
Abstract
The plasmid-mediated tet(X4) gene has exhibited a high-level resistance to tigecycline (TGC), which has raised concerns globally regarding antibiotic resistance. Although the widespread tet(X4) has been found widely in Escherichia coli, it is scarcely found in other Enterobacteriaceae. This study aimed to characterize a ST469 Salmonella enterica serovar Rissen (S. Rissen) isolate harboring tet(X4) from pork, which was identified and characterized via antimicrobial susceptibility testing, conjugation assays, plasmid curing testing, whole-genome sequencing, and bioinformatic analysis. Ten ST469 S. Rissen isolates of 223 Salmonella spp. isolates were isolated from food samples in China during 2021–2023. One of 10 S. Rissen isolates, SM2301, carrying tet(X4) conferred high-level resistance to TGC (minimum inhibitory concentration > 8 µg/mL). The tet(X4) could be conjugated into different recipients, including E. coli, S. enteritidis, and K. pneumoniae isolates. Plasmid curing confirmed that tet(X4) was plasmid-mediated. Genetic analysis revealed that the tet(X4) in the SM2301 isolate was located in the IncFIA(HI1)-IncHI1A-IncHI1B(R27) hybrid plasmid, and the structure of tet(X4) was abh-tet(X4)-ISCR2. To the best of our knowledge, this is the first report of a tet(X4)-positive food-derived S. Rissen isolate. The extending bacterial species of tet(X4)-bearing plasmids suggested the increasing transmission risk of the mobile TGC resistance gene tet(X4) beyond E. coli. This study highlights the emerging and evolution risk of novel resistance genes across various bacterial species. Therefore, further surveillance is warranted to monitor the prevalence of tet(X4) in Salmonella spp. and other bacterial species.
Introduction
Salmonella is an important zoonotic pathogen that causes various infections and economic burden among humans worldwide (Sanni, et al., 2024). Salmonella isolates exhibiting multiantibiotic resistance have been frequently reported (Kakooza, et al., 2021; Huang et al., 2013; Lu et al., 2019), particularly carbapenemase-producing and mcr-1-positive Salmonella isolates (Falagas et al., 2008). Infections caused by these multidrug-resistant bacteria are difficult to treat. One of the few available drugs that can be used to treat infections caused by extensively drug-resistant pathogens is tigecycline (TGC; Brust et al., 2014).
TGC, first reported by Sum in 1999, represents a new class of antibiotic agents (Sum & Petersen, 1999) and was used first in the United States in 2005. It exhibited an extraordinary broad spectrum of activity against pathogens. In China, TGC was approved for use in 2011 (Jiang, 2022). However, TGC resistance has rapidly occurred in the following years mainly due to chromosome-encoding mechanisms, such as efflux pump overexpression and ribosome protection (Gerson et al., 2018; He et al., 2018), which mostly exhibited low-level resistance to TGC. Until 2019, high-level TGC resistance of tet(X3) and tet(X4) mediated by plasmid in Enterobacteriaceae and Acinetobacter was first reported by He et al. (2020). Subsequently, the emergence of highly resistant TGC genes, namely, tet(X5) and tet(X6), has been reported (He et al., 2020; Liu et al., 2020). A retrospective study demonstrated that tet(X3) and tet(X4) were widely distributed not only in China but also worldwide, such as in Thailand and England (Jiang, 2022). This raised concerns globally regarding antibiotic resistance.
Clinical isolates of Salmonella resistant to TGC have also been reported (Hentschke et al., 2010). Despite the mobile resistance gene tet(X4) in Salmonella from patients having been studied (Wang et al., 2021), little is known about the spread and distribution of Salmonella with tet(X4) in food. S. Rissen is one of the most common serovars found in pigs and pork products in different countries, including China (Raquel et al., 2016; Qiu et al., 2019). Furthermore, S. Rissen isolates have been reported to be strongly associated with pork food products and were dominant in the collection studied (Campos et al., 2016). Therefore, in this study, we analyzed TGC-resistant Salmonella from food and showed the molecular characteristics of one S. Rissen isolate with plasmid harboring tet(X4) from pork in China, which could potentially increase the risk of infection and treatment failure in humans (He et al., 2019) and pose further threat to public health.
Materials and Methods
Isolate collection
A total of 223 Salmonella spp. isolates were collected from food in the period of 2021–2023 in Shanghai, China (Supplementary Table S1). Salmonella enteritidis SM2211 isolate from food, Escherichia coli EC1905 isolate from food, and Klebsiella pneumoniae KP2297 isolate from a patient were used as recipients in conjugation experiments. SM2211 isolate without a plasmid, EC1905 isolate with a plasmid, and KP2297 isolate with a plasmid were susceptible to TGC and resistant against gentamicin (GEN; Tables 1 and 2), which could be used to screen transconjugants.
Basic Information of Three Recipient Isolates in Conjugation Experiments
Minimum Inhibitory Concentrations of Donor Isolates, Recipient Isolates, and Transconjugants
Antimicrobial susceptibility testing
Using the broth microdilution method, we determined the minimum inhibitory concentrations (MICs) of minocycline (MIN, 0.5–64 µg/mL), ciprofloxacin (CIP, 0.016–32 µg/mL), ampicillin (AMP, 1–32 µg/mL), polymyxin E (CT, 0.25–8 µg/mL), chloramphenicol (CHL, 2–64 µg/mL), (TGC, 0.25–32 µg/mL), (GEN, 1–32 µg/mL), tetracycline (TET, 1–32 µg/mL), doxycycline (DOX, 0.5–64 µg/mL), polymyxin B (PB, 0.25–8 µg/mL), imipenem (IPM, 0.25–8 µg/mL), cefotaxime (CTX, 1–32 µg/mL), and cefepime (FEP, 2016 µg/mL) of all S. Rissen isolates. Using antibiotic gradient diffusion strip methods (E-test, bioMérieux, https://www.biomerieux.com), the MICs of the TGC-resistant isolates and transconjugants were further determined. The results were interpreted according to the 2022 guidelines of the Clinical and Laboratory Standards Institute (2022), except that the results of MICs against TGC were interpreted according to the European Committee on Antimicrobial Susceptibility Testing breakpoint for TGC (http://www.eucast.org.clinical_breakpoints). E. coli ATCC 25922 served as the quality control isolate.
Conjugation experiments
To investigate the transferability of tet(X4), conjugation assays were conducted as previously described (Li et al., 2021). Briefly, the SM2301 carrying tet(X4) was used as the donor and E. coli EC1905, K. pneumoniae PK2297, and S. enteritidis SM2211 as the recipients. First, the donor and recipients were cultured at 36°C overnight. Second, 0.5 McFarland density cultures of donors and recipients were mixed at a ratio of 1:1. Then, 0.1 mL of the mixed cultures was plated onto tryptose soya agar (TSA) plates containing GEN (8 μg/mL) and/or TGC (1 μg/mL) antimicrobials. Third, after cultures were kept at 36°C for 24 h, individual bacterial colonies were plated on the same TSA plate with antimicrobials. Lastly, after being cultured at 36°C for 24 h, the transconjugants carrying tet(X4) were determined via reverse transcription–polymerase chain reaction (RT-PCR) of tet(X4) and corresponding resistant phenotyping. The frequencies of conjugative transfer were calculated by the number of transconjugants per recipient. The primers and probe of tet(X4) were as follows: forward primer: 5′-GAAAGAGACAACGACCGAGAG-3′; reverse primer: 5′-CACCCATTGGTAAGGCTAAGT-3′; and probe: FAM-CCTACACAAAGGTTCAGGTCAGGAAGC-BHQ1.
Curing experiment
The curing experiment was conducted as previously described with minor modifications (Ghosh et al., 2000). For plasmid elimination, isolates were cultured overnight at 36°C in nutrition broth (NB). Then, 50 μL of the cultured isolates was taken at a ratio of 1:100 to 5-mL NB containing subinhibitory concentrations (2.5 μg/mL) of novobiocin. Subsequently, the culture was incubated for 24 h at 40°C, followed by 14 subcultures (1% inoculum) in the same medium containing novobiocin. After growth under these conditions, the cultures were plated on a nutrient agar plate, and single colonies from such cultures were isolated and subjected to plasmid identification, which was determined via RT-PCR using tet(X4). The S. Rissen isolate without plasmid was named SM2301-P.
Genomic DNA extraction and whole-genome sequencing
In this study, 10 S. Rissen isolates including SM2301 have been obtained from food; the genetic relationship among them could be analyzed using core genome multilocus sequence typing (cgMLST). Therefore, the 10 S. Rissen isolates were used for short-read sequencing. These isolates were cultured overnight. Genomic DNA was extracted using the DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s protocol with minor modifications. The cells were prelysed with lysozyme for 30 min. DNA concentration, quality, and integrity were identified using a Qubit fluorometer (Thermo Scientific Waltham, MA, USA). Then, genomic DNA was sent out for short-read sequencing (next generation). The Illumina HiSeq platform (Illumina, USA) was utilized to generate short-read sequencing data with the 2 × 150 strategy. The quality of sequence data generated by Illumina sequencing was evaluated using FastQC v0.11.2 (Cambridge, London, UK). SPAdes version 3.13.0 was used to construct the draft genomes. S. Rissen carrying tet(X4) was further sent out for nanopore long-read sequencing using PromethION (Oxford Nanopore, England). The quality of sequence data generated by PromethION nanopore sequencing was evaluated using Porechop v0.2.4. Unicycler version 4.8 (Wick et al., 2017) was used to generate complete genomes including plasmid and determine the genetic environment and location of tet(X4) in plasmid by informatics analysis.
Bioinformatic analysis
Short-read raw sequences of S. Rissen isolates were de novo assembled using CLC Genomics Workbench v7.0 (CLC Bio, Aarhus, Denmark). The reads were trimmed using Trimmomatic v0.36. Then, the trimmed reads were assembled using BioNumerics v7.6 (Applied Maths, Kortrijk, Belgium), and the assembled sequence was used for further analysis. De novo assembly was performed utilizing the long-read data and short-read data using Canu v1 and GATK v1.6-13.
Subsequently, the Rapid Annotation using Subsystems Technology server (https://rast.nmpdr.org/rast/cgi) was then used to annotate the completed genome (Bai et al., 2019; Wang et al., 2022). MLST was determined using an online tool (https://pubmlst.org/bigsdb?db=pubmlst_salmonella_seqdef&page=sequenceQuery). Furthermore, cgMLST was performed based on the whole-genome sequencing (WGS) tool of BioNumerics v7.6 (Applied Maths) with an integrated 3002 loci cgMLST scheme. The threshold values for interpreting clonality with cgMLST were as follows: ≤ 10 allelic differences, related (https://enterobase.warwick.ac.uk/species/index/senterica).
Plasmid replicons and antimicrobial resistance genes were analyzed using PlamidFinder v2.1 and ResFinder v4.1 (https://cge.cbs.dtu.dk/services/). Furthermore, circular comparisons between plasmids were performed using the BRIG tool v0.95 (Alikhan et al., 2011). Genetic comparison was also performed using Easyfig v2.2.3 (Sullivan et al., 2011).
Results
Identification of isolates and resistance phenotypes
A total of 10 (4.48%, 10/223) S. Rissen isolates were detected in Shanghai, China, in the period of 2021–2023. Only one of 10 S. Rissen isolates, SM2301 from pork, carried tet(X4). Other 213 Salmonella spp. did not carry tet(X4).
All 10 S. Rissen isolates were identified as ST469 (Table 3). Of these isolates, two carried plasmids, whereas the other eight did not have any plasmids. The SM2301 isolate had an IncFIA(HI1)-IncHI1A-IncHI1B(R27) replicon type of plasmid, whereas the SM2249 isolate had an IncY replicon type of plasmid. Except for the SM21208 isolate, the other nine isolates carried different resistance genes, such as dfrA12, aadA2, sul3, bla TEM-1, qnrS1, aadA22, floR, and lnuG (Table 3). Most of them conferred resistance to AMP, CHL, TET, and MIN (Table 4). In addition, the SM2301 isolate was resistant to DOX. However, most S. Rissen isolates were susceptible to CIP, CT, TGC, GEN, DOX, PB, IPM, FEP, and CTX (Table 4).
Basic Information of Plasmids and Resistant Genes in 10 S. Rissen Isolates
Minimum Inhibitory Concentrations of 10 S. Rissen Isolates in This Study
Content Footnote: CIP, ciprofloxacin; AMP, ampicillin; CT, polymyxin E; CHL, chloramphenicol; TGC, tigecycline; GEN, gentamicin; TET, tetracycline; MIN, minocycline; DOX, doxycycline; PB, polymyxin B; IPM, imipenem; CTX, cefotaxime; FEP, cefepime.
Transferability of the tet(X4) gene
To investigate the transferability of tet(X4), we subjected the SM2301 isolate to conjugation experiments with E. coli EC1905, K. pneumoniae KP2297, and S. enteritidis SM2211 (Table 2). The TGC-resistant gene tet(X4) could be successfully transferred from SM2301 into the recipients E. coli EC1905, K. pneumoniae KP2297, and S. enteritidis SM2211, with transfer efficiency values of ∼10−7, ∼10−6, and ∼10−6, respectively. The transconjugants were named EC1905-tetX4, KP2297-tetX4, and SM2211-tetX4, respectively, showing at least eight-fold increases in MICs against TGC in comparison with the recipients (Fig. 1). These results indicated that tet(X4) was located in plasmid or other mobile genetic elements (MGEs). Meanwhile, all three transconjugants exhibited resistance to all tetracycline antibiotics tested in this study, with MICs of 8–64 mg/L for MIN, TET, and DOX, representing at least eight-fold increases in MIC compared with their recipients (Table 2).

Antimicrobial susceptibility testing using E-test. It presents the recipients and transconjugants that are resistant to tigecycline.
The recipient EC1905 was resistant to polymycin (PB = 4 mg/L, CT = 4 mg/L), which was mediated by mcr-1.1 (Table 2). Therefore, transconjugants EC1905-tetX4 exhibited resistance to both PB and TGC, which might result in untreatable infections. TGC and PB are considered as few of the “last-resort” antibiotic defenses used in antiinfection therapies against multidrug-resistant bacteria (Xu et al., 2021). Furthermore, pSM2301-tetX4 was successfully cured from SM2301, and SM2301-P was acquired.
TGC resistance of isolate with plasmid curing
Bacterial plasmids are known to harbor genes for resistance to antibiotics, and curing plasmids from bacteria is a way to eliminate the bacteria plasmid and determine the antibiotic resistance mediation (Letchumanan et al., 2015). Therefore, in this study, the plasmid pSM2301-tetX4 in S. Rissen SM2301 was successfully cured, as confirmed by RT-PCR using the tet(X4) gene, and was named SM2301-P. SM2301-P exhibited a 64-fold decrease, from 16 µg/mL to 0.25 µg/mL, compared with SM2301 in the MIC of TGC. (Fig. 2). This result indicated that resistance against TGC of SM2301 was affected by plasmid curing; therefore, SM2301 carrying tet(X4) could be classified as “plasmidial” (Letchumanan et al., 2015). The high-level resistance against TGC of SM2301 isolate was mediated by plasmid carrying tet(X4). The plasmid has a potential risk of horizontal dissemination by conjugation between different species.

Curing of plasmid pSM2301-tetX4. SM2301 and SM2301-P isolates are resistant to tigecycline.
Clonal relationship analysis based on WGS
The 10 ST469 isolates were characterized by WGS using the Illumina HiSeq 2500 platform to generate 2 × 150-bp paired-end read data. The diversity of cgMLST clusters of these isolates is presented in Figure 4. Among the 10 S. Rissen isolates, the number of allelic differences ranged from 17 to 117 (Fig. 3), suggesting that these isolates do not belong to the clonal group, with more than 10 allelic differences (Hoff et al., 2022; Ripolles-Avila et al., 2020).
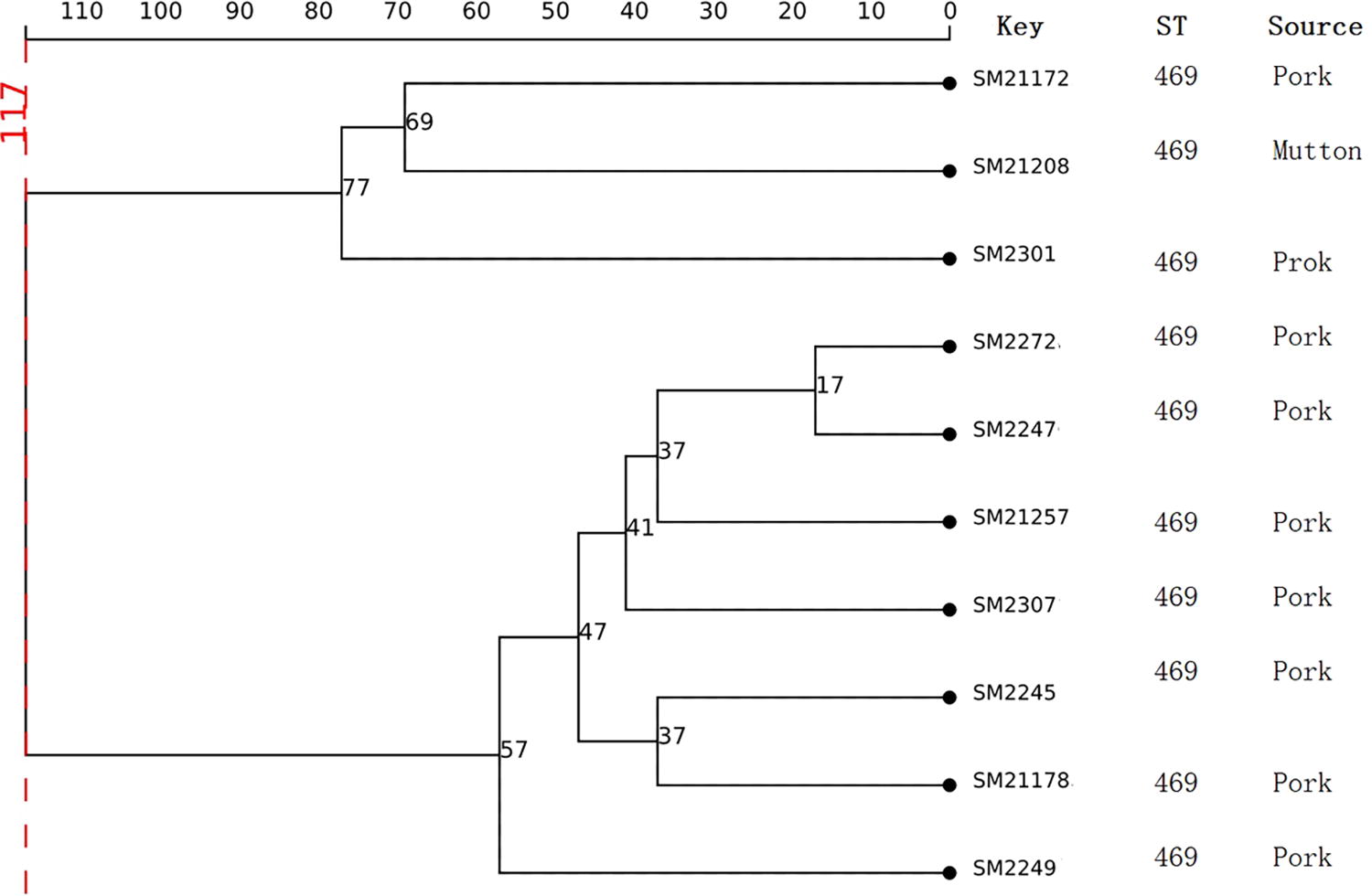
Clustering of 10 ST469 S. Rissen isolates based on core genome. The value represents the number of allelic differences among the 10 isolates. The corresponding data, including the name of the isolates (Key), ST, and Source, are displayed on the right side of the dendrogram.
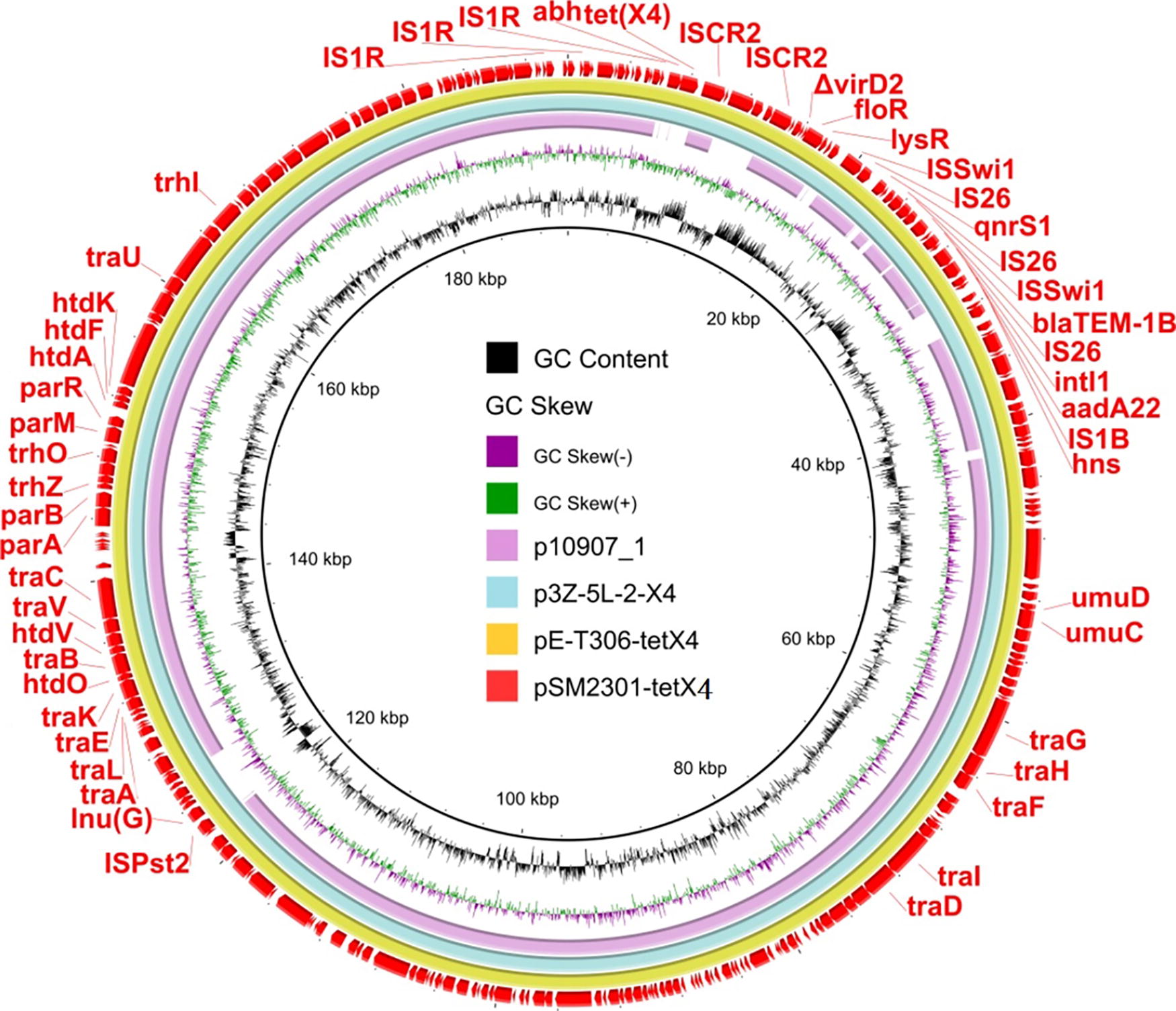
Genetic context of tet(X4) in the IncFIA(HI1)-IncHI1A-IncHI1B plasmid. Circular comparison between the tet(X4)-bearingIncFIA(HI1)-IncHI1A-IncHI1B plasmid in the study and other homologous plasmids p10907_1, p3Z-5L-2-X4, and pE-T306-tetX4 in the NCBI database.
Sequence features of nanopore sequencing
To learn the genetic contexts of tet(X4) of SM2301, we carried out PromethION nanopore long-read sequencing to obtain complete genome sequences of the SM2301 isolate. A total of 8,738,130 reads with 1274 Mb were obtained. The mean read length of the total sequencing data was 7273 bp. The read length, N50 of the total data, was 9857 bp. The longest read length was 86,760 bp. The nanopore long-read sequencing data and Illumina short-read sequencing data of SM2301 were used to perform de novo assembly using a hybrid strategy to obtain the completed chromosomes and plasmids. Bioinformatic analysis showed that the SM2301 isolate harbored a chromosome and one plasmid pSM2301-tetX4 (Table 3).
Genetic environment analysis of tet(X4)
The tet(X4) gene was located in the plasmid pSM2301-tetX4. Plasmid replicon analysis revealed that it belonged to the IncFIA (HI1)/IncHI1A/IncHI1B(R27) hybrid plasmid group carrying IV secret system (T4SS; Fig. 4). BLASTn search revealed that pSM2301-tetX4 exhibited 100% nucleotide identity and 100% query coverage to the sequence of plasmid p3Z-5L-2-X4 in K. pneumoniae 3Z-5L (GenBank: CP072517). Analysis also showed that pSM2301-tetX4 exhibited 99.97% nucleotide identity and 100% query coverage to the sequence of plasmid pE-T306-tetX4 in E. coli E-T306 (GenBank: CP090284) and 99.88% nucleotide identity and 92% query coverage to the sequence of plasmid p10907_1 in Salmonella spp. 01_0907 (GenBank: CP090284). Except for tet(X4), pSM2301-tetX4 carried other resistance genes, including qnrS1, bla TEM-1B, aadA22, floR, and lnuG. However, the chromosome of SM2301 carried dfrA12, aadA2, sul3, and bla TEM-1 (Fig. 4).
We then examined the tet(X4)-neighboring genetic elements in pSM2301-tetX4 and found that the structure of tet(X4) was abh-tet(X4)-ISCR2 (Fig. 4). ISCR2 has been reported to be associated with the acquisition of diverse antimicrobial resistance genes (Toleman et al., 2006). It could adopt a rolling-circle transposition process to transpose adjacent DNA sequences (Toleman et al., 2006). Unlike other plasmids carrying tet(X4), the abh-tet(X4)-ISCR2-ISCR2 element in pSM2301-tetX4 was found to be located upstream of two ISCR2 copies (Fig. 5), which was different from the previous finding (Chen et al., 2019), indicating that tet(X4) on pHNGS471-2 was located between two ISCR2 copies that form a circular structure (Fang et al., 2019).
The tet(X4)-harboring genetic context in pSM2301-tetX4 was abundant in MGEs, such as insertion sequences and composite transposon, which were flanked by tet(X4) (Fig. 4).
Discussion
Salmonella is an important pathogen that causes foodborne illnesses to humans worldwide (Rincón-Gamboa et al., 2021). It is also a major reservoir of genes that exhibit antimicrobial resistance. The World Health Organization declared Salmonella as a high-priority pathogen owing to its increased resistance to first-line antibiotics, fluoroquinolones, and third-generation cephalosporins (Tacconelli et al., 2018; Thames & Sukumaran, 2020). It has also been recognized to pose a risk to human health, thus necessitating the development of new antibiotics (Tacconelli et al., 2018). In Salmonella, tet(X4) has a very low prevalence. It was first detected in a stool sample of a patient in China in 2019 (Wang et al., 2021). To the best of our knowledge, the present study is the first to report on tet(X4)-positive S. Rissen isolate in food (pork liver) from China. Enterobacteriaceae that is tet(X4)-positive, including E. coli and K. pneumoniae, were detected in samples from humans, animals, and their surrounding environment from different geographical regions (He et al., 2019; Sun et al., 2019; Zhai et al., 2022). These findings suggest that tet(X4)-encoding TGC resistance has already spread throughout China.
The ST469 S. Rissen isolate is a commonly reported serovar. In the present study, the cgMLST results indicated that the 10 ST469 isolates do not belong to the clonal group (>17 allelic differences; Fig. 3). Furthermore, most of the ST469 S. Rissen isolates (80%, 8/10) did not have plasmids (Table 3). Given this difference, we speculated that the ST469 S. Rissen isolate has obtained tet(X4) by horizontal transfer. tet(X4) is considered to be the most predominant plasmid-mediated TGC-resistant gene in the Enterobacteriaceae family, particularly in E. coli (Gao et al., 2022). Furthermore, in this study, plasmid pSM2301-tetX4 carrying tet(X4) in S. Rissen could be successfully transferred to E. coli, S. enteritidis, and K. pneumoniae isolates. These results broadened the knowledge regarding the predominance of tet(X4) mediated by plasmids.
In this study, tet(X4) was located in the IncFIA(HI1)-IncHI1A-IncHI1B(R27) hybrid plasmid (pSM2301-tetX4). This type of plasmid has been discovered in Klebsiella spp., Citrobacter spp., Leclercia spp., Citrobacter werkmanii, and E. coli, which were isolated from animals, pet dogs, wastewater from different regions in China, and so on (Lu et al., 2022). Xun et al. reported that 21 IncFIA-IncHI1A-IncHI1B hybrid plasmids carrying tet(X4) exhibited high similarity to pHNGS471-2 in K. pneumoniae in terms of coverage and identity (100% and 99.99%, respectively; Gao et al., 2022). In this study, hybrid plasmid pSM2301-tetX4 exhibited high identity with plasmids p3Z-5L-2-X4 in Klebsiella quasipneumoniae from swine, pE-T306-tetX4 from E. coli, and p10907_1 from S. enteritidis (Fig. 4). These results indicated that the spread of similar IncFIA-IncHI1A-IncHI1B plasmids has occurred among Enterobacteriaceae species, rendering plasmid an epidemic vector for the spread of tet(X4) conferring high-level resistance to TGC. Although tet(X4) mediated by plasmids is predominant in E. coli, it might further spread to Salmonella spp. and even to other species.
The transfer efficiencies of (10−7–10−6) pSM2301-tetX4 from SM2301 to three recipients (E. coli EC1905, K. pneumoniae KP2297, and S. enteritidis SM2211) were lower than previously reported (10−2–10−3; Lu et al., 2022; Sun et al., 2019). Genetic environmental analysis revealed that, unlike the previous finding, tet(X4) was located between two ISCR2 copies that form a circular structure. The genetic environments of tet(X4) can be categorized into four groups, of which one had a conserved structure, abh-tet(X4)-ISCR2-virD2-floR; the difference between them is the upstream region with different genes (ICSR2, ISCR2, and IS26; Yu et al., 2022). All genetic structures of tet(X4) have been reported to be associated with ISCR2, although they have various genetic contents of tet(X4) (Li et al., 2020; Lu et al., 2022; Yu et al., 2022). However, in this study, the upstream region was IS1R, which is contrary to the previous report. IS1R has been proven to promote transposition of tet(X4) from a suicide plasmid to an E. coli genome (Yu et al., 2022). These results indicate that ISIR-abh-tet(X4)-ISCR2-virD2-floR might be a new mechanism to spread tet(X4). According to the transmission mode of tet(X4) in bacteria based on the One Health approach, tet(X4) has the potential to spread across the food chain, community, and hospital, which will further cause untreatable infection. Therefore, it is recommended to further investigate tet(X4) and take control measures to limit its spread.
We herein report a tet(X4)-positive S. Rissen isolate mediated by a hybrid plasmid. The plasmid coharbored several types of resistant genes. We found that tet(X4) could be successfully transferred to E. coli, K. pneumoniae, and S. enteritidis and that the transconjugants acquired resistance to TGC; meanwhile, these transconjugants were resistant to all the tetracycline antibiotics tested in this study. Notably, the recipient carrying mcr-1 could obtain tet(X4) mediated by plasmid. The conjugative IncFIA(HI1)-IncHI1A-IncHI1B(R27) hybrid plasmid may further contribute to the coexistence of mcr-1.1 and tet(X4), which conferred resistance to “last-resort” antibiotics TGC and polymyxin. Therefore, collective efforts are urgently needed to preserve the potency of these essential antibiotics.
Footnotes
Authors’ Contributions
H.Z. and W.J. performed the research and draft the article. X.Y., Y.Y., Q.X., and T.Q. analyzed and interpreted these data. H.Z., H.U., and R.C. designed the study, supervised the whole project, and analyzed data. All authors reviewed, revised, and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Three-Year Initiative Plan for Strengthening Public Health System Construction in Shanghai (2023–2025) Key Discipline Project No. GWVI-11.1-09.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.