flavone, and rutin were prepared in LB broth at a concentration of 103 CFU/mL B. cereus, adjusted to their noninhibitory concentrations (NIC, Supplementary Table S2). The broth was incubated at 37°C, and aliquots of 1 mL were centrifuged at 13,000 × g for 1 min at 30-min intervals. Subsequently, 150 µL of the supernatant was applied to the kit’s sample pad. Samples displaying red lines in both test and control areas after 30 min were recorded as positive for enterotoxin presence. The detection time and B. cereus count were noted at the initial identification of the toxin.
Determination of hemolytic activity
B. cereus (105 CFU/mL) was cultured in NB medium with quercetin, kaempferol, luteolin, flavone, and rutin at 125 μg/mL (≤NIC). The cultures were shaken at 37°C and 160 r/min for 8–24 h. Postcultivation, NB medium absorbance was normalized to 0.5 at 620 nm, and cells were removed using 0.22 μm filtration. For hemolysis detection, sheep blood was diluted to 5% in phosphate buffer solution (pH 7.4) and kept in a water bath at 37°C for 30 min. Then, 500 μL of the filtrate was mixed with 50 mL of blood suspension and incubated at 37°C for 20 min followed by incubation and centrifugation (3000 × g for 10 min). Absorbance at 405 nm was measured against control groups. The assay was performed in triplicate, and the hemolysis percentage was calculated as follows:
AS is the OD405 value of the metabolite-added group, A0 is the OD405 value of the blank control group, and A100 is the OD405 value of the positive control group.
Quantitative real-time polymerase chain reaction (RT-PCR)
B. cereus cells were collected during the logarithmic phase and then incubated in NB broth with 105 CFU/mL. The concentration of the metabolites in the broth was adjusted to 125 μg/mL. B. cereus was then cultivated in a shaker at 37°C and 160 rpm/min for 8 h. After cultivation, total RNA was extracted using a Bacteria Total RNA Isolation Kit (Sangon Biotech, Shanghai, China), and a One Step RT-qPCR Kit (Sangon Biotech) was used according to the manufacturer’s instructions. The expression of the genes presented in Supplementary Table S3 was determined using a LightCycler 96 PCR System from Roche as described previously (Park et al., 2019). A 16S rRNA gene was used as an internal reference, and all quantitative RT-PCRs were repeated three to five times. Relative gene expression was calculated by the 2−ΔΔCT method (Jeßberger et al., 2015).
Statistical analysis
All data were expressed as mean ± standard deviation of triplicates. Significant differences were calculated by Duncan’s multiple range test using SPSS 22.0.0.0 software (SPSS, Inc., Chicago, IL, USA). Results were considered statistically significant at p < 0.05.
Results and Discussion
The interactions between PlcR and garlic metabolites
The QuickVina-W was applied to study the protein–ligand interactions between PlcR and 82 garlic metabolites through blind molecular docking. The 18 metabolites with the highest affinity and their binding energies are listed in Table 1, ranging from −6.1 to −8.9 kcal/mol. Nine metabolites, with binding energies less than or equal to PapR7 (−6.5 kcal/mol), could potentially compete with PapR7 for the active site, making them promising QS inhibitors of B. cereus. Notably, the metabolites with the lowest binding energies (quercetin, kaempferol, luteolin, flavone, and rutin) are flavonoids. Flavonoids, secondary metabolites with variable phenolic structures, are abundant in garlic leaves (above 250 mg/100 g DM) (Li et al., 2023; Skoczylas et al., 2023). They have been shown to disrupt QS in Gram-negative bacteria (e.g., Pseudomonas aeruginosa and Chromobacterium violaceum) by antagonizing LasR and RhlR receptors or acting as competitive inhibitors (Paczkowski et al., 2017). This disruption reduces toxins, biofilms, and swarming motility. However, B. cereus, a Gram-positive bacterium, uses AIP as signaling molecules, differing significantly from Gram-negative bacteria in signal processing, transmission, and receptor binding. The potential of flavonoids to interfere with B. cereus QS requires further validation.
The Highest Affinity Compounds to PlcR in Garlic Metabolites
Using LigPlot+ software, the intermolecular bonds between PlcR and the five metabolites with the highest binding energies, alongside PapR7, were visualized (Fig. 1). Quercetin counts about 3.4 mg/100 g DM in garlic leaves (Skoczylas et al., 2023) and formed hydrogen bonds with Cys227, Arg228, and Asn230 of PlcR and had hydrophobic interactions with Lys87, Gln88, and Met272 (Fig. 1A); Luteolin has about 2.1 mg/100 g DM in garlic leaves (Skoczylas et al., 2023) and formed hydrogen bonds with Asn230, Ala233, Gln237, and Ala274 of PlcR and had hydrophobic interactions with Lys87, Gln88, Leu234, Glu271, Met272, His273, and Tyr275. Interestingly, luteolin and PapR7 (Fig. 1F) both bound to Leu234, Gln237, and Tyr275 of PlcR noncovalently, suggesting that luteolin could compete with PapR7 for the binding site and affect the QS process of B. cereus (Fig. 1B); Kaempferol (about 1.8 mg/100 g DM in garlic leaves; Skoczylas et al., 2023) also bound to Tyr275 of PlcR noncovalently like PapR7 (Fig. 1F), indicating a possible competition at the binding site (Fig. 1C); and Flavone formed a hydrogen bond with Gln88 of PlcR and had hydrophobic interactions with Lys87, Asn230, Ala233, Glu271, Met272, His273, Ala274, and Tyr275. It shared the same binding site with PapR7 (Fig. 1F) at Tyr275 (Fig. 1D); Rutin counts about 24.1 mg/100 g DM in garlic leaves (Skoczylas et al., 2023) and formed hydrogen bonds with Gln88, Cys227, Asn230, Ala233, and Leu270 of PlcR and had hydrophobic interactions with Met84, Lys87, Arg228, Gln237, Glu271, and Met272. It could also compete with PapR7 (Fig. 1F) for the same binding site at Gln237 (Fig. 1E).
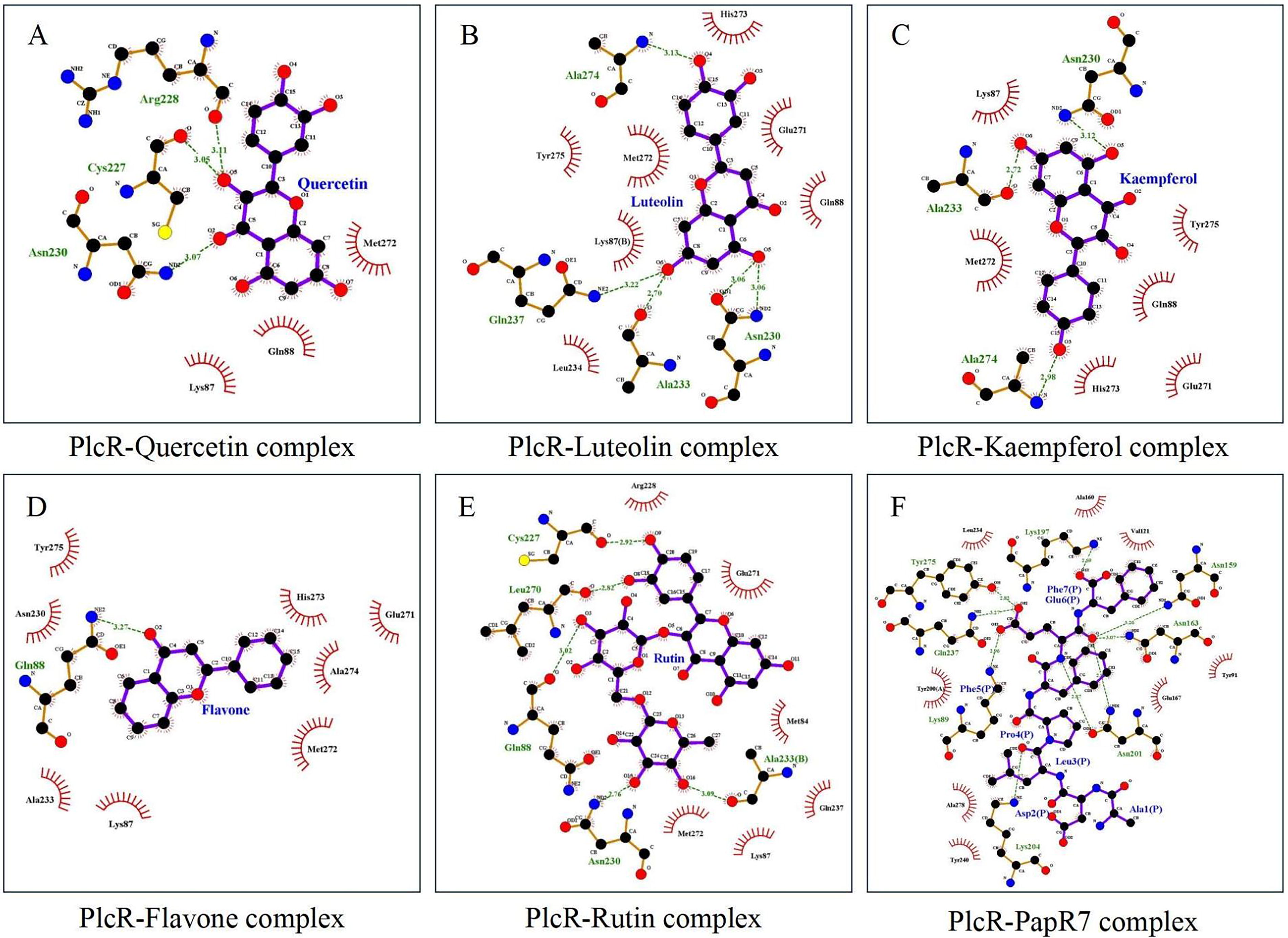
The intermolecular bonds between PlcR and the metabolites. The green dotted lines indicate the hydrogen bonds that have formed, with the numbers above them representing the distances of these bonds. The semicircles with vertical lines illustrate the hydrophobic interactions between molecules.
Impact of the flavonoids on the enterotoxin production and hemolytic activity of B. cereus
To further verify the effects of the five flavonoids on the QS process of B. cereus, a series of physiological experiments were conducted. The NIC of the flavonoids was determined (Supplementary Table S3). We then evaluated the effects of these flavonoids on toxin production by measuring bacterial counts and toxin production times at the NIC (Fig. 2). The control produced Nhe at 5.5 h (5.80 lg CFU/mL) and Hbl at 6 h (6.10 lg CFU/g) at 37°C. Quercetin, flavone, and rutin delayed the toxin production as follows: flavone produced Nhe and Hbl at 6.5 h and 7 h, respectively; rutin produced both toxins at 6.5 h; and quercetin produced Nhe at 6.5 h. The bacterial count needed for toxin production also increased with these flavonoids, suggesting that they may inhibit the QS process of B. cereus, as predicted by molecular docking. Previous studies indicated that B. cereus poses a health risk in food when exceeding 5 lg CFU/g (Stenfors Arnesen et al., 2008). Our results confirmed that B. cereus produced Nhe and Hbl at this level. Quercetin, flavone, and rutin increased the time and load for toxin production, which could reduce the foodborne illness risk. Surprisingly, luteolin and kaempferol enhanced the toxin production, producing Nhe or Hbl earlier (Nhe: 4.5 h for luteolin and 5 h for kaempferol; Hbl: both 5 h) and with lower counts than the control, indicating that they may act as AIP analogs, stimulating the QS process of B. cereus.
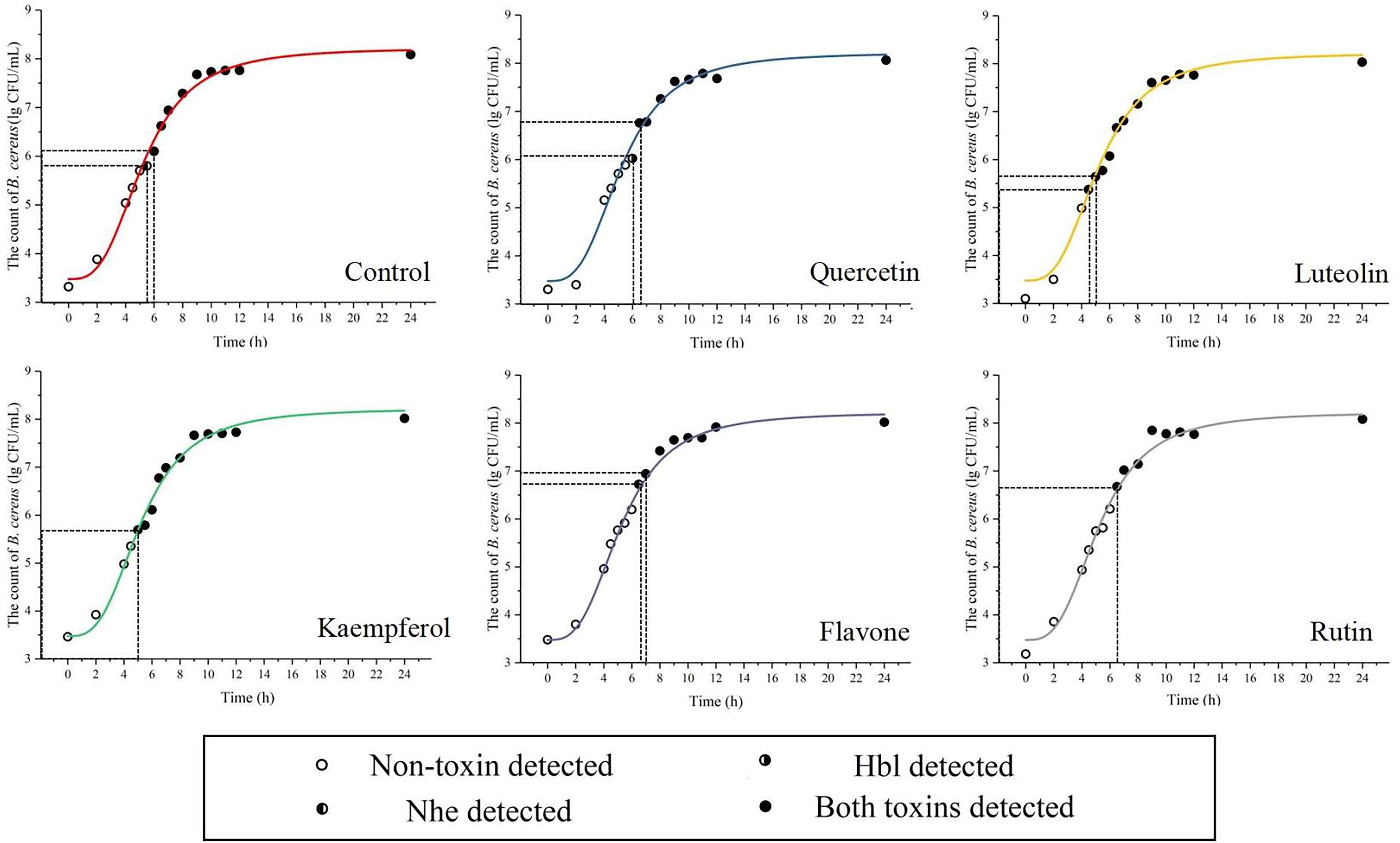
Impact of the flavonoids on the enterotoxin production of Bacillus cereus. The growth of B. cereus was evaluated by the Gompertz model.
The effects of five flavonoids on the hemolytic properties of B. cereus were also tested at the NIC (Fig. 3). After 8 h of culture, the hemolytic properties of B. cereus treated with flavone and rutin were reduced to 44.05% and 67.81% of the control, respectively, while those treated with luteolin and kaempferol were increased to 153.03% and 139.31% of the control, respectively. The quercetin-treated group showed no significant difference from the control. The hemolysis results were consistent with the Hbl production, confirming that flavone and rutin can effectively inhibit the toxicity of B. cereus, while luteolin and kaempferol can enhance it. As the incubation time increased, the inhibition of B. cereus hemolytic activity by flavone and rutin gradually weakened. After 16 h of culture, the hemolytic activity of the flavone-treated group was 53.22% of the control; after 24 h, it was 79.82% of the control. The hemolytic activity of the rutin-treated group was 85.17% of the control after 16 h; there was no significant difference between the rutin-treated group and the control group after 24 h. Since flavone and rutin did not inhibit the growth and reproduction of B. cereus at the NIC, the number of B. cereus cells increased exponentially as the culture progressed. Consequently, flavone and rutin were gradually unable to interfere with the QS process of B. cereus at this concentration, weakening their inhibition of B. cereus hemolysis. In addition, rutin’s poor stability in air may further reduce its inhibitory effect on the hemolytic activity (Tobar-Delgado et al., 2023).
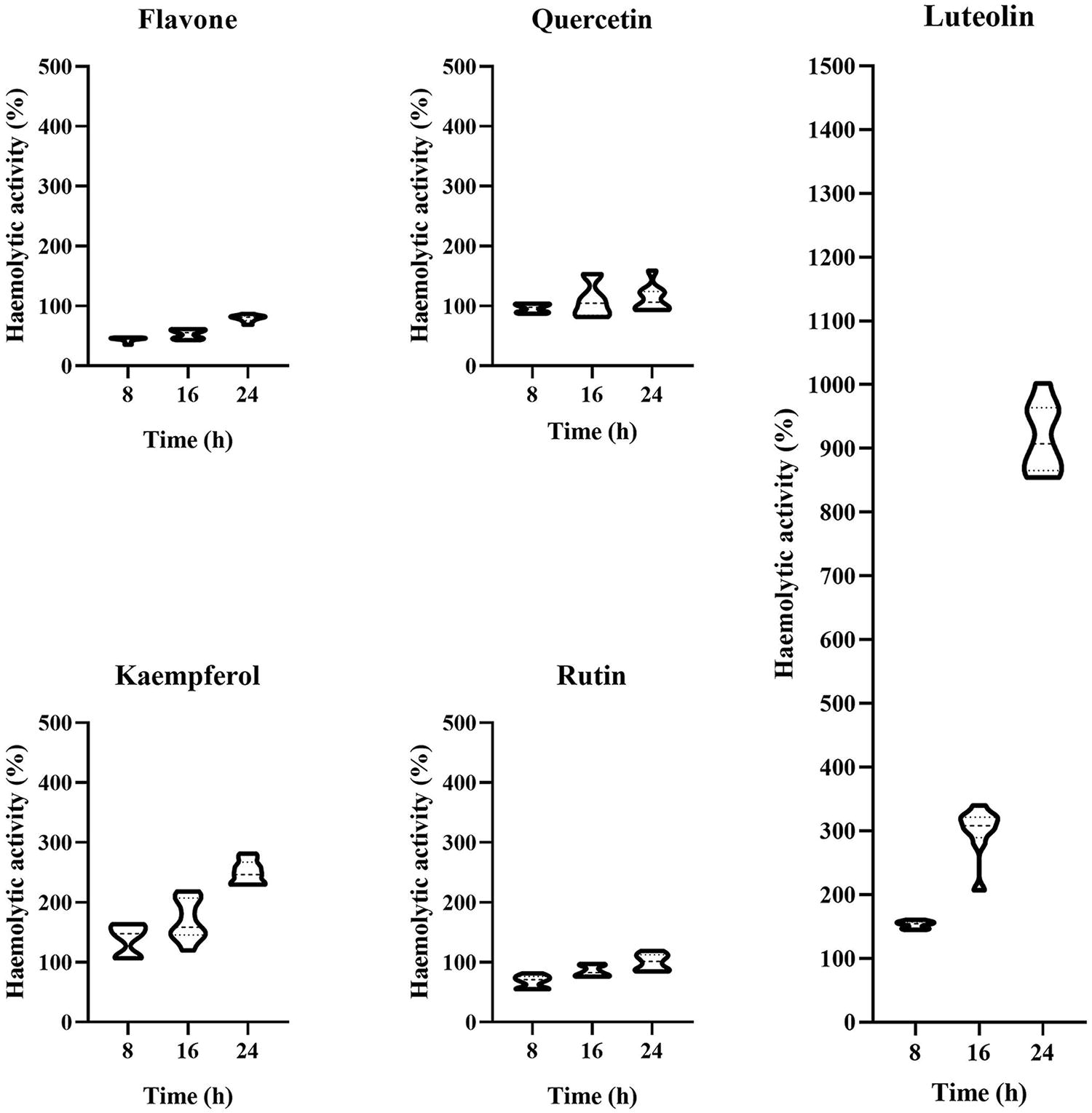
Effect of the five flavonoids on the hemolytic properties of B. cereus at the NIC. Hemolytic activity exceeding 100% indicates that the substance enhances the toxin production of B. cereus. Conversely, a value below 100% suggests that it inhibits toxin production. NIC, noninhibitory concentrations.
Effects of the flavonoids on the expression of QS-related genes
The expression of QS-related genes in response to flavonoids is shown in Figure 4, and the QS pathway is illustrated in Figure 5. The QS mechanism of B. cereus involves the following steps: PapR protein is synthesized and secreted by the bacteria, then cleaved by the extracellular NprB protease to produce AIPs (PapR7). These AIPs are transported back into the B. cereus cells via oligopeptide permease (OPP) and bind to PlcR, which triggers its conformational changes, oligomerization, and DNA binding, leading to the expression of different virulence factors (such as nhe, hbl, and cytK), as well as papR, plcR, nrpB, and other genes (Rutherford and Bassler, 2012). Flavone and rutin treatment reduced plcR gene expression compared with the control group, while luteolin and kaempferol treatment increased it significantly (Fig. 4A). The expression levels of virulence factors also changed accordingly as follows: flavone treatment decreased nhe, hbl, and cytK expression levels, rutin treatment decreased nhe and hbl expression levels, and luteolin and kaempferol treatment increased hbl and cytK expression levels significantly. The experimental results agree with the previous phenotypic studies (toxin production, hemolysis), suggesting that flavone and rutin can interfere with the QS of B. cereus by competing with AIP (PapR7) for the PlcR binding site, whereas luteolin and kaempferol act as AIP analogs and bind to PlcR to enhance the QS process (Fig. 5). This phenomenon should be related to the binding sites of flavone, rutin, luteolin, and kaempferol with PlcR. According to prior studies, the activation of B. cereus QS depends on the Glu6 and Phe7 of PapR7 (Yehuda et al., 2018). Alterations in these two amino acids will cause significant changes in the toxicity of B. cereus (lower or higher) or even lose the ability to trigger QS. Interestingly, Glu7 of PapR7 forms a hydrogen bond with PlcR, whereas flavone has a hydrophobic interaction with Tyr275, rutin has a hydrophobic interaction with Gln237, luteolin can form a hydrogen bond with Gln237 and hydrophobic interaction with Tyr275, and kaempferol also has a hydrophobic interaction with Tyr275 (Fig. 1). These similar or opposite forces may be the key for flavone, rutin, luteolin, and kaempferol to influence the QS process of B. cereus. Moreover, results indicated that flavone and rutin could further inhibit the QS of B. cereus by downregulating oppC expression and reducing PapR7 uptake.
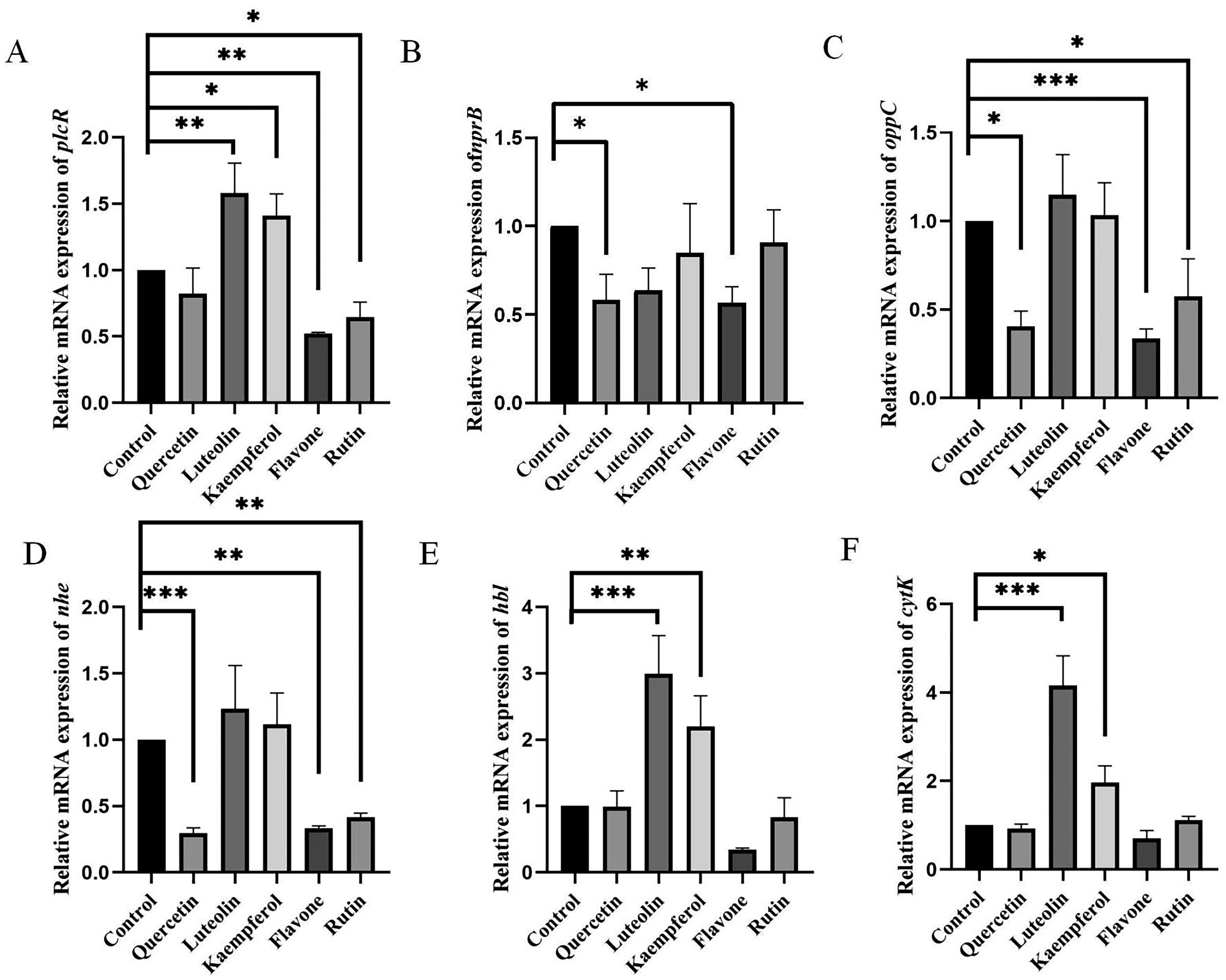
The expression of QS-related genes in response to the five flavonoids. A single asterisk (*) denotes a p-value <0.05, indicating statistical significance. Two asterisks (**) signify a p-value <0.01, and three asterisks (***) represent a highly significant p-value <0.001. QS, quorum sensing.

B. cereus QS circuit. The QS circuit is initiated by the secretion of pro-AIP PapR, which is processed into mature AIP by NprB. After re-entry into the cell via OPP, AIP activates PlcR, influencing virulence factor synthesis and papR expression. Compounds like flavone and rutin disrupt, while luteolin and kaempferol enhance, this QS process by interacting with PlcR. AIP, autoinducing peptide; OPP, oligopeptide permease.
Despite the phenotypic effects of quercetin on toxin production and hemolysis of B. cereus, this compound did not show clear evidence of enhancing QS at the molecular level (Fig. 4A, no difference with control in plcR expression). However, a significant downregulation of oppC expression in quercetin treatment group could lead to barriers in AIP absorption and thus downregulate nhe and other toxin gene expression. Moreover, toxin production of B. cereus is also regulated by other factors such as HlyII and RpoN (Hayrapetyan et al., 2015; Stenfors Arnesen et al., 2008). The changes in toxicity of B. cereus in quercetin treatment groups might be related to other pathways as well.
This study identified and validated quercetin, kaempferol, luteolin, flavone, and rutin in garlic as potential QS inhibitors of B. cereus targeting the PlcR receptor. These compounds had significantly lower binding energy (≤−8.2 kcal/mol) to PlcR than its ligand PapR7 (−6.5 kcal/mol). Among them, kaempferol, luteolin, flavone, and rutin exhibited different or opposite intermolecular interactions with PapR7 at the Gln237 or Tyr275 residues of PlcR, which resulted in the suppression or enhancement of the QS process, respectively. Flavone and rutin reduced enterotoxin secretion and hemolysis without affecting bacterial growth, whereas kaempferol and luteolin increased QS-mediated virulence. Therefore, flavone and rutin were identified as the main QS inhibitors in garlic based on their effectiveness at both phenotypic and molecular levels.
Footnotes
Authors’ Contributions
Z.J.: Methodology, investigation, data curation, and writing—original draft. S.Z.: Investigation. H.L.: Investigation. Q.O.: Validation, supervision, and writing—review and editing. N.T.: Conceptualization, resources, supervision, and writing—review and editing.
Disclosure Statement
There is no conflict of interest, and all the coauthors have agreed for submission of their work to this journal.
Funding Information
This study was supported by the National Natural Science Foundation of China, grant number
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.