
Abstract
Trichinellosis, a zoonotic disease transmitted through food and caused by Trichinella spiralis, is a significant health concern worldwide. Therefore, developing a safe and effective vaccine to combat T. spiralis infection is essential. In this study, a nonantibiotic Lactobacillus plantarum strain lacking the alr gene served as a live bacterial vector to deliver antigens to the host, creating a novel oral vaccine. A nonantibiotic vaccine was constructed to coexpress T. spiralis Nudix hydrolase (TsNd protein) and the gp43 protein alongside a food-grade murine IL-33 adjuvant. The objective of this study was to evaluate the efficacy of this novel strategy in preventing trichinellosis and mitigating intestinal damage. The integrated vaccination strategy was found to increase specific serum immunoglobulin G and mucosal secretory IgA (sIgA) concentration in BALB/c mice. Additionally, this approach significantly increased the release of cytokines such as interleukin-4 (IL-4) from lymphocytes, along with increased CD4+ expression on lymph cells from both splenic and mesenteric sources. Post-larval exposure, significant reductions in the populations of adult worms by 59.80% at 7 d postinfection (dpi) and in the larval load by 51.29% at 28 dpi were documented. Furthermore, oral vaccination reduced the number of encysted larvae present in the tongue and masseter muscles after challenge with T. spiralis. Overall, the results demonstrated that combined vaccination with the IL-33 adjuvant effectively prevented trichinellosis and reduced intestinal damage, highlighting the potential of food-grade Lactobacillus vaccines in preventing intestinal parasitic infections, with IL-33 serving as an effective adjuvant.
Keywords
Introduction
Trichinellosis caused by Trichinella spiralis is a worldwide food-borne parasitic zoonosis (Iacob et al., 2022). Mebendazole and albendazole, two anthelmintic medications for treating T. spiralis in humans, may cause teratogenic effects in pregnant women (Shimoni and Froom, 2015). Therefore, developing a vaccine against foodborne infections is the optimal approach for managing parasite infestations and enhancing public health (Hewitson and Maizels, 2014). The three potential types of vaccines against T. spiralis are DNA, recombinant antigens, and purified native antigens. Considering the intricate life cycle of T. spiralis and its diverse array of antigenic components expressed across different infection stages, relying solely on a vaccine targeting a single antigen may be insufficient for combating T. spiralis infection. Therefore, vaccines designed to target T. spiralis infection typically require the incorporation of multiple antigenic components to ensure comprehensive immune protection across various stages of infection.
The excretory–secretory products found in muscle larvae (MLs) predominantly comprise glycoproteins derived from stichosomes. These glycoproteins function as target antigens that are directly presented to the host immune system during infection with T. spiralis. Research indicates that attenuated Salmonella expressing a 30-mer peptide from the Trichinella glycoprotein 43 (gp43) antigen can significantly increase immunoglobulin G (IgG) titers in mice (Castillo Alvarez et al., 2013). Nudix hydrolase (ND), which is present in diverse organisms, degrades harmful intracellular metabolites and mitigates oxidative stress. In T. spiralis, TsND binds to intestinal epithelial cells (IECs) and is upregulated during intestinal infective larval infection (Ren et al., 2013a, 2013b). Studies have shown that immunizing mice with TsND peptides that are displayed by the T7 phage results in protective immunity (Cui et al., 2013).
The intestinal stage is the first step of the host–parasite interaction occurring after oral T. spiralis infection. Recombinant lactic acid bacteria (LAB), which express foreign antigens, have the capacity to function as conduits for oral vaccine delivery while also acting as probiotics that increase the functional capacity of the immune system (Koko et al., 2019; Yu et al., 2017). Lactobacillus plantarum NC8 is widely utilized as a bacterial host for the expression of exogenous proteins and has demonstrated advantageous effects in combating infections from T. spiralis and coccidiosis (Liu et al., 2020b; Wang et al., 2020). Erythromycin, ampicillin, or kanamycin—markers of antibiotic resistance—remain components of conventional recombinant L. plantarum vaccinations. However, an increasing number of studies have indicated that antibiotic resistance genes have the potential to impact the environment; therefore, they may not be suitable for use in food or oral vaccine applications. Many nonantibiotic selection markers, including thr (Glenting et al., 2002) and alr (Lu et al., 2016), have been developed to address these drawbacks.
During helminth infections, IECs secrete interleukin-33 (IL-33), one of three key innate cytokines—along with IL-25 and thymic stromal lymphopoietin—which collectively trigger Th2 immune responses. IL-33 modulates immune cell activity by binding to its receptor IL-33R, which regulates the Th2 CD4+ T-cell response, thereby enhancing immune protection against helminth infections (Humphreys et al., 2008; Schmitz et al., 2005). Furthermore, IL-33 can stimulate the expression of Th2 cytokines, such as IL-4, IL-9, and IL-13, which are crucial for the clearance of parasitic worms (Cortés et al., 2017; Humphreys et al., 2008). This is the first study to show that IL-33 isoforms are effective adjuvants that assist in the creation of vaccines by boosting antigen-specific effector and memory T-cell responses in vivo and encouraging therapeutic effects in Human papillomavirus (HPV) related cancer immunotherapy (Villarreal et al., 2014). Nevertheless, to date, no comprehensive studies have explored the use of IL-33 as an adjuvant.
In this study, we utilized the L. plantarum NC8 strain, in which antibiotic resistance genes were knocked out as a food-grade expression system. This system was used to coexpress the T. spiralis TsNd and gp43 proteins combined with a murine-derived IL-33 adjuvant to enhance mouse immune function and resistance to T. spiralis infection. Our findings present a novel strategy for the development of vaccines and adjuvants against T. spiralis, highlighting the potential of food-grade Lactobacillus vaccines for preventing intestinal parasitic infections, with IL-33 as an effective adjuvant.
Materials and Methods
Plasmids, bacterial strains, and growth conditions
The plasmids used in the study are listed in Table 1, and the bacterial strains used are listed in Table 2. The pLp-TsNd-S-pgsA′-gp43 plasmid, a coexpression vector for the T. spiralis proteins TsNd and gp43, was constructed by Wang et al. (2021). The plasmid IL-33 (GenBank: AY905582.1), containing murine IL-33 and carrying XbaI and HindIII with flag restriction sites, was constructed by our laboratory. The plasmid pSIP409-pgsA′ carrying XbaI and HindIII restriction sites was constructed by Cai et al. (2016). The plasmid pLp-S carrying SalI and XhoI and the plasmid pLP-1261 Inv, a derivative of pLp_2588sAmyA with the erm gene replaced by asd-alr, were provided by Liu et al. (2020a). All the plasmids were stored at −80°C in our laboratory. Escherichia coli and L. plantarum NC8 were cultured in lysogeny broth and De Man, Rogosa, and Sharpe (MRS) media, respectively, each maintained at 30°C without stirring. Competent L. plantarum NC8/Δalr and E. coli χ6212 strains were prepared and cultured according to methods described previously (Galán et al., 1990; Josson et al., 1989). Unless otherwise specified, erythromycin was used at 10 μg/mL for L. plantarum NC8 and at 200 μg/mL for the other strains. After overnight incubation, the liquid cultures were diluted in fresh MRS medium until an optical density (OD600) of approximately 0.3 was reached, resulting in a concentration of 1 × 109 colony-forming units (CFUs) per 200 µL. In this study, the T. spiralis isolate (ISS534) utilized was derived from domestic pigs in Henan Province, China, and has been propagated consistently in Kunming mice.
Strains Used in This Study

Plasmids Used in This Study

Development of a food-grade expression vector for the T. spiralis proteins TsNd and gp43
The asd-alr fragment was seamlessly integrated into the pLP-1261 Inv plasmid via seamless integration of the ATA-F/R primers. To construct the pLp-TsNd-S-pgsA′-gp43 plasmid without the erm gene, we used 409 (ata) F/R primers for seamless cloning. These fragments, along with the asd-alr fusion fragments, were then seamlessly assembled to create the pLp-TsNd-S-pgsA′-gp43(ata) plasmids. Using the asd gene as an auxotrophic marker, we initially amplified these expression vectors in E. coli χ6212-competent cells and subsequently introduced them into the L. plantarum NC8/Δalr strain via electroporation. This strain was constructed using a Cre-lox-based vector mutagenesis system (pNZ5319/pNZ5348) (Lambert et al., 2007), as described in detail by Nguyen et al. (2011).
Construction of a food-grade expression for Mus musculus IL-33
IL-33 fragments were amplified from the plasmid IL-33 via the IL-33F/R primers and subsequently cloned and inserted into pEasy-Blunt (Table 3). The single expression plasmid for Mus musculus IL-33 enables the expressed protein to be presented on the surface of the bacterial cell. The resulting vector pEasy-Blunt-IL-33 (DH5α) carried the common XbaI/HindII restriction sites with those of pSIP409-pgsA′. The plasmid underwent digestion and ligation, which led to the formation of pSIP409-pgsA′-IL-33 (DH5α), which was subsequently transformed into L. plantarum NC8 (Sørvig et al., 2005) via electroporation, producing NC8-pSIP409-pgsA′-IL-33. The process described in the “Development of a food-grade expression vector for the T. spiralis proteins TsNd and gp43” section was used to guarantee the effective surface display of IL-33 in the L. plantarum NC8/Δalr strain, yielding NC8/Δalr-pSIP409-pgsA′-IL-33(ata). We used the same cloning method for IL-33, applying the asd-alr segment as a marker and utilizing common restriction sites, and then followed the same transformation protocol for L. plantarum NC8/Δalr.
List of PCR Primers Sequence

PCR, polymerase chain reaction.
Western blot and indirect immunofluorescence assay for detection
The recombinant strains harboring the vectors NC8/Δalr-pLp-TsNd-S-pgsA′-gp43(ata) and NC8/Δalr-pSIP409-pgsA′-IL-33(ata) were aerobically incubated in MRS medium until they reached OD600 nm at 30°C with agitation. Subsequently, bacterial suspensions were used to induce protein expression, followed by ultrasonic disruption on ice to release the target proteins. A total of 2 mL of these bacterial cultures was collected and used to assess the coexpression of TsNd and gp43 via mouse anti-TsNd and mouse anti-gp43 polyclonal antibodies at a 1:5000 dilution. Additionally, the expression of IL-33 was analyzed utilizing a mouse anti-IL-33 polyclonal antibody, also at a dilution of 1:5000.
Indirect immunofluorescence assay (IFA) was used to detect surface-anchored proteins. Bacterial pellets were incubated with polyclonal antibodies at room temperature for 1 h, then with 200 μL fluorescein isothiocyanate (FITC)-conjugated goat antimouse IgG (1:2000) at 37°C in the dark for 1 h. Cells were placed on sterilized slides, fixed with 4% paraformaldehyde for 10 min, and observed under a fluorescence microscope. The same procedure was used to prepare the NC8/Δalr strain, which served as a control.
Oral immunization procedures in mice
All experimental animals were approved by the Animal Care and Ethics Committees of Jilin Agriculture University 200707001. Six-week-old wild-type female BALB/c mice (specific pathogen free, 19 ± 1 g) were obtained from Beijing Huafukang Biotechnology Co., Ltd., China. All mice were maintained on a 12-h light/12-h dark cycle and had ad libitum access to food and water throughout all tests. The mice were allowed to acclimate for 7 d prior to the administration of the vaccines to ensure their adaptation to the environment. Next, one-hundred and twenty 6-week-old female BALB/c mice were evenly divided into six groups, each containing 20 mice: the phosphate-buffered saline (PBS) group, control group (empty vector), IL-33 group (recombinant IL-33 group), TsNd-gp43 group (recombinant TsNd-gp43 group), TsND-gp43+IL-33 group (recombinant TsNd-gp43 + recombinant IL-33 (1:1) group), and T. spiralis group (MLs of 300T. spiralis). On day −35, a total of 10 mice from each group were euthanized for the purpose of conducting enzyme-linked immunosorbent assays (ELISAs). Serum and fecal samples were collected and stored at −80°C until use, and cells from the spleen (SPL), mesenteric lymph nodes (MLNs), and Peyer’s patches (PPs) were harvested for flow cytometry. Additionally, 15 mice were utilized to evaluate the burdens of intestinal adult worms (AWs) and MLs, except for those in the PBS control group. Five mice were euthanized at 7 d postvaccination (7 dpi) for AW counts and duodenum histopathology was performed via hematoxylin and eosin (HE) staining. Another 5 mice were euthanized at 28 dpi to measure the number of larvae per gram (LPG) and to prepare histopathology sections of the lingual and masseter muscles.
The vaccination protocols were implemented as outlined below: oral administration was performed three times at 14-d intervals prior to challenge with infectious T. spiralis. The mice in the control and recombinant LAB groups were orally administered 200 μL of live bacterial suspension at a cell density of 1 × 109 CFU. Similarly, the PBS group was orally administered 200 μL of sterile PBS. Prior to challenge with 300T. spiralis MLs, the group designated for this parasite was given an identical dose of PBS (Fig. 1).

The oral immunization was administered three times at 14-d intervals prior to the infection with 300T. spiralis muscle larvae on day 0. ELISA detection was conducted on day −35. Flow cytometry analyses were performed following the final immunization (on day −35) and again on 28 d postinfection (dpi). Additionally, the adult worms and muscle larvae were collected and quantified on 7 and 28 dpi to evaluate the immunoprotective efficacy of the recombinant vaccine and adjuvant against T. spiralis. ELISA, enzyme-linked immunosorbent assays.
Flow cytometry analysis of cytokines
The cells from the SPL, MLN, and PP of the mice were isolated as single-cell suspensions under sterile conditions and subjected to flow cytometry analysis as previously described (Wang et al., 2021). Briefly, 1 × 106 SPL and MLN cells were suspended in Roswell park memorial institute 1640 medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. The cells were stimulated with 50 ng/mL phorbol 12-myristate 13-acetate, 1 μg/mL ionomycin, and 10 μg/mL protein transport inhibitor for 5 h. Afterward, the cells were resuspended to perform fluorescence-activated cell sorting for extracellular marker detection via antimouse CD3+ and CD4+ antibodies. PP cells were stained with allophycocyanin-conjugated anti-mouse CD45R/B220, followed by washing and fixation. The levels of the intracellular markers IL-4, IL-17, and Interferon-gamma (INF-γ) were detected in SPL and MLN cells via appropriate antibodies. PP cells were stained with IgA antibodies, and samples were analyzed by flow cytometry. Additionally, SPL and MLN cells were stained with carboxyfluorescein succinimide ester, incubated with TsNd and gp43 proteins at 37°C for 72 h in vitro, and then stained with CyTM5.5 CD3ε and FITC CD4 to identify CD4+ T cells via flow cytometry. All these data were analyzed via FlowJo software (version 7.6.1; Tree Star Inc.).
Detection of systemic and mucosal antibody responses
ELISA plates were coated with purified mouse-derived TsNd and mouse-derived gp43 protein antigens (5 μg/mL) separately. Fecal and serum samples were used as the primary antibody source, followed by the addition of horseradish peroxidase-conjugated goat anti-mouse IgG/sIgA as the secondary antibody for detecting specific IgG and sIgA antibodies via ELISA as described previously (Wang et al., 2021).
Clinical evaluation of immunological efficacy
To assess the protective efficacy of the recombinant vaccine and adjuvant against T. spiralis infection, adult intestinal worms were collected and counted at 7 dpi, and duodenal samples were collected for histopathological analysis. At 28 dpi, MLs were collected, and histopathological sections of the lingual and masseter muscles were prepared for evaluation. The worm reduction rate was calculated by comparing the LPG content in muscle tissues between the immunized and control groups.
The samples were processed for paraffin section staining, fixed in 4% formalin for 48 h, embedded in paraffin wax, and prepared as 3-μm-thick muscle sections. After deparaffinization, the sections were stained with HE and observed under a microscope (Leica) at 200 × magnification.
Statistical analysis
Statistical analyses were conducted via GraphPad Prism 9.0 software, with the data presented as the means ± standard errors of the means. Group differences were assessed through one-way analysis of variance followed by post hoc tests. A significance threshold of p < 0.05 was applied to determine statistical significance.
Results
Construction and expression of target proteins in L. plantarum NC8/Δalr
The recombinant L. plantarum NC8/Δalr strain was identified via colony polymerase chain reaction. One strain expressing surface-anchored TsNd and gp43 was referred to the TsNd-gp43 vaccine (Fig. 2a–c), whereas the other strain expressing surface-anchored IL-33 was referred to the IL-33 adjuvant (Fig. 2b–c). Western blotting was used to detect the expression of target protein bands in the recombinant L. plantarum, anti-TsNd mouse polyclonal (62 kDa) and anti-gp43 mouse polyclonal (58 kDa) antibodies that recognized TsNd-gp43, indicating that they were colocalized (Supplementary Fig. S1a), and the anti-IL-33 mouse polyclonal (52 kDa) antibody recognized the IL-33 adjuvant (Supplementary Fig. S1b). IFA was applied to analyze recombinant protein expression in the strains. Fluorescence microscopy demonstrated an absence of green fluorescence on the surface of the control group, whereas the recombinant strains displayed pronounced fluorescence in short rod-shaped strains (Fig. 2d).
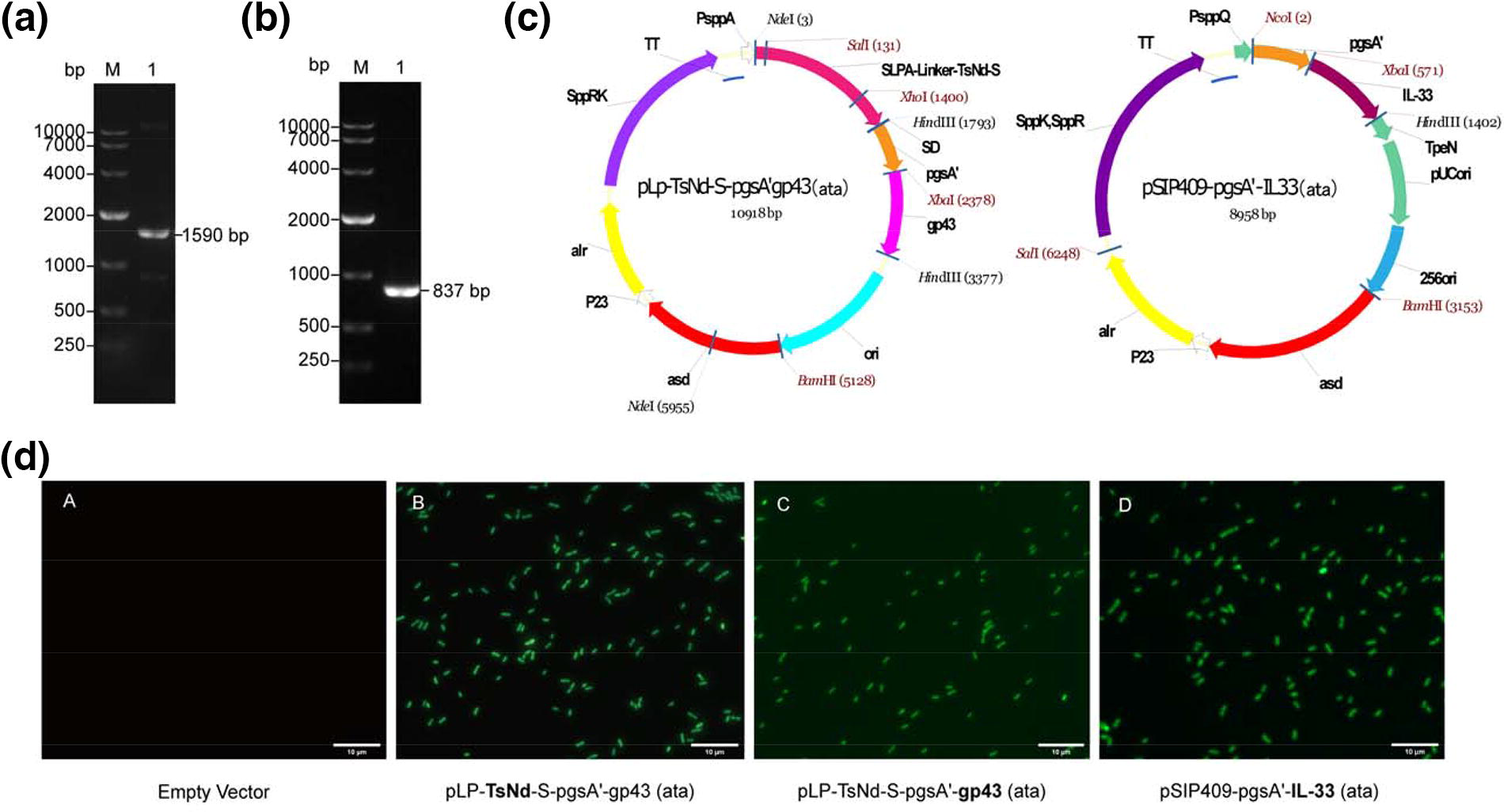
Construction and expression of target proteins in recombinant L. plantarum NC8/Δalr.
The TsNd-gp43 vaccine combined with the IL-33 adjuvant enhances systemic and mucosal antibody responses in mice
On day −35, both TsNd-specific and gp43-specific IgG levels in the serum of the mice in the TsND-gp43+IL-33 group and TsNd-gp43 group were significantly greater than those in the control group (p < 0.01). Similarly, the sIgA levels in the feces of the mice in the TsND-gp43+IL-33 group and TsNd-gp43 group were significantly greater than those in the control group (p < 0.01) (Fig. 3a). Compared with the control group, both the TsNd-gp43 (p < 0.05) and TsNd-gp43+IL-33 (p < 0.01) groups presented significantly greater numbers of IgA+ B cells (Fig. 3b). These results indicated that the T. spiralis vaccine combined with an adjuvant activated the humoral and mucosal immune response.

The TsNd-gp43 vaccine combined with IL-33 adjuvant enhanced specific IgG and sIgA levels.
The TsNd-gp43 vaccine combined with the IL-33 adjuvant enhances the cellular immune response in mice
When stimulated with the gp43 protein and TsNd protein in vitro, the number of CD4+ T cells in the TsNd-gp43 group was significantly greater than that in the control group, regardless of whether the T cells were in the SPL or MLN (p < 0.01). Compared with those in the control group, the number of CD4+ T cells in the SPL in the TsND-gp43+IL-33 group was significantly greater, and the MLN displayed a trend similar to that of the SPL results (p < 0.01) (Fig. 4a, c).
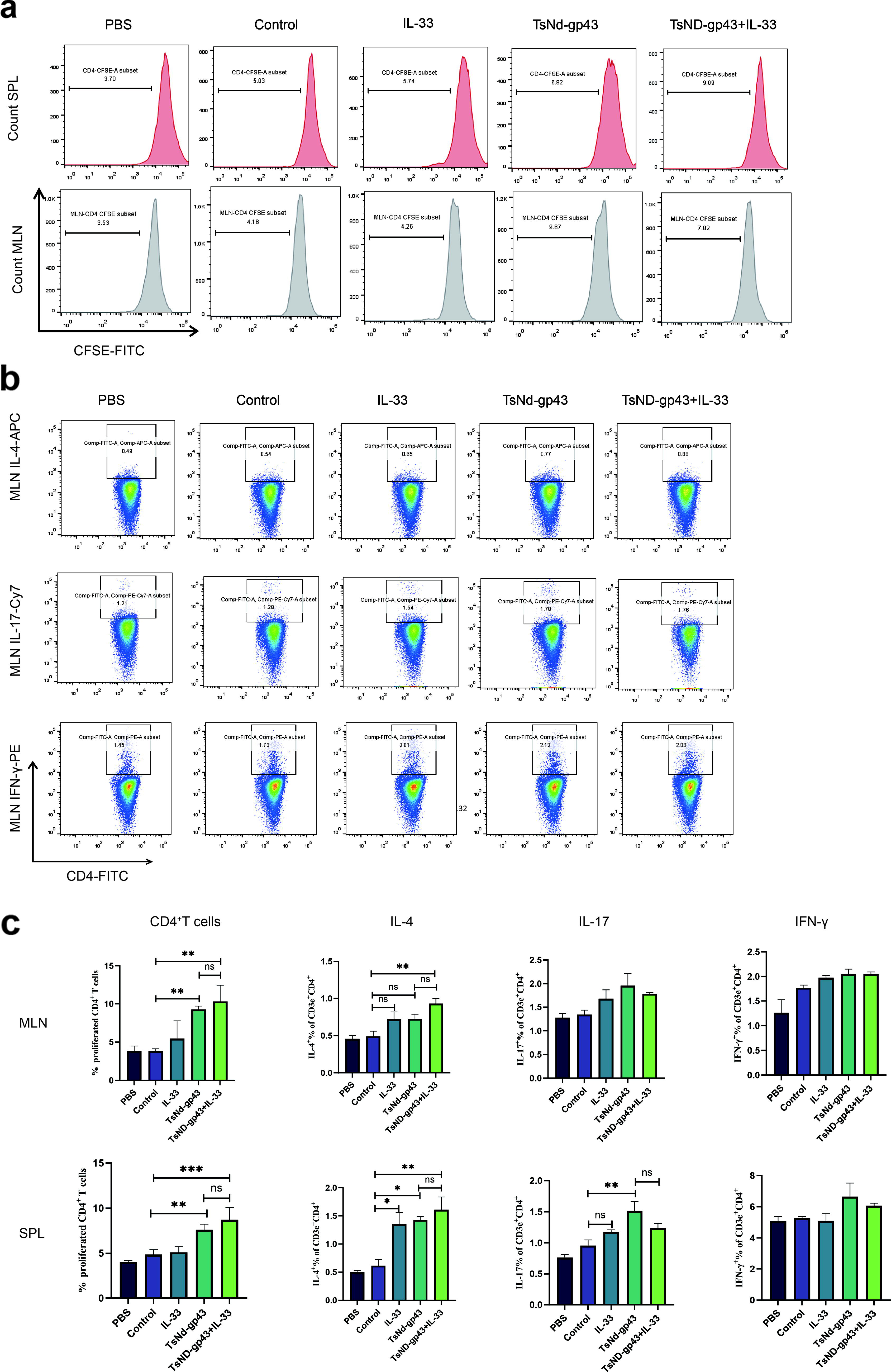
The TsNd-gp43 vaccine combined with the IL-33 adjuvant promotes immunological effectiveness and immune response.
Compared with the control group, the TsND-gp43+IL-33 group presented significantly greater IL-4 expression in both MLN and SPL cells (p < 0.01). In SPL cells, both the TsNd-gp43 group and IL-33 group exhibited a significant increase in IL-4 expression (p < 0.05), whereas no significant change was observed in MLN cells. Furthermore, only the SPL cells from the TsNd-gp43 group presented elevated levels of IL-17 expression (p < 0.01) (Fig. 4b–c). Notably, neither group showed any difference in the expression of the cytokine IFN-γ. These results indicated that the TsNd-gp43 vaccine could activate the Th17 immune response and that the addition of the IL-33 adjuvant further activated the Th2 immune response.
The TsNd–gp43 vaccine combined with the IL-33 adjuvant protected mice from infection by T. spiralis
Following infection with T. spiralis, the proportion of IL-4-producing SPL cells in the TsND-gp43+IL-33 group was significantly greater than that observed in both the control group (p < 0.01) and the T. spiralis group (p < 0.05), particularly the MLN cells (p < 0.01) (Fig. 5a–c). Furthermore, IL-4 expression in MLN cells was markedly elevated in the TsND-gp43 group compared with the T. spiralis group (p < 0.01), whereas no significant difference was detected in SPL cells. Additionally, there was no discernible difference in the expression of IL-17 and IFN-γ in either SPL or MLN cells between the TsNd-gp43 and TsND-gp43+IL-33 groups compared with the T. spiralis group (Fig. 5c). IgA-producing PP cells were significantly greater in the TsND-gp43+IL-33 group than in both the control group and the T. spiralis group, while there was no significant difference in the TsND-gp43 group (p < 0.01) (Fig. 5d). Following T. spiralis infection, TsNd-gp43 vaccination combined with the IL-33 adjuvant efficiently increased Th2 and intestinal mucosal immune responses.

The TsNd-gp43 vaccine combined with IL-33 adjuvant protected mice from 300T. spiralis muscle larvae infection.
Protective immunity against T. spiralis ML infection and histopathological analysis
The TsND-gp43+IL-33 group exhibited significant protection against T. spiralis infection (Table 4). However, there was no discernible difference between the TsND-gp43+IL-33 group and the TsND-gp43 group. The IL-33 group demonstrated partial protective effects against T. spiralis infection (Fig. 6a–b). Histopathological scoring of the duodenum revealed significantly lower scores in the TsND-gp43+IL-33 group than in the control and T. spiralis groups (p < 0.001). In the T. spiralis group, there was an increase in the number of inflammatory cells in the mucosal layer, epithelial cell edema, and significant villus shortening, atrophy, local defects, and vacuolar changes. The control group presented similar changes. In the IL-33 group, villus atrophy and defects were reduced, and the number of inflammatory cells was decreased. Both the TsNd-gp43 and TsND-gp43+IL-33 groups presented reduced inflammatory cell infiltration, with intact villi and crypt structures. During the larval stage, both the T. spiralis group and the control group presented a substantial presence of Trichinella cysts within the lingual and masseter muscles, accompanied by significant infiltration of inflammatory cells. In the IL-33 group, there was a marked reduction in the number of cysts; however, the level of inflammatory cell infiltration remained consistent. Conversely, both the TsND-gp43+IL-33 and TsND-gp43 groups demonstrated significant decreases in both the number of cysts and inflammatory cell infiltration. Furthermore, compared with the TsND-gp43+IL-33 group, the TsND-gp43+IL-33 group presented more pronounced alterations in cyst morphology. These results indicate that the combination of the vaccine with the adjuvant may enhance protective efficacy.

Protective immunity against T. spiralis muscle larva infection and histopathological analysis.
The Reduction Rates of Worms at Different Stages

Discussion
LAB can act as efficient carriers of exogenous proteins for presentation to immune cells, triggering systemic and mucosal immune responses and making them excellent carriers for oral vaccines. In our study, we constructed a gene-knockout strain of L. plantarum NC8 (NC8/Δalr) carrying a plasmid that coexpressed T. spiralis TsNd and gp43 proteins for vaccination purposes and engineered the L. plantarum NC8/Δalr strain to express the murine IL-33 gene on its surface, creating a food-grade adjuvant. Following the oral immunization of BALB/c mice with an adjuvant, we evaluated the adjuvant effect of IL-33 and its role in enhancing protection against T. spiralis infection. Although a previous study (Wang et al., 2021) successfully constructed an oral vaccine utilizing recombinant L. plantarum NC8 to resist T. spiralis infection, the use of nonantibiotic recombinant L. plantarum strains as vaccine carriers may be a safer approach.
Our results showed that both the vaccine and the combined adjuvant effectively boosted the humoral immune response, especially mucosal immunity. The IgA produced B cells is effectively translocated into the intestinal lumen, resulting in elevated levels of sIgA and an increase in the population of B cells. This observation suggests that the adjuvant effectively stimulated mucosal immune responses. Additionally, the number of AWs and MLs in the intestines was reduced, effectively protecting against T. spiralis infection. Our results align with those of previous studies showing that oral immunization with the recombinant TsNd vaccine is more effective at reducing T. spiralis in AWs than is subcutaneous immunization (Liu et al., 2015; Long et al., 2014).
Th17 cells proliferate during the acute phase of T. spiralis infection and secrete the cytokine IL-17, which contributes to the contraction of ileal muscles (Lee et al., 2016). Our findings suggested that the recombinant vaccine could effectively stimulate the Th17 response. The cytokine IL-17 can facilitate early recognition of antigens, reduce the parasite burden, and effectively control host pathological damage, thereby contributing to resistance against T. spiralis invasion of the host. Furthermore, when administered in conjunction with the IL-33 adjuvant, it promotes Th2 immune polarization. This observation is consistent with previous studies indicating that IL-33 augments Th2-type cytokine responses while concurrently suppressing Th1 and Th17-type cytokine expression, which is beneficial for combating Trichuris muris infection (Humphreys et al., 2008). Moreover, IL-33 has been shown to induce goblet cell hyperplasia within the intestinal tract (Schmitz et al., 2005). Goblet cells play a crucial role in safeguarding IECs by secreting substantial quantities of mucus and immunoregulatory substances, such as secretory sIgA, which are instrumental in preventing T. spiralis infections. Histopathological assessments of the duodenum indicated that the administration of the IL-33 adjuvant in conjunction with the vaccine significantly mitigated the damage caused by T. spiralis, as evidenced by a marked decrease in the number of larval cysts present in the masseter and lingual muscles, as well as a reduction in the associated inflammatory response. Our evaluation of immune responses was conducted in a mouse model, which presents a constraint. Consequently, more research and development are still needed for adjuvants and vaccines that target swine and human immunity.
In summary, we effectively engineered a food-grade L. plantarum NC8/Δalr strain to simultaneously express two antigens of T. spiralis on its surface. This was complemented by the incorporation of a food-grade murine IL-33 adjuvant. Together, these interventions enhanced the immune response in mice and conferred partial protection against T. spiralis challenge.
Authors’ Contributions
H.-Y.L.: Writing—review and editing, writing—original draft, and data curation. D.W.: Writing—original draft, methodology, and data curation. Q.-Y.X.: Methodology. X.L.: Methodology. J.-Y.L.: Methodology and data curation. H.-B.H.: Data curation. N.W.: Methodology. C.-W.S.: Methodology. T.-X.P.: Methodology. M.-H.L.: Writing—review. D.-Y.Z.: Methodology. S.-Y.Y.: Data curation. J.-Y.G.: Data curation. G.-L.Y.: Writing—review and editing, supervision, project administration, investigation, funding acquisition, and conceptualization.
Footnotes
Funding Information
This work was supported by the National Key R&D Program of China (grant number 2023YFD1800302); the National Natural Science Foundation of China (grant numbers 32072888, U21A20261, 32202819); the China Agriculture Research System of MOF and MARA (grant number CARS-35); and the Science and Technology Development Program of Jilin Province (grant numbers 20220202057NC, 20230101346JC, 20230202080NC, YDZJ202102CXJD029, and YDZJ202301ZYTS326). The authors declare that they have no conflicts of interest related to this study. The results of this study are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation.
Disclosure Statement
No potential conflict of interest was reported by the authors.