
Abstract
This study aimed to explore the ameliorative effect of Millettia speciosa Champ. polysaccharides (MSP) on excessive exercise-induced fatigue (EF) mice. The physicochemical characterization and in vitro antioxidant activities were analyzed. The fatigue-related biochemical indexes were determined, and gut microbiota was analyzed by 16S rDNA sequencing. The results showed that MSP primarily consists of total carbohydrates, uronic acids, and proteins. It was mainly composed of glucose, arabinose, and galactose. Additionally, MSP showed good scavenging effects on free radicals. In the experiment of EF mice, MSP intervention effectively extended the pole-climbing and swimming time, significantly increased the levels of muscle and liver glycogen, blood glucose, adenosine triphosphate, glutathione peroxidase, and superoxide dismutase, while significantly decreased the levels of fatigue-related lactate dehydrogenase, creatine kinase, and urea nitrogen. Furthermore, MSP intervention reduced harmful bacteria (Helicobacter, Anaerotruncus, Erysipelatoclostridium, and Lachnospiraceae_FCS020_group), and enriched the beneficial bacteria (Lactobacillus, Alistipes, Ruminococcaceae, and Roseburia). Therefore, MSP may utilize the gut microbiota as a target to alleviate fatigue caused by excessive exercise.
Introduction
Nowadays, the number of people suffering from fatigue is gradually increasing, affecting more than 1/5 of the global population (Xu et al., 2017). Fatigue is a health hazard, sometimes even a warning sign of serious illness. Long-term vigorous exercise may trigger energy imbalance, muscle damage, and oxidative stress, ultimately leading to excessive exercise-induced fatigue (EF) (Wang et al., 2021; Zhou et al., 2023). Furthermore, fatigue can result in disturbances in gut microbiota, which has been correlated with serious risks, such as intestinal barrier dysfunction and oxidative damage (Jabeen et al., 2023; Li et al., 2023).
Polysaccharides are natural polymers and functional foods that possess antifatigue activities, such as scavenging free radicals, enhancing immune function, regulating gut microbiota, influencing the synthesis and release of neurotransmitters in the brain, and modulating glucose metabolism (Jiang et al., 2023; Luo et al., 2019). It can protect gut barrier by increasing the abundance of probiotics and maintaining interactions between tight junction proteins (Yan et al., 2024). According to report, Flammulina polysaccharides can regulate energy metabolism and the gut microbiota, improve the body’s antioxidant defenses, and exert an alleviate effect on EF (Yin et al., 2020). Accordingly, research focused on gut microbiota has become the target of polysaccharides for relieving EF.
Millettia speciosa Champ. (MS), belonging to the family Leguminosae, is a traditional Chinese medicine, which is frequently employed by people to make soup or various health foods for enhancing their physical conditions (Huang et al., 2022; Huang et al., 2020). MS polysaccharides (MSP) have significant effects on human health, such as antioxidant and immunomodulation (Yu and Liang, 2019). However, the effects of MSP in regulating EF and gut microbiota remain unclear. Therefore, MSP was extracted to analyze its physicochemical characterization, antioxidant activities, and fatigue-related biochemical factors, and the alleviative effect on EF through regulating gut microbiota was explored. The results demonstrate that MSP has the potential to act as a new prebiotic.
Materials and Reagents
MS root was purchased from the Zangyuchunzi flagship store (Guangxi, China) (#1818855807478139869). Monosaccharide standards (purity ≥98%) were obtained from Bo Rui Saccharide Biotech Co. Ltd (Jiangsu, China) (#Q18F10N80946). Trifluoroacetic acid (CAS: 76–05-1, purity ≥99%) was obtained from Thermo Fisher Scientific Inc. (Madison, USA) (#A0356762). 1,1-Diphenyl-2-picrylhydrazyl (DPPH) was purchased from Kasei Industries Co., Ltd (Tokyo, Japan) (#D4313). 2,2′-Azinobis-(3-ethylbenzthiazoline-6-sulphonate) (ABTS) was purchased from Aladdin Biochemical Technology Co., Ltd (Shanghai, China) (#A109612). The 30% hydrogen peroxide was purchased from Xilong Chemical Co., Ltd (Guangdong, China) (#UN 2014). Assay kits of glucose (GLU) (#A154-1-1), liver/muscle glycogen (LG/MG) (#A043), ATPase (#A016-2-2), blood urea nitrogen (BUN) (#C013-2-1), creatine kinase (CK) (#A022-1-1), lactate dehydrogenase (LDH) (#A020-2-2), catalase (CAT) (#A007-1-1), malondialdehyde (MDA) (#A003-1-1), glutathione peroxidase (GSH-PX) (#A005-1-2), and superoxide dismutase (SOD) (#A001-1-1) were purchased from Jiancheng Bioengineering Institute (Nanjing, China).
Preparation of MSP
The MS root was ground into powder and passed through a 40-mesh filter. Subsequently, the MS root was extracted with distilled water (95°C, 1:20 [w/v], twice for 2 h each). The combined extracts were then concentrated to 1/5 of the original volume. Protein and starch were removed with Sevag reagent and α-amylase (Li et al., 2022). Sevag reagent was removed using a rotary evaporator at 45°C, and the α-amylase was inactivated by boiling for 5 min. At 4°C, the concentrated filtrate was precipitated for 24 h with 95% ethanol (v/v). The precipitate was collected by centrifuged at 4000 rpm for 10 min, redissolved in ultra-pure water, and then dialyzed in distilled water. Eventually, MSP was obtained by freeze-drying (Filgueiras and Borges, 2022).
Characterization of MSP
Ultraviolet analysis
The ultraviolet (UV) spectrum of MSP solution (1 mg/mL) was scanned by using a UV spectrophotometer (Cary 60 spectrophotometer, Shanghai, China) within the range of 200∼400 cm−1.
Fourier transform-infrared analysis
In a dry environment under an infrared lamp, 1 mg of MSP was weighed and combined with 100 mg KBr. The sample was flattened into thin sheets and scanned by Fourier transform-infrared (FT-IR) spectra (Nicolet iS5 FT-IR, Thermo Electron Co., Madison, USA) from 4000 to ∼400 cm−1 (Gómez-Ordóñez and Rupérez, 2011).
Scanning electron microscope analysis
The microstructural characteristics of MSP were imaged with a scanning electron microscope (SEM) (JSM6701F, Japan). The sample was sprayed with gold and examined at an accelerating voltage of 3 kV at 500-fold magnification (Wang et al., 2022a).
X-ray diffraction analysis
An X-ray diffractometer (D8 ADVANCE, BRUKER, Germany), operating at 40 kV and 40 mA, was used to analyze MSP. Data were collected from 2θ of 5∼70° by sequential scanning with a step width of 0.02° and a scanning speed of 10◦/min (Ren and Liu, 2020).
Thermogravimetric analysis
The thermal stability of MSP was assessed by thermogravimetric analysis (TGA 4000, USA). Measurements were carried out in an air atmosphere, and the temperature ranged from 30°C to 600°C at a heating rate of 10°C/min.
Molecular weight analysis
High-performance gel permeation chromatography (HPGPC) on a Mass Lynx-DEFAULT HPLC system (Shimadzu, UK) fitted with a RID-10A detector was used to measure the molecular weight of MSP (Wu et al., 2022). A 2 mg/mL sample solution was filtered applied to a BRT105-103-101 tandem gel columns (8 mm × 300 mm), at a flow rate of 0.8 mL/min at 40°C. The mobile phase contained 25 μL of 0.2 M NaCl.
Chemical compositions analysis
The Bradford, m-hydroxydiphenyl, and phenol-sulfuric acid methods were used to determine the contents of total carbohydrates, uronic acid, and protein, respectively (Filgueiras and Borges, 2022; Ibarz et al., 2006; Masuko et al., 2005). An ICS5000 high-performance anion-exchange chromatography system with a pulsed ampere detector was used to analyze the monosaccharide content of MSP. Sixteen types of monosaccharides were used as standards. The sample was hydrolyzed in an ampoule bottle at 120°C for 2 h with 2 mL trifluoroacetic acid (TFA 3 M). After the acid hydrolysis solution was nitrogen-blown to dryness, 5 mL of water was added. The supernatant was detected by the Mass Lynx-DEFAULT HPLC system (Shimadzu, UK) with Dionex CarbopacTM PA20 columns (3 mm × 150 mm). The injection volume was 25 μL, and the column temperature was 30°C (Xie et al., 2016).
Antioxidant activity in vitro
The antioxidant ability of MSP was assessed by hydroxyl radical (·OH), ABTS+·, and DPPH· scavenging assays. The concentration of MSP and ascorbic acid was from 0.1 to 5.0 mg/mL (n = 3) (Qiao et al., 2022; Xian et al., 2023).
A total of 50 μL of MSP solution was mixed with 1 mL of ABTS+· (7 mmol/L) solution. The solution was shaken thoroughly and reacted in the dark for 20 min, and the absorbance at 734 nm was measured.
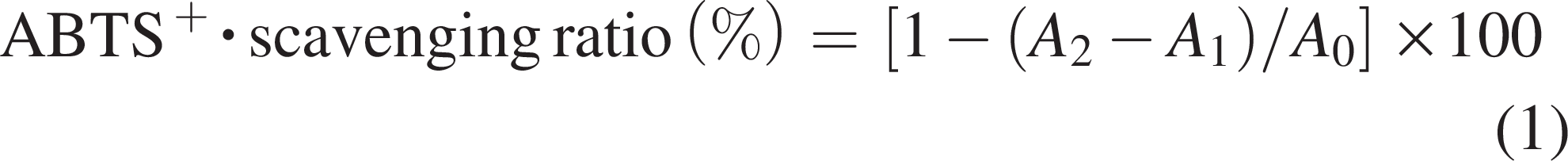
Distilled water in the ABTS+· solution, samples in the phosphate buffer saline (PBS) solution, and samples in the ABTS+· solution were represented by the absorbances A0, A1, and A2.
Note that 0.5 mL DPPH-anhydrate ethanol solution (v/v) and 0.5 mL samples were combined, and the mixture was allowed to sit dark for 30 min. At 517 nm, the absorbance was measured.
Distilled water in the DPPH· solution, samples in the anhydrate ethanol solution, and samples in the DPPH· solution were represented by the absorbances A0, A1, and A2.
A total of 0.1 mL sample, 2 mL 6 mM FeSO4, and 2 mL 6 mM H2O2 were mixed successively. The mixture was left at room temperature for 10 min. Subsequently, 2 mL 6 mM salicylic acid-anhydrous ethanol was mixed, which was then incubated in water bath at 50°C for 30 min. At 510 nm, the absorbance was measured.
Distilled water in the H2O2 solution, samples in the distilled water, and samples in the H2O2 solution were represented by the absorbances A0, A1, and A2.
Animals
Sixty 4-week-old male Institute of cancer research (ICR) mice (20–22 g; Spife Biotechnology Co., Ltd Beijing) were kept in a suitable environment (25 ± 2°C, 50 ± 10% relative humidity, with a 12-h light and 12-h dark cycle). The mice had free access to food and water for 1 week for adaptation and were divided into six group (10 mice per group). The animal designs and procedures employed in this study were approved by the Animal Care and Use Department of Jiangxi Agricultural University, and were conducted in accordance with the guidelines for the administration of experiment (Approval No. JXAUA01). The Rest control (R-CON) and Training control (T-CON) groups were gavaged of 0.1 mL of distilled water, while the MSP-L (200 mg/kg), MSP-M (400 mg/kg), MSP-H (800 mg/kg), and 15 mg/kg taurine (Tau) (Wang et al., 2022b) groups were treated accordingly. All mice were continuously administered for 30 days. Food intake and body weight were measured daily.
Pole test
Mice were trained to climb poles once a week, and each training session lasted 20 min. On day 25, all mice underwent pole test. After three repetitions of placing the dropped mice back the pole, the experiment came to an end. The time of the third fall for each mouse was recorded.
Weight-loaded swimming test
Weight-loaded swimming was conducted once a week. A lead wire weighting 10% of the mice’s body weight was attached to the tail root, after which the mice were immediately placed in a swimming pool with a water depth of 40 ± 1 cm and a temperature of 25 ± 1°C. The training was terminated each time when the mouse nose tip was immersed in water for more than 10 s. On day 30, after gavage, the mice were allowed to rest for 30 min before undergoing the weight-loaded swimming test. The swimming time was recorded, and the mice were dried rapidly. Subsequently, all the mice were executed by cervical dislocation. To extract the plasma, blood was drawn from the eye vein and centrifuged for 10 min at 4°C 3500 rpm. The heart, kidneys, liver, and gastrocnemius muscle were weighed and recorded. All samples were rapidly stored the −80°C.
Biochemical indicators
Assay kits were used to determine the levels of BUN, CK, GLU, LDH, GSH-PX, SOD, MDA, and CAT in plasma. Simultaneously, LG, Ca2+·Mg2+ ATPase, and Na+·K+ ATPase in the liver, as well as MG in the gastrocnemius muscle were determined.
Gut microbiota analysis by 16S rDNA gene sequencing
The cecum contents of 5 mice in each group were randomly selected to extract total genomic DNA using E.Z.N.A.® Soil DNA Kit (Omega Bio-Tek, Norcross, GA, USA), and the DNA was quantified with Nanodrop. Note that 1.2% agarose gel electrophoresis was used to assess the quality of the DNA extraction. The V3-V4 regions of 16S rDNA were selected for amplification and then sequenced by Nomidaixie Biotechnology Co., Ltd (Jiangsu, China). The Genes Cloud Platform was used to analyze all data (https://www.biodeep.cn).
Statistical analysis
All determinations were carried out in triplicate, and the results were presented as mean ± standard error of the mean. The results were processed using SPSS 26 software (IBM Corporation, New York, USA). Post hoc Duncan’s test and one-way Analysis of variance (ANOVA) were used to detect significant differences at p < 0.05.
Results
Characterization of MSP
MSP had total carbohydrate, uronic acid, and protein, respectively (Table 1). The UV spectra showed that a peak at 280 nm was weak, indicating that MSP contained trace of protein (Fig. 1A). The FT-IR spectra (Fig. 1B) showed the distinctive absorption regions of MSP. The peak at 3415.60 cm−1 was attributed to the hydroxyl (-OH) stretching vibration (Wang et al., 2016). The peaks at 2923.88 cm−1 and 1401.36 cm−1 were assigned to the stretching and variable angle vibrations of C-H, respectively (Wu et al., 2021). The peak at 1629.69 cm−1 belonged to the asymmetrical vibration absorption of ester group (COO−) (Wu et al., 2021). Peaks at 1147.11 cm−1, 1019.79 cm−1, and 1026.89 cm−1 indicated that pyranose configuration existed (Liu et al., 2020). As shown in Figure 1C, the SEM result demonstrated that the morphological characteristics of MSP appeared sparse, coarse, and irregularly flaky. The X-ray diffraction result indicated that the crystallinity of MSP was 8.3%. The degree of crystallization was low, which made it unable to form single crystal (Fig. 1D). The residual weight percentage of MSP at 600°C is 33.88%, which indicated the caloric stability was good (Fig. 1E).
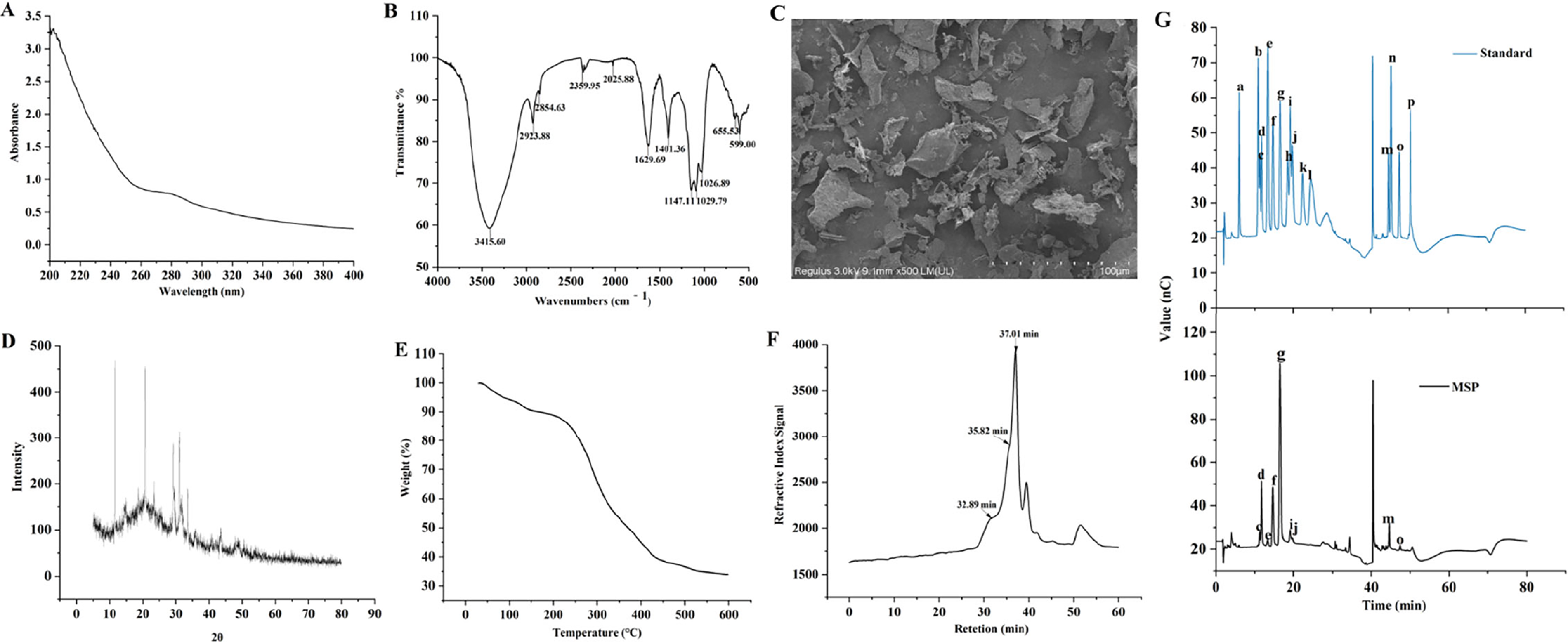
The structural characteristics, physical, and chemical properties of Millettia speciosa Champ. polysaccharides (MSP).
Chemical Properties of Millettia speciosa Champ. Polysaccharides

Glc, glucose; Ara, arabinose; Gal, galactose; GalA, galacturonic acid; Rha, rhamnose; Xyl, xylose; GlcA, glucuronic acid; Man, mannose; GlcN, glucosamine hydrochloride.
The HPGPC plot of MSP is shown in Figure 1F. The polysaccharide exhibited three peaks (32.89 min, 35.82 min, and 37.01 min). Table 1 shows the corresponding molecular weights.
The ion chromatogram of MSP is shown in Figure 1G. In Table 1, it indicated that MSP was a heteropolysaccharide, which mainly composed of glucose, arabinose and galactose, and the molar percentage ratio.
In vitro antioxidant activity of MSP
In Figure 2A–C, the ability of MSP to scavenge ABTS+·, DPPH·, and ·OH radicals showed an upward trend, with the IC50 values of 0.25, 0.20, and 8.18 mg/mL, respectively. These results demonstrated that MSP plays an antioxidant role.
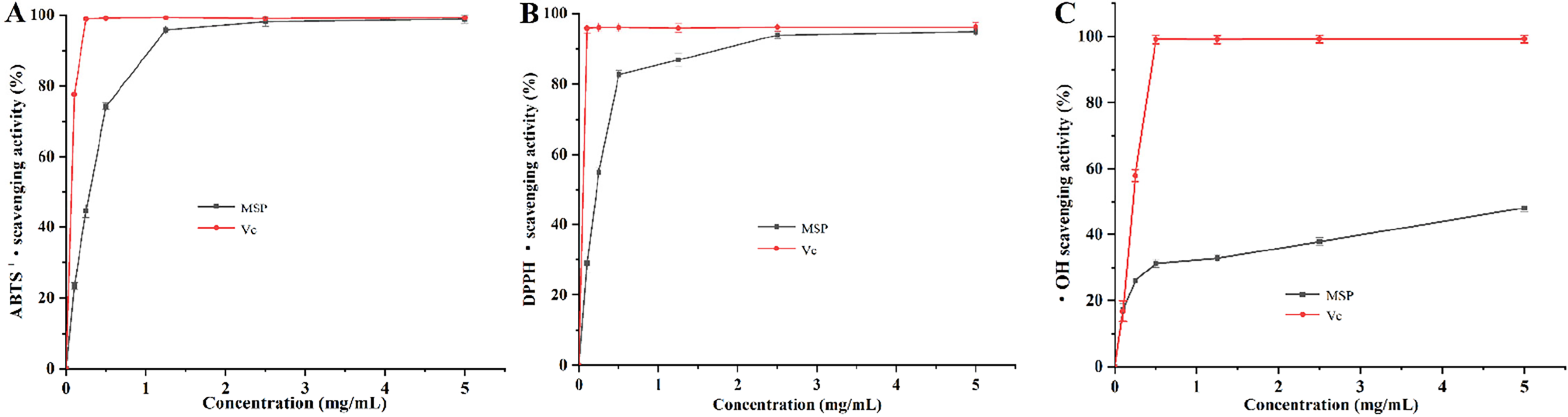
Antioxidant capacities assay of Millettia speciosa Champ. polysaccharides (MSP) in vitro (n = 3).
Effect of MSP on exercise endurance in mice
The weight-loaded swimming test and pole test were commonly used to reflect symptoms of excessive EF. The animal experimental process is shown in Figure 3A. In Figure 3B–E, the food intake and organ weights of the mice did not exhibit significant differences (p > 0.05), indicating that MSP had almost no toxic side effects. In Figure 3F and G, compared with T-CON, MSP treatment significantly increased the pole test time and swimming time (p < 0.05).

Effect of Millettia speciosa Champ. polysaccharides (MSP) on excessive exercise-induced fatigue mice.
Effect of MSP on fatigue-related biochemical indices in mice
In Figure 4A–F, compared with T-CON, MSP treatment for 30 days led to a significantly increased levels of biochemical indices associated with GLU, LG, MG, Na+·K+ ATPase, and Ca2+·Mg2+ ATPase, while significantly reduced with BUN, CK, and LDH (Fig. 4G, H) (p < 0.05). In Figure 4I and J, compared with T-CON, the MSP treatment mice showed a significantly increased level of GSH-PX and SOD (p < 0.05). In addition, after MSP treatment, no discernible changes were observed on plasma MDA and CAT (p > 0.05).
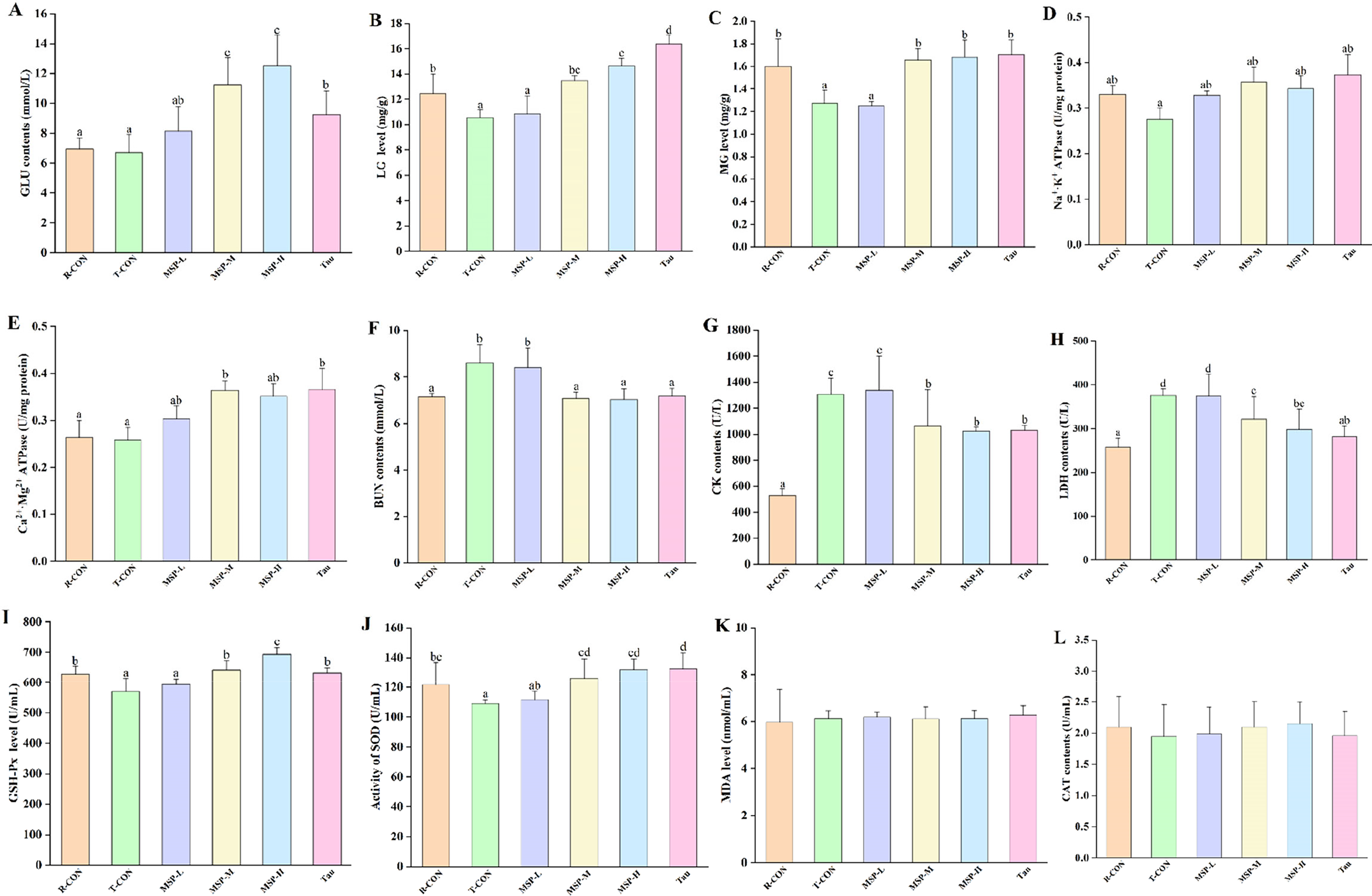
Effect of Millettia speciosa Champ. polysaccharides (MSP) on fatigue-related biochemical (energy metabolism, metabolic waste, muscle damage, and oxidative stress) indicators (n = 9).
Impact of MSP on gut microbiota diversity
In Figure 5A and B, both the rarefaction curve and rank abundance curve gradually flattened, indicating that the sequencing depth was sufficient. In Figure 5C and D, when compared with T-CON, there was no discernible difference in the Chao1 and Shannon index of MSP (p > 0.05). The Venn plot was used to investigate species differences. The results indicated that no significant differences in unique operational taxonomic units (OTUs) were observed among the groups (Fig. 5E). In Figure 5F and G the nonmetric multidimensional scaling and the principal coordinates analysis are displayed. There was a significant difference in the distance between T-CON and MSP-H. In Figure 5H, the cluster heat map showed that the abundance of the top 50 microbiota was significantly altered. Compared with the T-CON, MSP-H significantly restored 40 microbiota taxa, representing the highest number of restored taxa among all treatments.

Effect of Millettia speciosa Champ. polysaccharides (MSP) on gut microbiota diversity and composition (n = 5).
MSP ameliorated EF-associated gut microbiota dysbiosis
In Figure 6A, four primary bacteria at the phylum level are shown, including Bacteroidetes, Firmicutes, Campylobacterota, and Desulfobacterota. In Figure 6B, compared with T-CON, the ratio of Firmicutes/Bacteroidetes decreased in MSP treatment, while the relative abundance of Proteobacteria decreased in MSP treatment (Fig. 6C) (p < 0.05). In Figure 6D, compared with T-CON, MSP treatment indicated an increase in the abundance of Muribaculaceae and Alistipes. In Figure 6E–G, compared with T-CON, MSP-H significantly increased the relative abundance of Lactobacillus, Desulfovibrio, and Roseburia (p < 0.05). In Figure 6H, Desulfovibrio, Ruminococcaceae, Enterorhabdus, Unclassified bacteria group-5 (UCG-5), and Unclassified bacteria group-10 (UCG-10) were significantly positively correlated with swimming time, pole test time, LG, MG, GLU, SOD, and GSH-PX. Bifidobacterium was negatively correlated with CK, LDH, and BUN (p < 0.05). Besides, Alloprevotella, Clostridia_Unclassified bacteria group-014 (Clostridia_UCG-014), Anaerotruncus, Erysipelatoclostridium, [Eubacterium]_coprostanoligenes_group, and ASF356 were positively correlated with CK, LDH, and BUN (p < 0.05).

Influences of Millettia speciosa Champ. polysaccharides (MSP) on gut microbiota composition (n = 5).
Discussion
EF can lead to energy consumption, muscle injury, oxidative imbalance, and microbiota dysbiosis (Liu et al., 2024). Previous studies have shown that the levels of GLU, LG, MG, ATPase, CK, BUN, LDH, GLU, SOD, and GSH-PX are closely related to EF (Chen et al., 2022). It is well known that EF causes decreased levels of GLU, LG, MG, ATPase, SOD, and GSH-PX, as well as increased levels of CK, BUN, and LDH, which pose a health threat (Gao et al., 2018; Yang et al., 2020). Researches have indicated that polysaccharides can supply energy, reduce muscle damage, and provide antioxidants for antifatigue effects (Cai et al., 2021; Liu et al., 2022). In this work, compared with T-CON, MSP markedly increased the levels of energy-related substances (GLU, LG, MG, Na+·K+ ATPase, Ca2+·Mg2+ATPase) and two antioxidant enzymes (GSH-PX and SOD), and significantly reduced the quantities of muscle damage-related substances (LDH, CK, BUN), which may facilitate its anti-EF effects.
Exhaustive exercise has been reported to induce gut barrier damage and decrease microbial abundance (Zhou et al., 2021). Natural polysaccharides possess unique advantages in the prevention of fatigue-related diseases due to the remodeling of gut microbiota (Lei et al., 2024; Wei et al., 2023). In this study, the MSP treatment ameliorated the dysbiosis of the gut microbiota, which in turn alleviated EF. EF can increase the Firmicutes/Bacteroidetes ratio (Frémont et al., 2013). Moreover, the MSP treatment significantly decreased the ratio of Firmicutes/Bacteroidetes. Proteobacteria disrupted metabolism by causing intestinal inflammation and an imbalance of the oxidative system (Morgan et al., 2012). Desulfovibrio is an anaerobic bacterium positively correlated with certain energy metabolism substances, consistent with Xiao et al. (2020). Lactobacillus can increase branched-chain amino acids and produce short-chain fatty acids (SCFAs) to maintain muscle energy homeostasis, thereby alleviating fatigue (Jäger et al., 2020). Bacteroidetes and Roseburia can produce butyric acid, which provides energy to the intestinal epithelial cells to fight against fatigue (Xing et al., 2018). Alistipes are Gram-negative anaerobes that produce SCFAs, which provide energy and help mitigate fatigue (Wu et al., 2019). In this study, MSP reversed the loss of beneficial microbiota, including Desulfovibrio, Ruminococcaceae, Lactobacillus, Roseburia, Alistipes, and Muribaculaceae, and decreased potentially harmful bacteria, such as Helicobacter, Clostridia_UCG-014, Anaerotruncus, Erysipelatoclostridium, and Lachnospiraceae_FCS020_group, thereby reshaping the gut microbiota. Restoring a balanced composition of the gut microbiota can lead to significant improvements in fatigue symptoms (Liu et al., 2021). For example, the gut microbiota can influence energy homeostasis by modulating adenosine monophosphate-activated (AMP-activated) protein kinase activity and attenuating the immune response to lipid oxidation (Yu et al., 2022). SCFAs are metabolites produced by the gut microbiota that promote fatigue relief through various mechanisms. They regulate oxidoreductases, enhance the activation of the nuclear factor-erythroid 2-related factor 2 pathway, repair intestinal barrier damage, and reduce oxidative stress (Liu et al., 2021). The aforementioned findings indicated that gut microbiota plays a crucial role in alleviating fatigue by enhancing energy production and mitigating oxidative damage induced by reactive oxygen species. These results confirmed that MSP could restore the composition of the gut microbiota to a certain extent and might be used as a promising prebiotic.
In sum, fatigue symptoms may be ameliorated and health promoted by MSP by modulating gut microbiota, enhancing energy metabolism, reducing oxidative stress, and lowing the levels of tissue damage indicators. However, to achieve broader applications, further research on human clinical trials is necessary.
Conclusions
This study demonstrated that 30-day MSP treatment exhibited a relieving effect on EF mice. In conclusion, MSP may enhance exercise tolerance in mice by improving energy metabolites, muscle injury, antioxidant activities, as well as influencing the gut microbiota. Therefore, MSP has broad application prospects as a potential prebiotic in the fields of food and medicine. It has the potential to be utilized in health supplements and food additives, thereby contributing positively to human health.
Authors’ Contributions
J.Z.: Research design, data curation, investigation, formal analysis, and writing—original draft preparation. W.Z.: Formal analysis. L.Y.: Writing—review and editing. Q.Y.: Project administration. W.W.: Designed the research. X.H.: Investigation. J.L.: Designed the research, writing—review and editing, and project administration. G.Z.: Funding acquisition and resources, research design, writing—review and editing, project administration, and supervision. All authors reviewed and approved the final version of the article.
Footnotes
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 82060165).
Disclosure Statement
The authors declare no competing interests.