
Abstract
Salmonella enterica is one of the most common foodborne pathogens associated with the consumption of contaminated porcine, dairy, and avian products. Nontyphoidal Salmonella is a major cause of bacterial diarrhea, responsible for ∼150 million cases and 60,000 deaths annually. The main goal of this study was to determine the prevalence of Salmonella spp. and to establish the virulence profile (VP) from genes (avrA, invE, ssaD, sseF, ssaQ, ttrC) and plasmid genes (pefA, spvB, spvC) in isolates obtained from cheese, chicken, and pork sold in food markets in Barrancabermeja, Colombia. A survey was conducted on 100 samples each matrix. The detection of Salmonella spp. followed the ISO 6579:2017 standards modified, and isolates were confirmed using the invA gene. In addition, single polymerase chain reaction assays were developed to detect the nine virulence genes. Salmonella spp. was found in 62%, 32%, and 14% of pork, chicken, and cheese samples, respectively. A total of 277 isolates were biochemically, serologically, and molecularly compatible with Salmonella spp. The most representative serogroups were C and B. Forty-seven combinations of virulence gene were detected; 53.5% of the pork isolates, 46.2% of the cheese isolates, and 39% of the chicken isolates were distributed among VP1, VP2, and VP3 suggesting a higher pathogenic potential. In addition, seven isolates harbored plasmid-encoded virulence genes (spvB and spvC), which are associated with increased invasiveness. The results revealed a higher prevalence of Salmonella spp. in pork and chicken compared with other studies conducted in Colombia. The serogroups identified include serovars that more frequently affect humans Salmonella Enteriditis, Salmonella Newport, and Salmonella Typhimurium. The isolations have the majority of the virulence genes studied. These findings highlight the need to improve control measures and educate food handlers to minimize the presence of Salmonella spp. and its potential transmission.
Introduction
Salmonella enterica is a prevalent foodborne pathogen globally, with reservoirs in various animals like poultry, pigs, cattle, and reptiles. The primary transmission to humans is through contaminated food (Brown et al., 2021). Nontyphoidal Salmonella (NTS) is a major cause of bacterial diarrhea, responsible for about 150 million cases and 60,000 deaths annually (Centers for Disease Control and Prevention, 2024). Approximately 80.3 million of these cases are foodborne, highlighting the significant public health and economic burden (WHO and Food and Agriculture Organization of the United Nations, 2016). In Colombia, the Instituto Nacional de Salud reported 2727 NTS isolates between 2020 and 2023, associated with diarrheal disease, sepsis, and meningitis, underscoring its clinical significance (Instituto Nacional de Salud, 2024).
The pathogenesis of NTS involves multiple virulence and resistance genes throughout the genome, including Salmonella pathogenicity islands (SPIs) and mobile genetic elements. Some elements are conserved across Salmonella species, while others are specific to certain serovars. Virulence genes in SPIs (SPI-1 to SPI-21) play critical roles in salmonellosis, particularly those in SPI-1 and SPI-2. For example, the inv gene aids in adhesion and invasion, while the mgtC5 gene supports pathogen survival (Elkenany et al., 2019). Some virulent strains carry plasmid-encoded genes such as pef and spv, enhancing their ability to overcome host barriers and cause disease (Lan et al., 2018).
Determining Salmonella spp. prevalence and detecting their virulence profiles (VPs) is crucial for identifying characteristic isolates in a region and tracing contamination routes. This information is essential for designing strategies to reduce contamination along the food production chain (Donado et al., 2012). This study aimed to determine the prevalence of Salmonella spp. and establish the VP by identifying genes in SPI-1 (invA, avrA, invE) and SPI-2 (ssaD, sseF, ssaQ, ttrC), as well as plasmid genes (pefA, spvB, spvC) in isolates from cheese, chicken, and pork sold in food markets in Barrancabermeja, Colombia.
Materials and Methods
Sample collection
A total of 300 samples, comprising 100 specimens each of cheese, chicken, and pork meats were randomly collected from markets registered and authorized by the Secretaría Local de Salud of Barrancabermeja, Santander, Colombia, between October 2017 and June 2018. The sample size was determined with a 95% confidence level, 5% of absolute error, and an expected prevalence of 40.97% based on a previous study in Barrancabermeja (David et al, 2016). Each sample was packaged in sterile bags and transported in containers with ice to the microbiology laboratory. All samples were processed within 24 h of collection.
Isolation of Salmonella
The isolation of Salmonella spp. was conducted following ISO 6579:2017 standards modified. Each sample (25 g) was aseptically weighed, minced into small pieces, and transferred into a sterile flask containing of peptone-buffered water (Merck, Darmstadt, Germany). After incubation, the pre-enrichment was subcultured on Rappaport-Vassiliadis and tetrathionate broths (Merck, Darmstadt, Germany), and incubated at 41.5°C and 37°C, respectively, for 24 h. Each broth was then cultivated on Hektoen Enteric, xylose lysine deoxycholate, Salmonella–Shigella, and Bismuth Sulfite agar (Merck, Darmstadt, Germany) (Fig. 1).

Isolation of Salmonella spp. according to modified ISO 6579:2017 Standards.
Up to two typical or suspected colonies of presumptive Salmonella spp. were selected per agar and subcultured onto nutrient agar (Merck, Darmstadt, Germany) and incubated at 37°C for 24 h for further biochemical (urease, lysine iron agar, triple sugar iron, citrate sulfide indole motility, and methyl red/Voges–Proskauer) and serological identification by direct slide agglutination with Salmonella antiserum poly O (A-I & Vi), poly H (a-z) (Becton Dickinson, Sparks, MD) and with the Wellcolex™ Color Salmonella latex test (Remel Products, Lenexa, KS) for the presumptive identification of Salmonella serogroups A, B, C, D, E or G, and the Vi antigen.
DNA extraction
DNA was extracted from Salmonella spp. using a modified boiling method (Cavaco et al., 2016) by boiling the cells in 200 μl of sterile distilled water for 20 min, followed by centrifugation at 2,817 × g for 5 min. The supernatant served as the template for polymerase chain reaction (PCR).
Molecular identification
Primers targeting the invA gen (285 pb) were used to confirm the Salmonella genus. The isolates were evaluated for the presence of the invE, ssaD, sseF, spvB, spvC, avrA, ssaQ, ttrC, and pefA genes by using One Taq DNA polymerase M0480S (New England BioLabs, Ipswich, MA) for the first four genes, and Taq DNA polymerase (Thermo Scientific, Asheville, NC) for the remaining genes. PCR products were visualized by electrophoresis in a 1X TAE buffer on a 2% agarose gel. Primers, annealing temperature, and the size of each fragment are defined in Table 1. VPs were established based on the molecular identification of the nine genes mentioned earlier. Each gene was considered a qualitative variable (presence/absence). Salmonella enterica serovar Typhimurium ATCC 14028 was used as a positive control for all genes in the PCR assays. A small number of randomly selected isolates for each gene, as well as those amplified from the control strain, were sent for sequencing to Macrogen, Korea.
Primers Used in This Work for Each Virulence Gene and the Size of the Polymerase Chain Reaction Product
Statistical analysis
The prevalence of Salmonella spp. and 95% confidence intervals (95% CIs) were assessed using STATA software, v15 (StataCorp, College Station, TX). Subsequently, the relative frequency of each serogroup was determined as the proportion of positives from specific serogroups over the total isolates from each matrix, expressed as a percentage. Finally, the absolute and relative frequency of the genes and the VPs were determined.
Results
Prevalence of Salmonella spp.
From the 300 samples collected, 108 (36%) tested positive for Salmonella spp. The highest frequency of Salmonella spp. contamination was 62% (95% CI: 52.2–70.9) in pork, followed by 32% (95% CI: 23.0–42.1) in raw chicken, and 14% (95% CI: 8.5–22.1) in cheese. Of these samples, 277 isolates were compatible with Salmonella spp. according to biochemical, serological, and molecular results. The distribution of isolates in food matrices is found in Figure 2.

Salmonella spp. isolates obtained from cheese, chicken, and pork in food markets in Barrancabermeja, Colombia.
The 277 isolates were distributed into five serogroups based on their antigen (O). Out of 122 Salmonella spp. isolates from chicken, the most common groups were the C serogroup (59.8%) and the B serogroup (24.6%), with two isolates (1.6%) classified into the E or G group. In contrast, pork isolates were mainly classified in the E or G group (38.8%), while the frequencies of the C and B serogroups were 23.3% and 20.9%, respectively. The most prevalent serogroup identified in cheese was the E or G group (42.3%), followed by the D group (19.2%), with B and C each accounting for 3.8%. It is important to highlight that 47 isolates (16.9%) were not determined (ND), mainly in cheese due to limitations of the Wellcolex kit, and none were found to belong to the serogroup A or had the Vi antigen from these three matrices (Fig. 3).
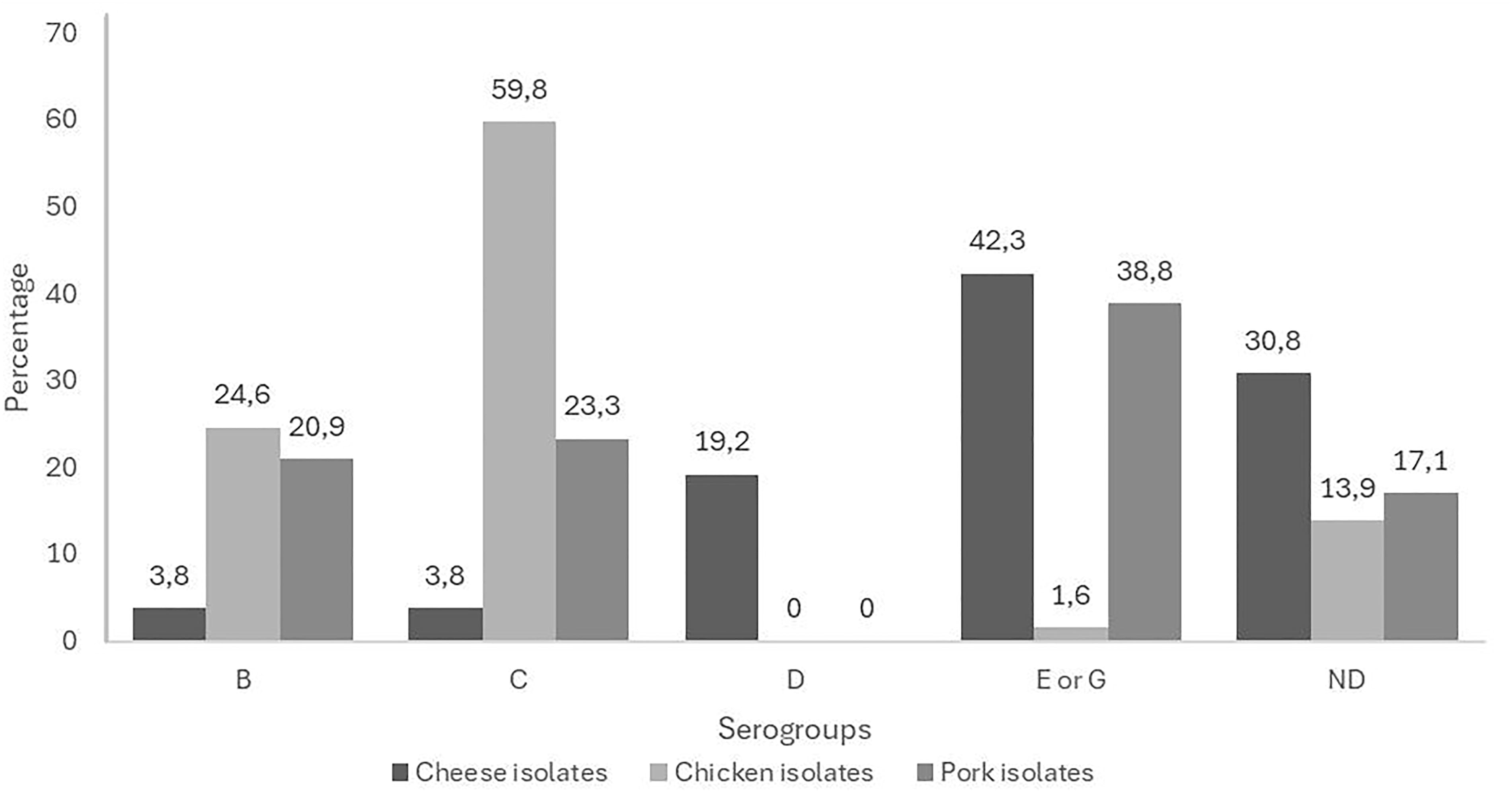
Percentage of serogroups identified by the Wellcolex™ Color Salmonella kit in Salmonella isolates from cheese, chicken, and pork meat.
Molecular confirmation and VPs
The invA gene was detected in 98.6% of chicken isolates and in 100% of pork and cheese isolates. Unconfirmed isolates were excluded. In total, 47 VPs were established, with 33 VPs identified in pork isolates, 31 VPs in chicken, and 16 VPs in cheese. Nevertheless, a large proportion of the isolates were VP1, VP2, and VP3, accounting for 53.5% of the VPs from pork, 46.2% from cheese, and 38.5% from chicken. Moreover, 50% of these isolates belonged to serogroup C (data not shown), suggesting a high level of similarity among the three matrices. The predominant VP in the matrices was VP1 (invA, invE, sseF, ssaD, ssaQ, ttrC, avrA), which was present in 33.3%, 23.8%, and 30.8% of the pork, chicken, and cheese isolates, respectively, the VPs detected are shown in Table 2. In addition, seven pork, one chicken, and two cheese isolates had at least one of the three plasmid genes evaluated (Table 3).
Virotype Frequency in the Three Food Matrices
Characteristics of Salmonella Isolates with spvB, spvC, and pefA
Identification.
Virulence profile.
Discussion
Salmonellosis is a disease that occurs in humans and is associated with the consumption of contaminated foods, particularly chicken. However, in this study conducted in the city of Barrancabermeja, the highest prevalence of Salmonella spp. was found in pork (62%), followed by chicken (32%) and cheese (14%). The prevalence of Salmonella spp. in pork in this study is the highest reported in Colombia to date, being nearly twice as high as the prevalence observed in Bogotá (37.8%) and almost five times higher than those reported in Montería (13%), Cartagena (6.3%), and Tolima (3.3%) (Arcos et al., 2013; Espinal et al., 2006; Mejía and Arbelez, 2007). Compared with other countries, the prevalence is higher than that reported in Latin American countries such as Mexico (22.5%) and Argentina (24.1%) (Colello et al., 2018; Villalpando et al., 2017), but lower than those observed in China (73.1%) and Ireland (69%) (Duggan et al., 2010; Zhang et al., 2018). These data highlight the contamination in these types of foods, which increases the risk of human infection and underscores the need for enhanced supervision and monitoring of food, particularly pork, in warm climates such as the tropics (Zhang et al., 2018).
The prevalence of Salmonella spp. detected in chicken was similar to that found in other studies in Santander (36.7%), Putumayo (33.3%), and Atlántico (33.3%), and higher than that in Bogotá (26%) (Donado et al., 2012). This is also comparable with studies in Guatemala (34.3%), Iran (33%), Belgium (36.5%), and Spain (35%) (Fardsanei et al., 2018; Jarquin et al., 2015). However, it is lower than in many developing countries, including Mexico (47.7%), Korea (42%), Vietnam (45.9%), Egypt (44%), China (63.6%), and Cambodia (88.2%) (Ahmed et al., 2017; Choi et al., 2015; Ta et al., 2014; Villalpando et al., 2017; Zhang et al., 2018).
The similarities in these prevalence rates may be attributed to the fact that, in most countries, Salmonella spp. control and monitoring systems primarily focus on poultry. In addition, the samples analyzed were collected from a poultry-producing region, such as Santander, where heightened awareness and stricter measures are likely implemented by poultry farmers to minimize Salmonella spp. contamination. However, the percentage observed still indicates a latent risk of human infection, highlighting the need for more rigorous management strategies throughout the entire poultry production chain, especially in traditional farming practices (Carrasco et al., 2012).
Regarding the cheese matrix, the prevalence was 14%, significantly lower than the 41% reported in a study using the enzyme-linked immunosorbent assay technique (David et al., 2016). However, it is higher than those reported in other Colombian cities such as Barranquilla (7.9%), Montería (5.3%), and Paipa (0%) (Castellanos et al., 2021; Durango et al., 2004). When compared with other countries, it is lower than the prevalence found in Cuba (17.9%) in artisanal fresh cheese, and in Mexico in panela cheese (34%) and adobera cheese (20%) (Martinez et al., 2020; Torres et al., 2012).
It is important to note that differences in methodology, sample size, cheese types, and geographical regions with varying climates and cultures complicate data comparison in the mentioned prevalence studies. Nevertheless, this study underscores the need to address Salmonella spp. prevalence in the production chain, conduct continuous surveillance, and engage producers and farmers in contamination prevention to minimize risk.
Monitoring sources beyond chicken is essential, as Salmonella spp. can also be present in contaminated vegetables and fruits (Ehuwa et al., 2021). Therefore, prioritizing quality processes on farms and disseminating policies that promote good food processing practices is imperative. This includes fostering a stronger sanitary culture among producers and improving food safety standards, especially in the pork industry (United States Department of Agriculture, 2020).
The identification of Salmonella serogroups plays a crucial role in understanding their distribution and potential sources and would facilitate the control and monitoring of this pathogen in food matrices. For instance, by 2024, nearly 49 new outbreaks have been reported in the United States, with most being caused by Salmonella Enteritidis (https://www.cdc.gov/ncezid/dfwed/BEAM-dashboard.html). However, reports on Salmonella spp. by serogroups are limited and are more commonly described by serovars. In this study, the most prevalent Salmonella spp. serogroups were C (29%) and E or G (27.3%), the latter of which could not be distinguished using the “Wellcolex Color Salmonella” kit used in this study. It should be noted that 20.3% of the isolates could not be classified into a serogroup. In addition, serogroup D1 was only found in the cheese matrix.
In pork, the most prevalent serogroups were E or G (38.8%), followed by C (23%) and B (20%). In contrast, in China, the highest prevalence was serogroup B (Salmonella Derby and Salmonella Typhimurium), followed by serogroup E (Salmonella London), with a prevalence for the latter group ranging from 1.25% to 23.08%, depending on the geographic location (Shen et al., 2022).
For the chicken matrix, the most prevalent Salmonella spp. serogroups in this study were C (59.8%) and B (24.6%), similar to what was found in the United States, where serogroup C was the most common, representing 25.7% of all reported cases, followed by serogroup B (20.5%). Conversely, in Europe, by 2012, 42% of cases were attributed to serogroup D, followed by serogroup B. This is like the situation in Africa, where serogroup D is the most prevalent (44.6%), followed by serogroup B (30.9%). Similarly, according to data collected in Asia, serogroup D is the most common (45.4%), followed by serogroup C (27.3%) (Fuche et al., 2016).
In Colombia, in 2003, six isolates belonging to serogroups D and B were reported in broiler chickens. In 2005, a study reported a Salmonella spp. positivity rate of 2.75%, with serogroup B being the most prevalent. Similarly, in 2009, in Santander, Botero reported a positivity rate of 12.94% in broiler chickens, with all isolates belonging to serogroup B. Although the number of studies is limited and not directly comparable with this research, it is noteworthy that serogroup B has been frequently reported in the country, similar to what was found in this study (Ministerio de la Protección Social and Instituto Nacional de Salud, 2011).
On the other hand, it is important to highlight that most of the cheese matrix isolates were not classified into any of the groups detected by the kit, indicating that the Salmonella spp. isolates belong to serogroups different from those detected by the test. The literature reports cheese outbreaks caused by common Salmonella serovars (Gould et al., 2014; Ung et al., 2019), such as Salmonella Enteritidis outbreak in the Marche region in the 2020 (Central Italy) due to the consumption of pecorino cheese made from raw sheep milk (Napoleoni et al., 2021); the outbreak of Salmonella Newport linked to Oaxaca soft cheese from Mexico in 2018 (Plumb et al., 2019); and the foodborne outbreak involving Saint-Nectaire cheese contaminated with S. Dublin in France in 2012 (Vignaud et al., 2017).
It is worth noting that finding isolates of Salmonella spp. belonging to serogroups B and D1 in any food allows us to infer the possibility of finding serotypes with high transmission potential to humans because they include species such as Salmonella Typhimurium and Salmonella Enteritidis, which have been recognized as the most common serotypes associated with nearly 60% of human salmonellosis cases reported in Colombia over the past 20 years (Instituto Nacional de Salud, 2019). One of the limitations of this study is the lack of serovars identification as well as the differentiation of serogroups E or G.
It should be emphasized that without complete serotyping of all isolates obtained, it is not possible to determine and evaluate the microbial risks for consumers. In addition, no evidence was found for the presence of the Vi antigen, suggesting the absence of invasive serotypes such as Salmonella Typhi, Salmonella Dublin, Salmonella Paratyphi A, and C. Prioritizing the implementation of standardized methodologies that allow the unification of criteria for the microbiological analysis of these foods is essential. Implementing these techniques in public health laboratories in different regions of the country is necessary to ensure the traceability and control of Salmonella spp. in food matrices, as well as to monitor antimicrobial resistance (Marshall et al., 2024).
On the other hand, the predominant VP in food matrices was VP1, which comprises all the analyzed genes and implies a greater invasive capacity of the bacteria. The analyzed genes are part of two type three secretion systems encoded in SPI-1 and SPI-2, which are involved in the translocation of proteins to penetrate the intestinal epithelial cells and allow the bacteria to survive intracellularly within the host during the systemic phase of the disease (Hensel, 2004; Kuhle et al., 2006). The results suggest that isolates presenting VP1 have a higher pathogenic potential because they possess the genetic elements necessary to invade human cells. The variations observed compared with other VPs could be explained by the loss or acquisition of virulence gene clusters that contribute to a particular virulence phenotype, which manifests at a specific stage of infection. Therefore, this potential could increase a microorganism’s pathogenicity (Hensel, 2004; Marcus et al., 2000).
The spv genes encoded by plasmids are in a highly conserved homologous region, which has only been found in some serotypes of Salmonella spp. belonging to subspecies I of Salmonella enterica, particularly those showing host adaptation, such as Salmonella Typhimurium, Salmonella Enteritidis, Salmonella Choleraesuis, Salmonella Arizona, and Salmonella Dublin (Guiney and Fierer, 2011 (Guiney and Fierer, 2011; Marcus et al., 2000). In this study, seven isolates presented virulence genes in the spv locus, of which two isolates belonging to VP1 contained the plasmid genes spvB and spvC. This could indicate a higher potential for virulence and invasion. However, further studies are needed to quantify their virulence and their risk of causing diseases in humans.
Finally, the results of this study are significant as they identify Salmonella enterica prevalence in different food matrices and the serogroups present. Although not providing complete serotyping, this approximation aids in improving food safety systems, especially for animal products such as pork, chicken, and cheese.
Conclusions
This study determined the presence of Salmonella spp. from the food markets in Barrancabermeja, Colombia. Among the analyzed matrices, the highest prevalence was found in pork meat (62%). In addition, the serogroup C was predominant, which includes serotypes associated with zoonoses. Moreover, the majority of isolates exhibited VP1, characterized by the presence of all analyzed genes, and only seven isolates harbored the spv gene, suggesting greater potential for virulence. Consequently, further studies are considered necessary to evaluate the virulence potential of these isolates more thoroughly.
Footnotes
Acknowledgments
The authors gratefully acknowledge the undergraduate students from UNIPAZ and UIS who actively participated during the research, to Dr. Daniel Svozil and Laura Padilla for the support during writing of this article and to everyone who in one way or another, both administratively or technically, helped to make this possible.
Authors’ Contributions
S.V.A.N.: Investigation, formal analysis, writing—original draft preparation. A.D.C.: Conceptualization, investigation. C.I.G.: Conceptualization, supervision. G.R.C.: Conceptualization, supervision. M.N.R.: Conceptualization. E.M.P.C.: Investigation. M.M.: Investigation. N.F.O.S.: Formal analysis. R.A.M.V.: Formal analysis, supervision. All authors contributed to the article and approved the submitted version.
Ethical Approval
This study was approved by the Ethics Committee of the UIS (24/8/2018).
Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was supported by Ministerio de Ciencia y Tecnología (MinCiencias) (grant number 1102-74455994); Universidad Industrial de Santander, Instituto Universitario de la Paz and Secretaría Local de Salud.
Supplementary Material
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.