
Abstract
Growing evidence supports the efficacy of antimicrobial peptides against foodborne pathogens, though their antimicrobial spectrum and mechanism can vary depending on their origin. We investigated the antimicrobial spectrum of antimicrobial peptides derived from Lactobacillus paracasei A1, their effects on the survival rate and bactericidal mechanisms against Vibrio parahaemolyticus, and identified the functional short peptides within them. The crude extracts of antimicrobial peptides exhibited antibacterial properties against 13 pathogenic bacteria, showing strong inhibition of V. parahaemolyticus by disrupting the structural integrity of cell membranes. At minimum inhibitory concentrations, these peptides significantly disrupted the initial adhesion, membrane formation, and existing biofilms of V. parahaemolyticus, effectively inhibiting pathogen spread, enhancing the efficacy of antimicrobial agents, and reducing food safety risks. LC–MS/MS identification revealed four effective short peptides, all demonstrating potent bacteriostatic effects against V. parahaemolyticus. Our findings indicate that antimicrobial peptides can effectively destroy bacterial structures as well as the stability and regeneration of biofilms, making them promising candidates for use as food additives to control foodborne pathogens.
Keywords
Introduction
Vibrio parahaemolyticus is a common pathogen in the seafood industry, causing acute gastroenteritis and hemorrhage in food poisoning cases. The detection rate of V. parahaemolyticus in the seafood consumption markets of Europe and China is around 20%, highlighting the need to mitigate foodborne illness risks in the seafood industry (Koutsoumanis et al., 2024; Huang et al., 2024). V. parahaemolyticus thrives in elevated temperatures, appropriate salt concentrations, and neutral pH environments, all of which facilitate the formation and development of biofilms (Brauge et al., 2024; Gautam et al., 2024; Han et al., 2016). These biofilms promote colonization, host infection, and cross-contamination between processing equipment and food (Su et al., 2025).
Antibiotics are extensively used in aquaculture to control pathogens, but this has led to the emergence of drug-resistant strains (Hu et al., 2020; Su and Chen, 2020). Hence, there is a growing interest in natural antibacterial substances to reduce drug resistance. Antimicrobial peptides from lactic acid bacteria (LAB) are promising candidates due to their strong bactericidal effects, safety, minimal side effects, and prolonged activity (Guryanova, 2023). Studies have shown that antimicrobial peptides (Qiao et al., 2021; Shen et al., 2023; Tan et al., 2021) exhibit broad-spectrum antibacterial activity. However, there is limited research on the antimicrobial mechanisms of Lactobacillus paracasei peptides against V. parahaemolyticus.
In our previous study, L. paracasei A1 from LAB showed strong antibacterial activity against various pathogens. This study aimed to purify the antibacterial peptides from L. paracasei A1 and to investigate their mechanisms by analyzing changes in V. parahaemolyticus cell, laying the groundwork for promoting these peptides as natural preservatives in the food industry.
Materials and Methods
Bacterial strains
Fourteen types of microorganisms, comprising 13 pathogen strains and L. paracasei A1, were listed in Supplementary Table S1. The experiments conducted adhered to the Regulations on Biosafety Management of Pathogenic Microorganism Laboratories of China and received approval from Zhejiang Wanli University.
Crude extraction of antimicrobial peptides
L. paracasei A1 was harvested in the mid-logarithmic phase. Following the extraction method (Wu, 2020), the cell-free supernatant was obtained by centrifugation and mixed with ammonium sulfate kept at 4°C overnight. After centrifugation, the precipitates were washed, dissolved, and centrifuged. The supernatant was collected to obtain the crude extracts of antimicrobial peptides, and the pH was subsequently adjusted to 7.0 for further research.
Antibacterial spectrum of crude antimicrobial peptide extracts
Thirteen types of pathogenic microorganisms were activated, cultured to the logarithmic phase, and adjusted to 1 × 107 colony-forming unit (CFU)/mL. The antibacterial spectrum was assessed using the inhibition zone test for antimicrobial peptides (Chen et al., 2016). Each pathogenic microbial solution was mixed with nutrient agar. In total, 40 μL of crude antimicrobial peptides extracts were added into the Oxford cup, and an equal volume of 0.9% NaCl was used as a control. The plates were incubated at 28°C or 37°C for 24–48 h.
Purification of antimicrobial peptides
The purification process of antimicrobial peptides followed a reported method with slight modifications (Zhao et al., 2016). The crude extracts were mixed with n-butanol, stirred magnetically, and subjected to rotary evaporation. The residue was redissolved in sterile water to obtain the purified antimicrobial peptide, designated as LP-A1S.
Antibacterial assay of LP-A1S
The twofold microdilution method was modified to determine the minimal inhibitory concentration (MIC) of various agents (Chen et al., 2016). Eight serial twofold dilutions of LP-A1S were prepared, ranging from 6.00 to 0.047 mg/mL. An 80-μL aliquot of each dilution was added to the wells of a 96-well plate. The mid-logarithmic phase of V. parahaemolyticus was adjusted to 105 CFU/mL. A 20-μL aliquot of this bacterial suspension was added to each well and incubated at 28°C for 24 h. The MIC was determined by measuring the absorbance of bacterial lipids at 600 nm.
Cell viability test
The cell viability of V. parahaemolyticus was determined using trypan blue dye. Cells were harvested in the mid-logarithmic phase and adjusted to 1 × 107 CFU/mL. LP-A1S was added to the suspension at final concentrations of 0× MIC, 1× MIC, 2× MIC, and 10× MIC and incubated. The cell suspension was mixed with trypan blue dye, and viable and dead cells were counted within 3 min. The cell viability was calculated using the equation (1):
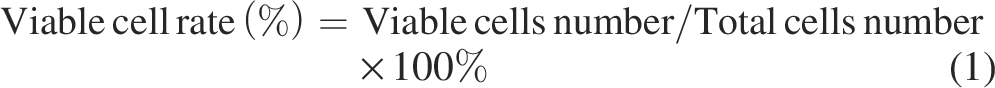
Scanning electron microscope
LP-A1S was mixed with V. parahaemolyticus to achieve 0× MIC, 1× MIC, 2× MIC, and 10× MIC. The cultures were incubated, washed, and fixed in glutaraldehyde. The cells were dehydrated using a gradient ethanol series. The samples were dried and coated with gold powder. Finally, the samples were imaged using a scanning electron microscope (SEM).
Transmission electron microscope
V. parahaemolyticus was mixed with LP-A1S at final concentrations of 0× MIC, 1× MIC, 2× MIC, and 10× MIC. The bacteria were then treated with glutaraldehyde and rinsed with PBS. Next, the samples were fixed with osmic acid and rinsed with PBS. The samples underwent dehydration with a gradient ethanol series, followed by absolute ethyl alcohol and acetone. The samples were then mixed with acetone and embedding agent. The final embedding was performed using a pure embedding agent. The samples were ultrathin sectioned and stained with uranyl acetate and lead citrate solution. Images were then observed and collected using a transmission electron microscope (TEM).
Cell permeability changes
Following the method reported (Cao et al., 2019), V. parahaemolyticus was adjusted to 1 × 107 CFU/mL. LP-A1S was added at final concentrations of 0× MIC, 1× MIC, 2× MIC, and 10× MIC, and the mixture was incubated. Subsequently, 5 μmol/L Fluo-4/AM solution was added to the mixture in the dark. The relative fluorescence intensity was measured and analyzed at an excitation wavelength of 488 nm, with a scanning wavelength range of 460–510 nm, D-Hanks solution as the blank control. Relative fluorescence intensity (%) was calculated using the following equation (2):
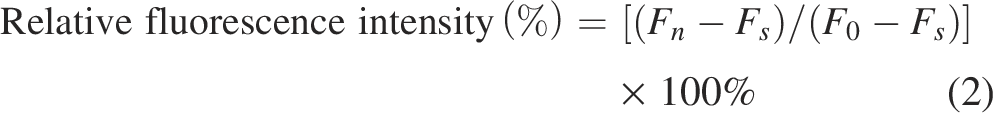
Initial adhesion of cells
Following the modified method (Shen et al., 2023), 20 μL LP-A1S, ranging from 2 to 0.125 MIC final concentration, was added to 180 μL of V. parahaemolyticus adjusted to 1 × 108 CFU/mL in a 96-well plate. The mixture was removed, and each well was washed with sterile PBS and fixed with anhydrous methanol. After discarding the methanol, the cells were air-dried, stained with ammonium oxalate crystal violet, and then dissolved in glacial acetic acid. The absorbance was measured at 600 nm. The effect of LP-A1S on the initial adhesion of bacteria was calculated using the following equation (3):

Formation of new biofilms
Following a modified method (Shen et al., 2023), the concentration of V. parahaemolyticus was adjusted to 1 × 105 CFU/mL. A 180-μL bacterial solution or culture medium was combined with 20 μL of twofold dilutions of LP-A1S (from 2 to 0.125 MIC final concentration) in each well of a 96-well plate, and the same amount of PBS was used as a control instead of LP-A1S. The wells were stained with ammonium oxalate crystal violet, and the stained biofilms were dissolved in glacial acetic acid. The absorbance was measured at 600 nm. The inhibitory effect of LP-A1S on the formation of new biofilms was calculated using the following equation (3).
Destruction of existing biofilms
Following a modified method (Shen et al., 2023), the concentration of V. parahaemolyticus was adjusted to 1 × 105 CFU/mL. A 200-μL bacterial solution or culture medium was added to each well of a 96-well plate. The liquid portion was removed, and the wells were rinsed with sterile PBS. In total, 180 μL of 10% medium and 20 μL of twofold dilutions of LP-A1S (from 2 to 0.125 MIC final concentration) or PBS as controls were added to the wells and incubated. The wells were then stained with ammonium oxalate crystal violet. The stained biofilms were dissolved in glacial acetic acid, and the absorbance was measured at 600 nm. The inhibitory effect of LP-A1S on the existing biofilms was calculated using the following equation (3).
Antimicrobial peptide sequence identification
Following the method (Jiang et al., 2022), the amino acid sequences of purified LP-A1S were identified using LC–MS/MS at Anhui Shenjian Detection Technology Co., Ltd. The steps were as follows: the purified LP-A1S was desalted and concentrated using a ZipTip C18 column with dissolution solution (0.1% formic acid and 5% acetonitrile). After centrifugation, 8 μL of the supernatant was analyzed by MS using the following conditions: mobile phase, 0.1% (v/v) formic acid in water (A) and a 0.1% (v/v) formic acid in acetonitrile (B) at 500 nL/min. A linear gradient was used as follows: 0–3.0 min, 3% B; 3.0–31.0 min, from 3% to 99% B; 31.0–35.0 min, 99% B; 35.0–36.0 min, from 99% to 3% B; and 36.0–40.0 min, 3% B. UV absorbance of the signal was monitored at 225 nm. Primary mass spectra were obtained in the range of 350–1550 m/z through a single full scan, and secondary mass spectra data were acquired using stepped normalized collision energy. The resulting sequences were identified in the PEAKS software de novo sequencing/database.
Using the published methods (Duan et al., 2021; Yu et al., 2018), the hydrophilicity >0.2 or <−0.2, aliphatic >25.00, and stability >4.00 indexes of the identified peptide sequences were calculated using the ExPASy ProtParam server. Hydrophobicity, electrostatic charge, and pI of the peptides were calculated using PepDraw. The BIOPEP database was used to predict the potential biological activities of the identified peptide sequences.
Antibacterial properties of synthetic peptides
Utilizing the characteristics of peptide sequences, antibacterial peptides were synthesized and their antibacterial properties were verified using the microbroth method. Through gradient dilution, the MIC of the synthetic peptides against V. parahaemolyticus was determined.
Statistical analysis
Each experiment’s data were obtained from three repeated experiments, and each repetition included three biological replicates. All data presented in the figures and tables are expressed as mean ± standard error of the mean. Statistical analyses were conducted using one-way analysis of variance followed by Tukey’s post hoc test, with SPSS version 13.0. A p value of <0.05 was considered statistically significant.
Results
Antibacterial activity of antimicrobial peptide
The crude extracts of antimicrobial peptides from L. paracasei A1 exhibited broad-spectrum antibacterial activity against 13 pathogen strains (Supplementary Table S1). Notably, it demonstrated strong antibacterial activity against nine pathogenic microorganisms, including V. alginolyticus, V. harveyi, V. parahaemolyticus, and so on. The antimicrobial peptide LP-A1S showed a MIC value of 0.75 mg/mL against V. parahaemolyticus.
Cell viability of V. parahaemolyticus with LP-A1S
As the concentration of LP-A1S increased, the survival V. parahaemolyticus decreased significantly (Fig. 1A). Specifically, the cell survival rates were 99.47%, 33.80%, 4.37%, and 0.56% after treatment with LP-A1S 0× MIC, 1× MIC, 2× MIC, and 10× MIC, respectively (Fig. 1B).
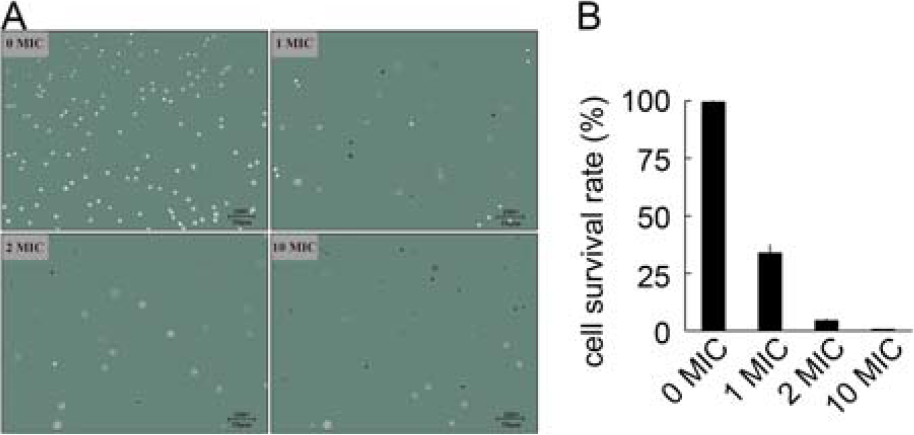
Cell viability of Vibrio parahaemolyticus with LP-A1S. The MIC used was 0.75 mg/mL. 0 MIC is control, whereas 1 MIC, 2 MIC, and 10 MIC are treatment groups, respectively. MIC, minimal inhibitory concentration.
LP-A1S acts on cell membrane of V. parahaemolyticus
As shown in Figure 2A, the fluorescence intensity of V. parahaemolyticus cells stained with Fluo-4/AM peaked at a wavelength of 488.2 nm. Compared with 0× MIC LP-A1S, the intracellular fluorescence intensity of bacteria treated with 1× MIC, 2× MIC, and 10× MIC LP-A1S decreased to 58.84%, 43.98%, and 28.18% of 0× MIC LP-A1S, respectively (Fig. 2B). This indicates that intracellular fluorescence intensity diminishes as the concentration of LP-A1S increases.

LP-A1S affects fluorescence intensity of Vibrio parahaemolyticus. The MIC used was 0.75 mg/mL. 0 MIC is control, whereas 1 MIC, 2 MIC, and 10 MIC are treatment groups, respectively. *p < 0.05, to 0 MIC as the control. MIC, minimal inhibitory concentration.
SEM images revealed that V. parahaemolyticus treated with 0× MIC LP-A1S retained a full shape, clear outline, complete structure, smooth surface, and no content leakage, appearing as short rods with blunt ends (Fig. 3A). In contrast, cells treated with increasing concentrations of LP-A1S displayed significant deformations, with the cell membrane showing depressions and shrinkage (Fig. 3B–D). Higher LP-A1S concentrations caused more ruffles on the bacterial surface, resulting in mostly spherical forms and substantial content leakage.
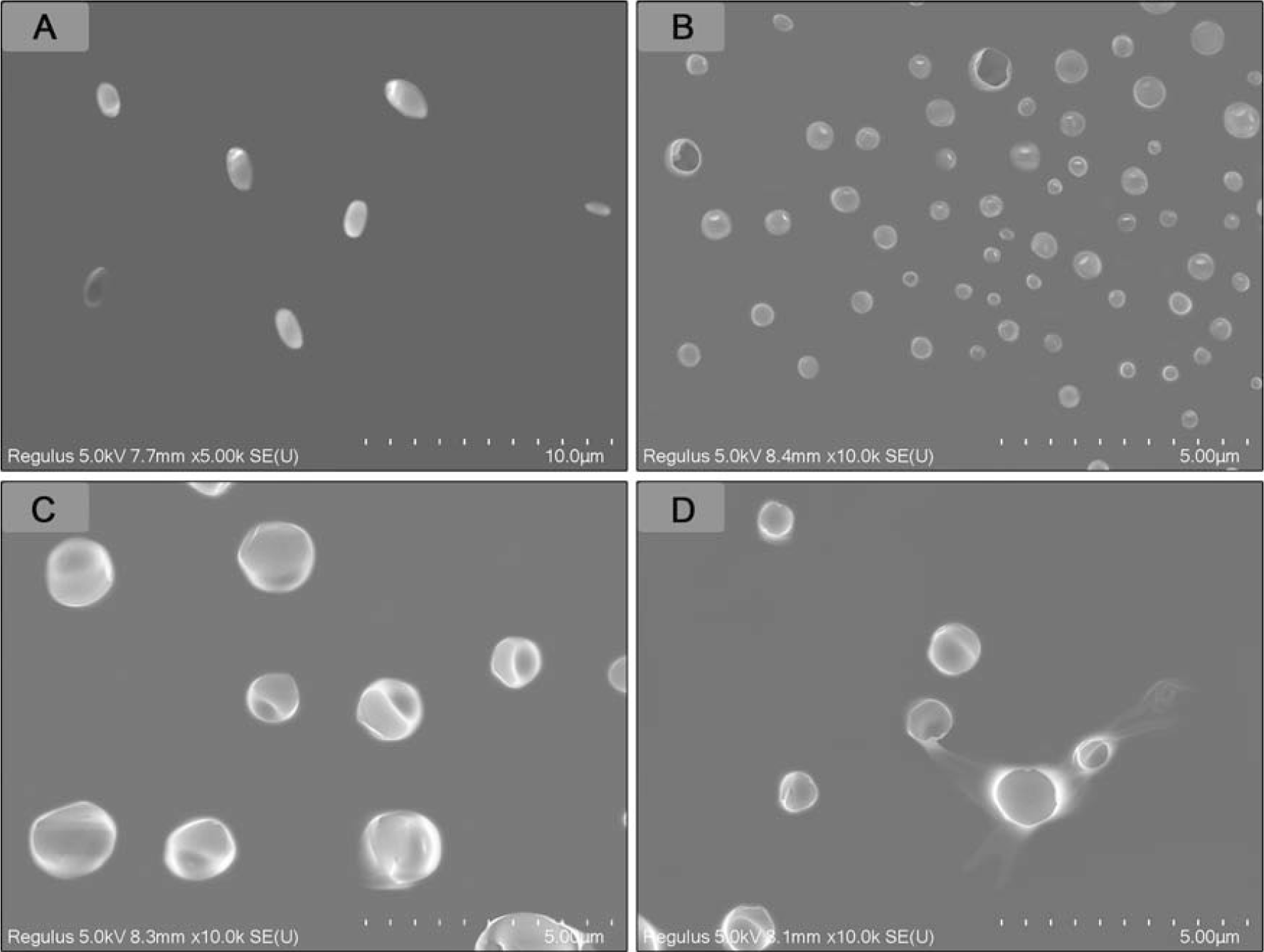
LP-A1S affects Vibrio parahaemolyticus by SEM. V. parahaemolyticus were treated with LP-A1S at the final concentration of 0 MIC
TEM ultrathin section analysis supported these findings. In the group treated with 0× MIC LP-A1S, V. parahaemolyticus cells had smooth, neat surfaces, tightly bound cell walls and membranes, and uniformly abundant cell contents (Fig. 4A). However, after treatment with 1× MIC LP-A1S, bacterial edges became blurred and shriveled, with cavities forming inside, complicating the maintenance of their rod-like shape, and partially detaching the cell wall from the membrane (Fig. 4B). At 2× MIC LP-A1S, the bacteria exhibited irregular shapes, ruptured cell membranes, noticeable perforations, cavities, and content leakage (Fig. 4C). At 10× MIC LP-A1S, the bacteria were completely disintegrated (Fig. 4D).
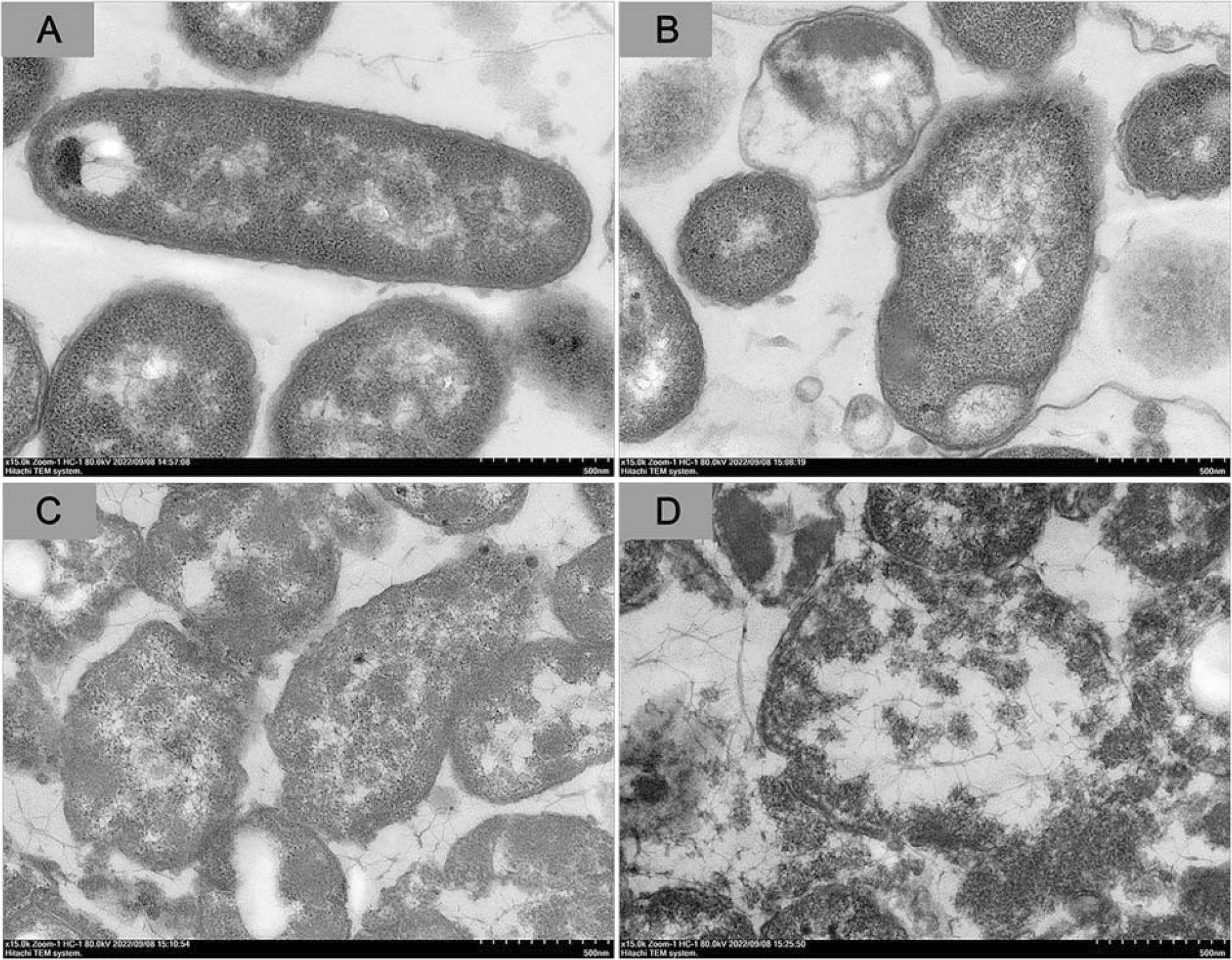
LP-A1S affects Vibrio parahaemolyticus by TEM. V. parahaemolyticus were treated with LP-A1S at the final concentration of 0 MIC
Biofilm analysis
After treatment with 0.125 to 2× MIC LP-A1S, the initial adhesion rates of V. parahaemolyticus were 26.60%, 24.30%, 21.77%, 19.27%, and 17.99% of the group treated with 0× MIC, respectively (Fig. 5A). Similarly, the new biofilm formation rates under these concentrations were 37.80%, 28.75%, 24.99%, 22.35%, and 20.15% of the group treated with 0× MIC, respectively (Fig. 5B). Furthermore, the destruction rates of old biofilms at the same LP-A1S concentrations were 58.14%, 48.31%, 41.18%, 13.22%, and 8.03% compared with the group with 0× MIC, respectively (Fig. 5C).

Effect of LP-A1S treatment on biofilm of Vibrio parahaemolyticus. The MIC used was 0.75 mg /mL. 0 MIC is control, whereas 0.125 MIC, 0.25 MIC, 0.5 MIC, 1 MIC, and 2 MIC are treatment groups, respectively. a, b, c, d, e, and f; p < 0.05. MIC, minimal inhibitory concentration.
Antimicrobial peptide sequence identification
The peptides in LP-A1S appeared in the latter part of the mass spectrum (Fig. 6). Secondary mass spectrometry identified 10 short peptides, labeled P1 to P10 (Supplementary Fig. S1). Comparing these sequences with the PEAKS and BIOPEP databases revealed no matches, indicating that these peptides are novel. They consist of 6–9 amino acids, with molecular weights between 723.43 and 980.43 Da, and mass-to-charge ratios ranging from 376.22 to 491.22 m/z. Each peptide has a unique amino acid arrangement (Supplementary Table S2).
Total particle flow diagram of LP-A1S. Chromatographic conditions: column: POROS R2/10 (50 × 4.6 mm); mobile phases, 0.1% (v/v) formic acid in water
Identification of properties of peptides
The properties of the 10 identified peptides were predicted. Hydrophobicity ranged from +2.59 to +16.83 Kcal/mol, hydrophilicity from −1.65 to 1.95 Kcal/mol, aliphatic index from 25.00 to 182.50, pI from 2.93 to 10.57, and net charge from −1 to +2. Stability scores ranged from 4.10 to 104.63. These findings indicate that all 10 peptides possess good thermal stability, solubility, and in vitro stability (Supplementary Table S3).
In terms of antibacterial potential, peptides P1, P3, P7, P8, and P9 demonstrated antibacterial activity, while P2, P4, P5, P6, and P10 exhibited antibacterial aptamer activity (Supplementary Table S4). Additionally, database analysis revealed potential anti-oxidative properties and the ability to inhibit angiotensin-converting enzyme and dipeptidyl peptidase (Supplementary Table S4).
MICs of synthetic peptides based on these sequences against V. parahaemolyticus were measured. Peptides P2, P3, P4, and P5 showed antibacterial activity with MICs of 1.5, 3, 1.5, and 3 mg/mL, respectively (Supplementary Table S5).
Discussion
In 2021, China’s Ministry of Agriculture and Rural Affairs highlighted the rapid growth of aquaculture in China. Despite this growth, the industry faces significant challenges due to disease outbreaks, resulting in substantial economic losses exceeding 2.31 billion yuan annually by the end of 2019, accounting for a significant portion of China’s fishery disaster losses (Yu et al., 2020). Efficient and broad-spectrum antimicrobial peptides are widely recognized for their potential to serve as a green, economical, and safe solution in the prevention and control of aquaculture diseases (Wang and Cheng, 2024). This study discovered that purified LP-A1S exhibits potent bactericidal effects against various microorganisms. The efficacy against V. parahaemolyticus increased with higher concentrations of LP-A1S, causing significant cell death. Ultramicroscopic analysis revealed a gradient damage pattern on the cell membrane of V. parahaemolyticus by LP-A1S, with high concentrations leading to disruption of bacterial nuclear structures and cell lysis. Recent studies have highlighted similar broad-spectrum antibacterial activities of peptides derived from animals, which have also been shown to exhibit significant antibacterial effects against V. parahaemolyticus (Liu et al., 2023; Zhao et al., 2024). These peptides, like LP-A1S, disrupt microbial cell structures and enhance cell membrane permeability to exert their antibacterial effects. Comparable findings in other literature (Cao et al., 2019; Gupta et al., 2016; Zhang et al., 2022) underscore the potential of LAB-derived peptides in combating pathogenic bacteria, indicating promising applications in various fields.
Biofilms of pathogenic bacteria, such as V. parahaemolyticus, pose significant public health risks due to their ability to adhere to surfaces, resist removal, develop antibiotic resistance, and interfere with immune responses. In our study, we investigated that LP-A1S not only inhibited initial biofilm adhesion but also prevented new biofilm formation and disrupted existing biofilms of V. parahaemolyticus. This finding holds crucial implications for food safety. Research has shown that various antimicrobial peptides, like LP-A1S, effectively inhibit pathogenic biofilms by mechanisms possibly involving extracellular matrix degradation (Krishnamoorthi et al., 2022; Roy et al., 2018; Wang et al., 2023). Antimicrobial peptide A20L (Zhou et al., 2024) is capable of both preventing biofilm formation and disrupting the stability of existing biofilms. Additionally, ferredoxin (Choi et al., 2024) has been shown to inhibit the formation of bacterial biofilms. These findings further demonstrate that LP-A1S is a natural antimicrobial peptide derived from the microorganism, possessing multiple antibiofilm activities.
Analysis of LP-A1S components revealed 10 short peptides with favorable properties such as thermostability, solubility, and antimicrobial potential. These results suggest that achieving effective antimicrobial activity involves balancing the distribution of hydrophobicity and charge, as well as ensuring the structural adaptability of the peptides (Pirtskhalava et al., 2021). The development of antimicrobial peptides (Chen et al., 2016) with biofilm inhibition capabilities represents a promising avenue for mitigating bacterial infections and addressing antibiotic resistance concerns in medical and food safety applications.
Authors’ Contributions
F.G. and Y.Y. conceived and designed the research. F.G., C.Y., L.Y., and Y.Y. conducted the experiments and analyzed the data. F.G. and C.Y. wrote the article. F.G. and Y.Y. critically reviewed the article. F.G. and Y.Y. supervised and revised the article. All authors read and approved the final article.
Footnotes
Author Disclosure Statement
The authors report no conflicts of interest.
Funding Information
This study received support from the Zhejiang Science and Technology Department Project (grant number LGN22D060003); Natural Science Foundation of Zhejiang Province (grant number LY22C190006); and Zhejiang Provincial Top Discipline of Biological Engineering (Level A) (grant number KF2023010).
Disclaimer
The authors alone are responsible for the content and writing of the article.