
Abstract
Rats are rodents commonly found in Thailand that carry various zoonotic pathogens. Bacterial zoonosis can occur in a shared environment between humans and rats, especially in human communities and agricultural areas. Escherichia coli, particularly pathogenic and multidrug-resistant strains, is a significant public health concern that is transmitted by rats. This study aimed to investigate the antibiotic resistance (ABR) and biofilm formation of E. coli in caught rodents from Nakhon Si Thammarat province, Thailand. Captured rats were dissected to collect intestinal content for E. coli isolation. Two hundred and two confirmed E. coli were subjected for pathotype identification, antibiotic susceptibility testing, biofilm-forming ability (BFA), and the presence of related genes. Two E. coli isolates from intestinal content samples were atypical enteropathogenic (aEPEC). Predominantly, 52.97% of E. coli had azithromycin resistance, which was harbored by 35.64% of captured rats. Multidrug resistance (MDR) was found in 12.38% of E. coli isolates with 17 different MDR patterns. Remarkably, 96% of MDR isolates were resistant to azithromycin. Most E. coli harbored ereA (52%), followed by the bla TEM and aacC2 genes (6.44% each). Approximately 87% of isolated E. coli revealed moderate-to-high BFA. Predominantly, moderate-to-strong biofilm-forming E. coli harbored pgaA and pgaC genes. aEPEC, azithromycin resistance, MDR, and moderate-to-strong formation were the aspects of concern. Furthermore, the study of antibiotic-resistant E. coli in rats should be performed, particularly in terms of the transmission pathway, and the application of rats as bioindicators for ABR surveillance in Thailand should be established.
Introduction
Rats are small mammals primarily found in Thailand which carry several zoonotic pathogens through their attacks and food contamination (Rabiee et al., 2018). Rodent-borne diseases can arise in the shared environment between humans and rats (Tanthanathipchai et al., 2023). Rats and humans share a crucial environment, including human communities and agricultural areas (Moradpour et al., 2018). Rattus rattus complex group, containing R. norvegicus and R. exulans, is the primary species associated with human settlements in Thailand (Herbreteau et al., 2011).
Enterobacteriaceae bacteria are typically found in rats (Desvars-Larrive et al., 2019; Pormohammad et al., 2019). Pathogenic Escherichia coli (E. coli) strains are harmful and considered potential sources of human infection (Malik and Memona, 2010). Seven pathotypes have been identified, including enteropathogenic E. coli (EPEC), enterohaemorrhagic E. coli, enterotoxigenic E. coli, enteroinvasive E. coli, enteroaggregative E. coli, diffusely adherent E. coli, and adherent-invasive E. coli (Pakbin et al., 2021). E. coli is a food sanitation indicator and fecal contamination. Nowadays, antibiotic resistance (ABR) E. coli is concerned. Interestingly, E. coli is also a significant horizontal gene transferer, allowing normal bacteria in rats to receive ABR genes from several sources and serve as reservoirs for resistant E. coli in the environment.
ABR is a global issue linked to the One Health concept, particularly in the context of human–animal environment interaction (Aslam et al., 2021). Animals are exposed to ABR bacteria from the environment, becoming reservoirs of these bacteria. Furthermore, bacterial biofilm formation can lead to ABR and help bacteria survive in a high-dose antibiotic environment (Rather et al., 2021). Multidrug resistance (MDR) in E. coli is a significant public health concern due to its ability to resist multiple classes of antibiotics (van Duin and Paterson, 2020).
Several countries, including Thailand, are increasingly concerned regarding MDR-E. coli. Mostly, beta-lactams, fluoroquinolones, and aminoglycosides are antibiotics that are reported to have resistance in E. coli (Urban-Chmiel et al., 2022). Rodents carry both commensal and pathogenic E. coli, isolated from their intestinal tract, reportedly indicating MDR (Azimi et al., 2021). Interestingly, the study on the ABR and biofilm-forming ability (BFA) of E. coli in Thai rodents is limited. Hence, the study aimed to determine the ABR and BFA of E. coli from rodents in Nakhon Si Thammarat (NST), Thailand.
Materials and Methods
Ethical considerations
The experiments involving animals were conducted in accordance with protocols of the CERoPath project (Herbreteau et al., 2011). Experimental procedures were approved by the Walailak University Institutional Animal Care and Use Committee (WU-ACUC-65034 and WU-ACUC-65076) and the Walailak University Institutional Biosafety Committee (WU-IBC-66–022).
Sample size calculation and rat capture
In total, 101 rat samples were calculated using a formula for estimating an infinite population proportion (https://epitools.ausvet.com.au/oneproportion) based on previous prevalence (Azimi et al., 2021), 10% desired precision, and a 95% confidence interval. Rodent live traps were placed in rural areas of NST, including residences, veterinary hospitals, research buildings, and food courts, from 5:00 p.m. to 6:00 a.m. in three subdistricts (Tha Sala, Thai Buri, and Hua Taphan) of Tha Sala district. Hua Taphan has mostly agricultural fields, especially rubber plantations, while other subdistricts have mostly residences and work buildings.
Rat identification
Rats were euthanized using carbon dioxide inhalation following the CERoPath guideline (Herbreteau et al., 2011). Euthanized rats were recorded for their hair color, body marks, and weight. Five length parameters, including head-body, tail, hind-foot, ear, and skull, were measured. The genitalia and reproductive organs were examined for sex determination. The genus of rats was determined using the mentioned data, following the CERoPath guideline (Herbreteau et al., 2011).
Sample collection and bacterial isolation
Rat carcass was placed on a sterilized tray. The abdominal area was disinfected using 75% ethanol before dissection. Duodenum, jejunum, ileum, caecum, and large intestine were cut and put in sterile buffered peptone water and incubated at 37°C for 24 h for enrichment. The enriched sample was diluted 10-fold, spread on eosin-methylene blue agar, and incubated at 37°C for 24 h. Two purple colonies with green metallic sheen were identified as suspected E. coli colonies. The suspected E. coli colonies were used for determination of ABR, BFA, and DNA extraction.
Polymerase chain reaction
E. coli-suspected colonies were extracted genomic DNA using the boiling method with some modification (Peng et al., 2013). Pathotype identification was performed by a thermal cycler (Eppendorf Vapo Protect, Germany). Polymerase chain reaction (PCR) reactions contained 0.2 mM of dNTPs each (ABM®, Canada), 10X Taq PCR buffer (ExcelTaq TM series, Taiwan), 1 µM of forward and reverse primers (Macrogen®, Korea), and 5 U/µL Taq DNA polymerase (ExcelTaq TM series, Taiwan). In total, 3 µL of extracted DNA was used for the template, and distilled water was added up to 25 µL for each reaction. Primers and PCR conditions are detailed in Supplementary Table S1. The PCR product was run on a 1.5% agarose gel, and DNA bands were observed using a UV illuminator (ChemiDocTM, USA). The suspected DNA band was purified using a DNA purification kit (BioFACTTM Gel & PCR Purification System, Korea) and analyzed by Sanger’s sequencing (Marcrogen®, South Korea).
Antibiotic susceptibility and detection of ABR gene
E. coli-confirmed colonies cultured in Mueller–Hinton broth (HiMedia®, India) were diluted to a turbidity of 0.5 McFarland and spread on Mueller–Hinton agar (HiMedia®, India). Twelve antibiotic discs (Supplementary Table S2) were placed on the plate and incubated at 37°C for 16–18 h. E. coli ATCC 25922 was used as the reference strain. The inhibition zone was measured and interpreted following the Clinical and Laboratory Standards Institute recommendation (Clinical and Laboratory Standards Institute [CLSI], 2021). Interpretation was categorized into susceptible (S), intermediate (I), and resistant (R). MDR was defined as an isolate showing phenotypic resistance to at least three classes of antibiotics, and the multiple antibiotic resistance (MAR) index of each MDR isolate is calculated (Krumperman, 1983). Detection of ABR genes carried by isolated E. coli was performed using the PCR condition in Supplementary Table S1.
Detection of BFA and biofilm-related genes
E. coli was cultured overnight in trypticase soya broth (HiMedia®, India) supplemented with 1% dextrose monohydrate (Loba Chemie™, India), adjusting to a density of 1 × 106 cells/mL. In total, 200 µL of adjusted cultured medium were cultured in a 96-well plate and incubated at 37°C for 24 h. E. coli ATCC 25922 and sterile tested medium were used as a positive and negative control, respectively. The producing biofilm quantity was measured by comparing biofilm mass to a negative control. The biofilm biomass was stained with 0.1% crystal violet and dissolved in 100% dimethyl sulfoxide after washing with 1X PBS. The isolates with higher dye concentrations were analyzed for optical density, indicating higher biofilm biomass production. The optical density at 590 nm was measured using a microplate reader (AccuReader M965 Plus, Taiwan). The BFA was interpreted following the previous study (Kulnanan et al., 2021) as having no, weak, moderate, or strong abilities. Biofilm-related genes were detected using PCR using the primers (Supplementary Table S1).
Multi-locus sequence typing
MDR with strong biofilm-forming isolates was analyzed using the Achtman-7 gene multi-locus sequence typing (MLST) scheme for further typing. Seven reference genes, including adk, fumC, gyrB, icd, mdh, purA, and recA, were detected and sequenced. The MLST-determination PCR protocol (Supplementary Table S1) was based on the protocols of a previous study (Wirth et al., 2006).
Statistical and bioinformatic analysis
The statistical analyses were conducted using the R program version 4.2.0 (R Core Team, 2020). The data was analyzed using descriptive statistics, and interesting risk factors were tested using Fisher’s exact test or the chi-square test. The analyses were conducted with a 95% confidence interval, and a p value <0.05 was considered significant. The DNA chromatogram was edited using SnapGene® Viewer version 5.3.2 (https://www.snapgene.com/snapgene-viewer). Then, the BLAST program was performed to analyze the matching of bacterial species and protein type (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The MLST analysis involved searching for the best-matched DNA sequence to the reference allele of each reference gene to define the sequence type (ST). Multiple alignment among edited sequences and the best-matched allele sequences of ST was performed under the CLUSTAL-W using MEGA-11 (https://www.megasoftware.net). The complete aligned sequences of each gene were concatenated and used for a maximum likelihood phylogenetic tree under the bootstrapping method with a fitted nucleotide substitution model using MEGA-11. In addition, the concatenated sequences were used for split decomposition tree construction using SplitsTree 6 (https://uni-tuebingen.de).
Results
In this study, rats were identified based on morphometric measurement and habitat-related factors (Table 1). In total, 202 E. coli was isolated from intestinal content samples of 101 captured Rattus spp. (two isolates/rat). Pathotype identification based on molecular detection revealed that two isolates (0.99%) were EPEC but only had the eae gene, which was identified as atypical enteropathogenic (aEPEC) (Table 1). Interestingly, 52.97% of isolates were resistant to azithromycin (Fig. 1A), and 35.64% of intestinal content samples contained azithromycin-resistance E. coli (AREC). Remarkably, 24 of 25 (96%) MDR isolates were AREC. In addition, 25/202 (12.38%) of isolates were MDR, and 64.85% of isolates (50.50% of rats) resisted at least one tested antibiotic drug. Overall, 17 different MDR patterns were found in MDR isolates (average MAR index of 42.65%), with CZ-CIP-CF-AZM being the most prevalent. One MDR isolate had the highest MAR index, with patterns of AMP-C-COT-TE-TR-CZ-AZM (6.90%; 95% confidence interval [CI] = 0.85–22.77%) (Fig. 1B).
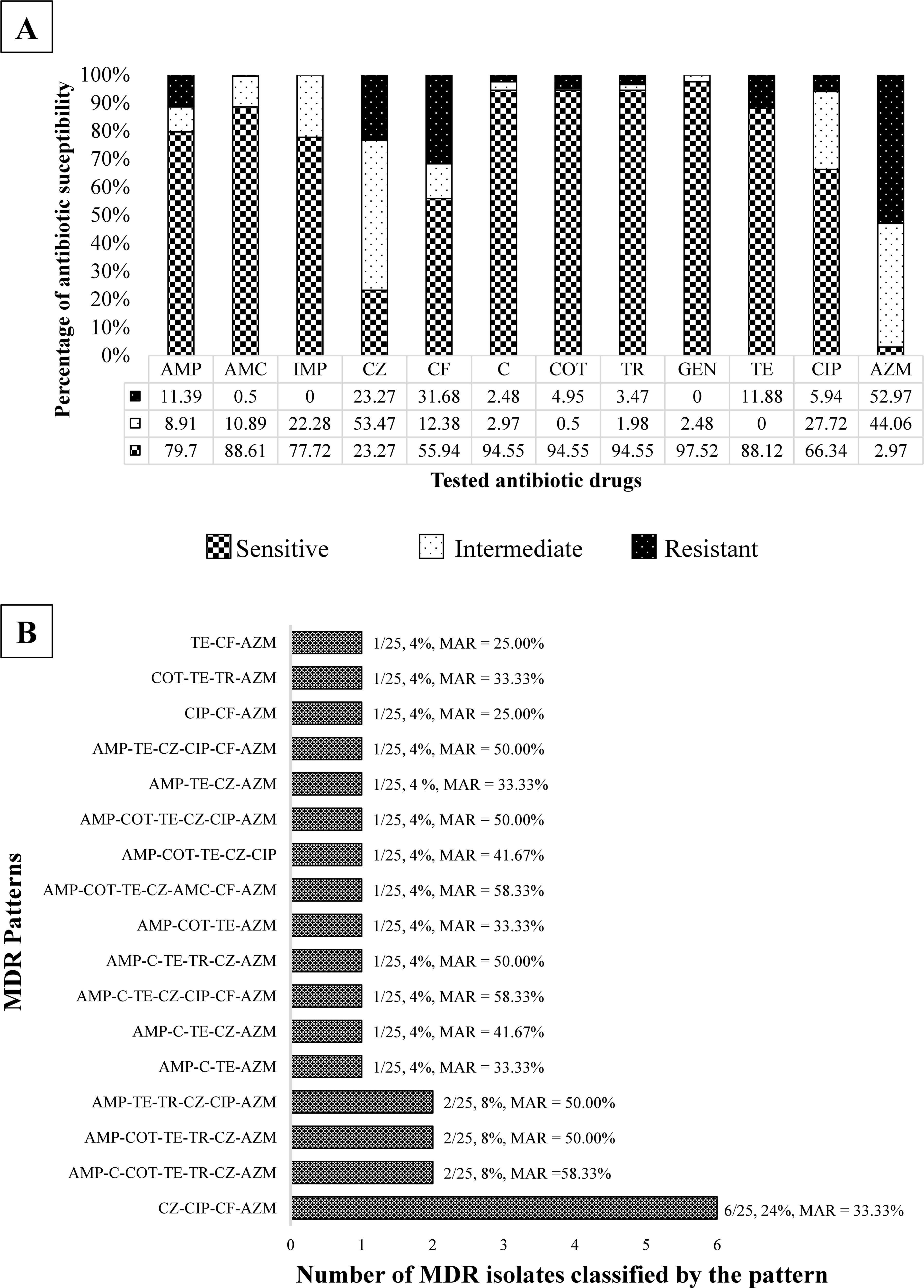
E. coli isolates proportion classified by antibiotic susceptibility and tested antibiotic drugs
Characteristics of Studied Rats and Identification of E. coli Isolated from Captured Rats
Morphometric parameters of 101 captured rats.
Count and percentage of 101 captured rats.
Count and percentage of 101 captured rats and number in [ ] is number of E. coli isolate
E. coli, Escherichia coli; EPEC, enteropathogenic; NF, not found.
The most phenotypic resistance in E. coli in this study was beta-lactam group. Among beta-lactam-related genes, 6.44% (95% CI = 3.47–10.75%) harbored bla TEM, 3.47% (95% CI = 1.40–7.01%) harbored bla SHV, and 0.50% (95% CI = 0.01–2.73%) harbored bla CTX-M (Fig. 2A). In addition, 13 of 202 isolates carried the aacC2 gene; however, no isolates presented phenotypic resistance to gentamicin. Interestingly, the most prevalent ABR gene in this study was the macrolide resistance gene, which is related to the phenotypic resistance to azithromycin (Fig. 2B), and the ereA gene was found to be present in 107/202 (52.97%) of isolates. Of this, 61 of 107 azithromycin-resistant isolates (57.01%; 95% CI = 47.08–66.54%) carried the ereA gene, which was found in 18 intestinal content samples (17.82%, 95% CI = 10.92–26.70%).
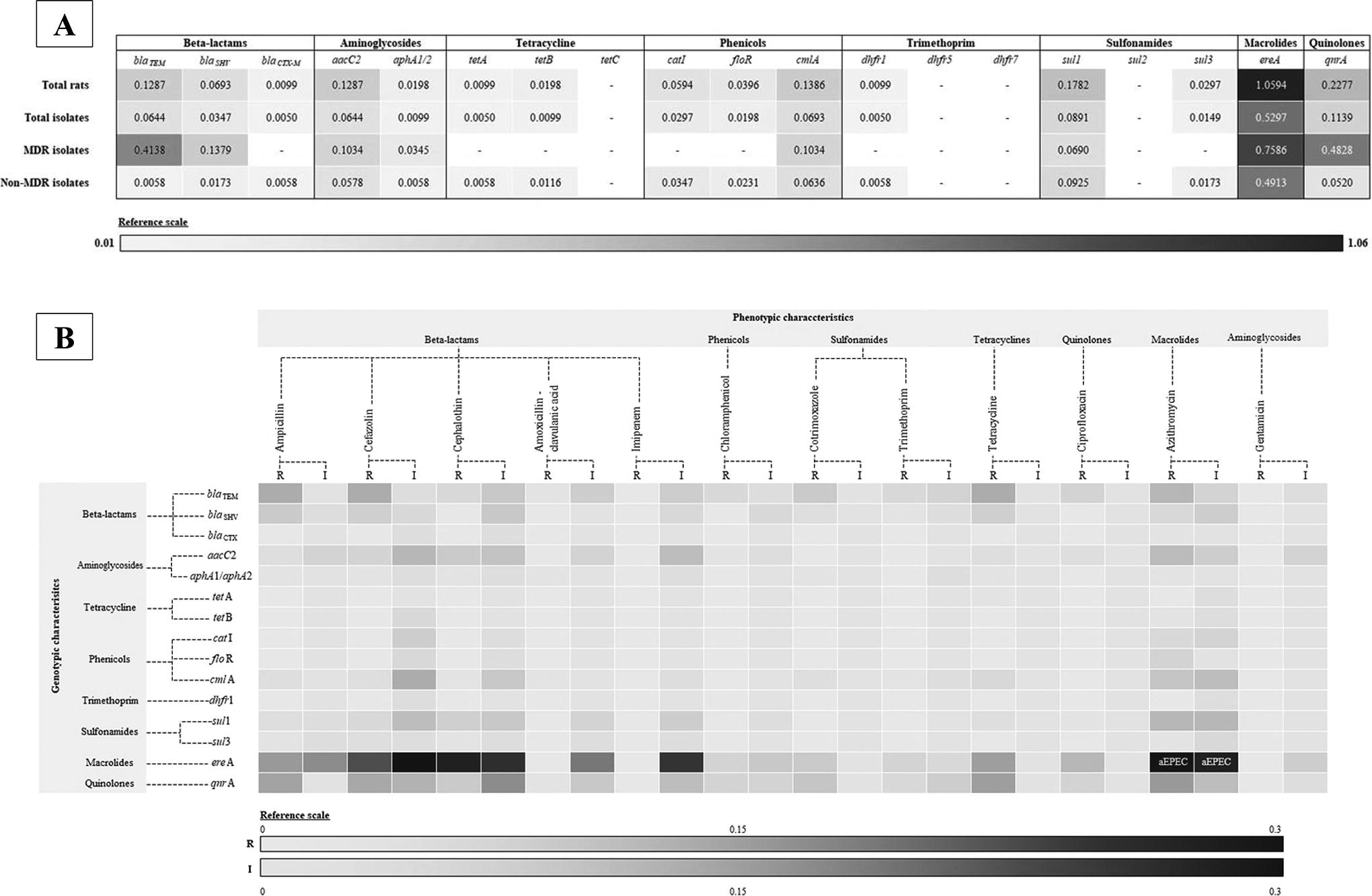
Heat map of rate of detected antibiotic resistance genes of isolated Escherichia coli (E. coli) from the intestinal tract of captured rats classified by different characteristics
For BFA, 88 of 179 (49.16%, 95% CI = 41.63–56.73%) had moderate BFA, followed by strong BFA (38.55%, 95% CI = 31.38–46.10%). Moreover, the proportion of BFA between MDR isolates (82.61%) and non-MDR isolates (88.46%) was similarly closed, and there was no association between the BFA and MDR characteristics (p = 0.49). Interestingly, 49.16% of AREC revealed a high rate of moderate-to-strong BFA (Table 2). Moderate-to-strong biofilm-forming isolates mostly carried both pgaA and pgaC genes (Table 3), however no association between the BFA and the presence of the pga gene was found (p = 0.89). Six isolates (2.97%) were MDR with strong BFA, which were mainly grouped into four clades and identified through a concatenated phylogenetic tree analysis, with one isolate being primarily dissociated from other reference E. coli (Fig. 3). The split decomposition tree, as shown in Figure 1, presents that out of the concatenated sequences of all reference genes, three clades were groups, and two isolates drifted apart. From molecular epidemiology, six isolates of E. coli in this study had the sequence of their reference genes closely similar to the reference genes of other disease-causing E. coli found in various infected hosts, including humans, domestic animals, wildlife, and various countries.
Number of Isolated E. coli from the Intestinal Tract of Captured Rats Classified by Biofilm-Forming Ability Group and Type of Resistance *
Number of tested E. coli for biofilm forming ability was 179 isolates.
Using crystal violet assay.
AMC, amoxicillin with clavulanic acid; AMP, ampicillin; AZM, azithromycin; C, chloramphenicol; CF, cephalothin; CIP, ciprofloxacin; COT, co-trimoxazole; CZ, cefazolin; E. coli, Escherichia coli; GEN, gentamicin; IMP, imipenem; MDR, multidrug resistance; TE, tetracycline; TR, trimethoprim.
Number of Escherichia coli (E. coli) Isolates Classified by Biofilm-Forming Ability, Biofilm-Formation-Related Gene, and Multidrug Resistance of Isolated E. coli from the Intestinal Tract of Captured Rats

Concatenated phylogenetic tree and split decomposition tree of six MDR-E. coli with strong biofilm formation. E. coli, Escherichia coli; MDR, Multidrug resistance.
Discussion
Rats are reported to be a reservoir of MDR-E. coli, ABR-E. coli, and EPEC. (Azimi et al., 2021). Moreover, rats are small mammals that live in close proximity to human settlements, induced likelihood of disease transmission from rats to humans (Tanthanathipchai et al., 2023; Thaikoed et al., 2024). Rattus species, particularly R. rattus, are prevalent in Southeast Asia and are known to transmit various foodborne pathogens, such as E. coli (Jahan et al., 2021). EPEC are categorized into typical EPEC and aEPEC, based on the presence of the bpfA gene. (Hernandes et al., 2009; Yamani and Elhadi, 2022). aEPEC is mentioned as zoonosis, which causes watery diarrhea in infants and children (Kaur and Dudeja, 2023), leading to financial loss and parental absence. In the studied area, lifestyle and poor sanitation of people increased the likelihood of receiving aEPEC. Of this, rats might play a role as a possible EPEC reservoir and transmitter in part of the whole transmission chain of an EPEC-causing disease. Even if the prevalence of aEPEC in intestinal content samples of rats in this study is low, the infections on infants and children are affected, particularly in the family in unwell sanitation on food, water, and milk.
ABR in rodents is increasing (Gwenzi et al., 2021), with rats being reported as a source of reservoirs (Desvars-Larrive et al., 2019). Rats have been identified as potential carriers of ABR to humans through food chain contamination and direct contact (Gwenzi et al., 2021). MDR proportion in this study was mostly different from several studies elsewhere in Asia (Azimi et al., 2021; Huy et al., 2021; LE Huy et al., 2020). The main factors influencing the ABR rate variation across regions encompass disparities in regulations governing antimicrobial usage, which impact the availability of specific antimicrobial agents; spatiotemporal variations that could affect bacterial adaptability; different ABR surveillance systems; distinct social cultures influencing hygiene practices; and variations in bacterial genetics. Several studies indicated that rats are implicated in the transfer of ABR determinants between animals, humans, and environmental sources, and rats pose a risk of storing transferable MDR-E. coli (Dominguez et al., 2023; LE Huy et al., 2020). Furthermore, urban rats, often exposed to contaminated environments, are potential reservoirs and transmitters of ABR bacteria in the chain of the human–animal environment interface (Sano et al., 2023). Of this, rats might play an important role in MDR-E. coli transmission through food chain contamination.
Southern Thailand is the main region that enhances aquaculture and meat production for export, and over the past decade, ABR (particularly beta-lactams) was mostly mentioned in E. coli, especially for bla gene carriers (Nhung et al., 2016). NST is a province that contains aquaculture, livestock farms, fresh markets, backyard farms, and local communities. This study filled the gap of the study in agriculture fields, human settlement, and working buildings. Interestingly, the mentioned places were reported to have both ABR- and MDR-E. coli, including vegetables and meats in markets (Romyasamit et al., 2021; Sornsenee et al., 2022), pig farms (Boripun et al., 2023; Mitsuwan et al., 2023), backyard farms (Wongtawan et al., 2022), and wastewater from community hospitals (Siri et al., 2024). Use of antibiotics in community hospitals in the studied area (mostly beta-lactams and quinolones) was reported to induce resistant bacteria to these drug groups, which were found in the wastewater of the hospital (Siri et al., 2024). Moreover, the dynamic transmission of ABR-E. coli among these mentioned places should be additionally studied.
A high rate of AREC was found in this study, which had a different rate from the previous studies elsewhere (Azimi et al., 2021; Himsworth et al., 2015; Uea-Anuwong et al., 2024). Azithromycin is an antibiotic drug that is widely used in Thailand, both in human and veterinary medicine. Especially in animals, the use of azithromycin is hardly controlled both in livestock and companion animals, and this might lead to AREC. Azithromycin resistance is primarily caused by overexpression of the efflux pump, mutation of the 23S ribosomal protein, methylation, esterase inactivation, and phosphotransferase (Gomes et al., 2017). In addition, Enterobacteriaceae bacteria possess a hydrolytic enzyme encoded by the ereA gene, which is crucial for macrolide hydrolysis (Xing-Wei et al., 2024; Zieliński et al., 2021) and leads to macrolide resistance, especially for erythromycin and azithromycin (Paul et al., 2023; Varela et al., 2021).
Moreover, cephalothin and cefazolin were found to be resistant next to azithromycin in this study. The reported resistance E. coli in NST was also carried ABR gene, with the highest resistance rate found in bla family genes, particularly bla TEM (Boripun et al., 2023; Mitsuwan et al., 2023; Romyasamit et al., 2021; Sornsenee et al., 2022; Wongtawan et al., 2022). This study found that beta-lactam antibiotics had the highest resistance rate, and bla TEM, bla SHV, and bla CTX-M genes were found in E. coli isolated from rats. Similarly, another study in the studied area reported the presence of the same bla gene group, particularly bla TEM, in patients (Romyasamit et al., 2024), retail markets (Romyasamit et al., 2021), pig farms (Mitsuwan et al., 2023), poultry farms (Thomrongsuwannakij et al., 2022), soil, fighting bulls, sport horses, and fighting cocks (Wongtawan et al., 2022). This reflects that E. coli in rodents might play an important role as a reservoir for beta-lactam resistance-related genes in the studied area.
Most isolates were found to produce biofilm biomass in this study; however, the correlations between high-to-moderate BFA and ABR were not significant. Biofilm formation is only one ABR mechanism; however, there are any other mechanisms that bacteria might perform such as efflux pumps, enzymatic degradation, and mutations of drug binding sites. Furthermore, biofilm formation frequently induced ABR is a virulence mechanism, particularly in environmental setting. Thus, biofilm might help ABR bacteria to survive in the environment before being received by hosts such as humans and animals. Then, ABR with FBA bacteria was received by rats and exchanged with the ABR gene inside the host’s intestine environment.
Conclusion
This study emphasizes pathotype, ABR, BFA, and related gene carrying in E. coli isolated from intestinal content samples of rats. The study presents a low prevalence of aEPEC; nevertheless, a high rate of MDR isolated from urban rats was high. Moreover, most isolates present a high rate of azithromycin resistance, which is found as a carrier of the macrolide resistance gene. A large proportion of isolates present moderate-to-strong BFA, however no correlation between BFA and ABR was found. The authors suggested that a transmission study of ABR-E. coli from rat reservoirs to humans is needed in Thailand, and rats as bioindicators for novel ABR surveillance should be established. The transmission of aEPEC and MDR-E. coli among humans, animals, and environmental reservoirs requires comprehensive surveillance systems, particularly, molecular surveillance, which might improve detecting emerging clones and spreading tracks. The intervention helps to understand the whole transmission chain of aEPEC and MDR-E. coli from rats to other hosts.
Footnotes
Acknowledgments
The authors express their gratitude to the staff at the Akkhraratchakumari Veterinary College, Walailak University, Nakhon Si Thammarat, Thailand, as well as the Research Institute for Health Sciences, Walailak University, Nakhon Si Thammarat, Thailand for providing the laboratory and collecting the samples.
Authors’ Contributions
W.M.: Methodology, investigation, conceptualization, writing—original draft. Ph.S.: Methodology, formal analysis, investigation, data curation and resources, writing—original draft, conceptualization and supervision, project administration, and funding acquisition. S.T.: Methodology, investigation, writing—original draft. N.T.: Methodology, investigation. Pa.S.: Investigation. K.C.: Methodology, investigation. S.B.: Writing—review and editing. Y.M.: Writing—review and editing. All authors read and approved the final article.
Accession Numbers
This article contains DNA sequence information. The nucleotide sequence data of the eae (intimin) gene from two aEPEC isolates were submitted to GenBank (NCBI) and assigned the following accession numbers: PP856229-PP856230.
Disclosure Statement
No competing financial interests exist.
Funding Information
The financial aspect of this study was partially supported by the Walailak University’s Individual Research Grant, Walailak University (Grant numbers: WU66271).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.