
Abstract
Enterococcus faecium (Ef) is a common microorganism in the gastrointestinal tract, recognized for its beneficial effects on human and animal health. It plays a vital role in promoting intestinal microbial balance and enhancing gut barrier function. A key feature of Ef extracellular vesicles (EfEVs) is their ability to specifically target sites within the intestine, leading us to hypothesize that EfEVs function as an important active component of Ef in intestinal microenvironment regulation. This study aimed to explore the impact of EfEV supplementation on the intestinal barrier function and microbiota composition in piglets. In vitro intestinal porcine epithelial cell J2 (IPEC-J2) cell culture experiments have shown that supplementation with EfEV significantly enhanced cell viability, increased the activity of the antioxidant enzyme, promoted cell proliferation, and inhibited autophagy. In vitro, fecal culture experiments demonstrated significant increases in OD630 and the contents of acetic aid, butyric acid, and isovaleric acid, and decreases in valeric acid following EfEV supplementation. Furthermore, EfEV supplementation altered the diversity and composition of the intestinal microbiota. It significantly reduced levels of Escherichia-Shigella at the family level, while increasing the abundance of beneficial microorganisms, including [Eubacterium]_eligens_group, unidentified_Mitochondria, Lachnoclostridium. In conclusion, EfEV plays a pivotal role in inhibiting pathogens growth, enhancing the production of metabolites such as acetic acid and butyrate acid, providing energy for microorganisms, and protecting the intestinal barrier.
Introduction
The intestine is the primary site for nutrient absorption and digestion, as well as serving as a crucial barrier for overall health (Huo et al., 2022). In pig production, the weaning period is considered one of the most critical stages. Weaning stress often leads to alterations in the morphology and function of the small intestine, disrupting digestion and absorption, compromising intestinal barrier function, and causing microbiota dysbiosis (Li et al., 2022; Shao et al., 2023). These disruptions ultimately result in reduced feed intake, increased rates of diarrhea, and stunted growth; therefore, special attention must be given to the gut health of piglets during weaning to ensure they smoothly navigate this critical stage.
Probiotics, live, viable microorganisms, have shown promise in enhancing intestinal microbiota diversity, promoting the proliferation of beneficial bacteria, and aiding in resistance against pathogenic microbial infections (Hu et al., 2018; Tang et al., 2020). Enterococcus faecium (Ef), a common intestinal bacterium found in both humans and animals, is one of the probiotics frequently used in pig production. Ef plays a crucial role in maintaining the balance of piglet intestinal microbiota by promoting the development of small intestinal villi, increasing nutrient absorption, and improving disease resistance (Bednorz et al., 2013). It also reduces oxidative stress and paracellular permeability in IPEC-J2 cells and inhibits the adhesion of Salmonella typhimurium and Escherichia coli (Palkovicsne Pezsa et al., 2022).
Extracellular vesicles (EVs) are integral to intercellular communication, facilitating the transfer of bioactive molecules such as proteins, nucleic acids, and lipids, thereby influencing physiological and pathological processes in recipient cells (Komuro et al., 2022; Wu et al., 2022). EVs can also modulate the intestinal environment by inhibiting the proliferation of pathogenic bacteria and altering microbial composition through their metabolic products (Liang et al., 2022; Rodovalho et al., 2020).
This study investigated the effects of EfEV on the viability of IPEC-J2 cells, and microbiota flora in the fecal cultures of weaned piglets. The findings provided valuable insights into the potential application of bacterial EV as modulators of intestinal health and microbiota in pig production.
Materials and Methods
Culture of bacteria
Ef used in this study was obtained from our laboratory. The bacteria were cultured in the Man Rogosa and Sharpe medium (Supplementary Table S1; Luo et al., 2024b). The culture was incubated at 37°C with shaking at 200 rpm for 14–16 h under aerobic conditions until the bacterial growth reached an OD600 of 0.4–0.6.
EfEV isolation and identification
EfEV isolation and identification were performed as previously described (Luo et al., 2024b). Briefly, the culture medium was centrifuged at 4°C and 2000 × g for 10 min, and the supernatant was collected and centrifuged again at 4°C and 10,000 × g for 30 min. The resulting supernatant was then ultracentrifuged at 4°C and 120,000 × g for 90 min to obtain the EfEV pellet. The pellet was resuspended in phosphate buffer saline (PBS) and ultracentrifuged again at 4°C and 120,000 × g for 90 min. The enriched pellet obtained was then purified EfEV, which was then resuspended in PBS and stored at −80°C.
Cell and cell culture
IPEC-J2 cell culture was performed as previously described (Luo et al., 2024a). IPEC-J2 cells were grown in DMEM/F-12 (GIBCO, China) supplemented with 10% (v/v) fetal bovine serum (GIBCO, New Zealand), 50 IU/mL penicillin (GIBCO, China), and 50 μg/mL streptomycin (GIBCO, China). The cells were maintained at 37°C in a humidified incubator under 95% air and 5% CO2.
Cell viability assay
Cell survival assay was performed using the Thiazolyl blue tetrazolium bromide (MTT) method. Briefly, IPEC-J2 cells were cultured in 96-well plates at a density of 2 × 104 cells/well with corresponding treatment. After 12 h, the cells were treated with 0, 0.001, 0.01, 0.1, 1, and 10 μg EV protein/mL EfEV, respectively. After 12 h, the cell culture supernatants were removed and replaced with 20 μL of MTT solution (5 mg/mL in PBS, Sangon Biotechnology Co., LTD., Shanghai, China), followed by incubation at 37°C for 4 h. Afterward, the medium was removed and 150 μL of DMSO was added to each well. The absorbance at 490 nm was measured using a microplate reader (Tecan, Männedorf, Switzerland) after shaking the plates for 10 min.
RNA extraction and real-time PCR (polymerase chain reaction)
Total RNA was isolated using Trizol reagent (Invitrogen), and its concentration was determined with a NanoDrop spectrophotometer (Thermo Fisher Scientific). Subsequently, 1 μg of RNA was transcribed into cDNA using the RT EasyTM IIcDNA Synthesis kit (Foregene, Chengdu, China). Real-time PCR was carried out using SYBR Green Supermix (Foregene, Chengdu, China) on the quantStudio3 (Applied Biosystems) machine. The specific primer sequences used are listed in Supplementary Table S2. Data analysis was conducted using the 2−ΔΔCt method, with the ACTB gene serving as the reference gene.
Cells antioxidant levels
Added 500 μL of pre-cooled PBS to the cell plate. Used a cell scraper to carefully detach the adherent cells, and transferred the cell suspension to a homogenization tube containing homogenization beads. Homogenized the sample for approximately 40 s. After homogenization, centrifuged the mixture and collected the supernatant for antioxidant index measurement. Next, measured the total antioxidant capacity (T-AOC), lactate dehydrogenase (LDH), superoxide dismutase (SOD), and glutathione peroxidase (GSH-Px) using the respective kits according to the instructions provided by Nanjing Jiancheng Bioengineering Institute.
Defined base medium
The defined growth medium comprised the components outlined in Supplementary Table S3 (Adamberg and Adamberg, 2018; Luo et al., 2024a).
Method for fecal culture
Collected fecal samples from 11 weaned piglets (32 days old, 4 days post-weaning) were kept at −20°C. The frozen samples were delivered to the laboratory within 12 h. Upon arrival, the fecal samples were diluted fivefold in sterile anaerobic 1 × PBS, then the dilution was supplemented with freshly prepared, filter-sterilized cysteine hydrochloride (0.05% in the final solution) (Adamberg and Adamberg, 2018; Luo et al., 2024a). In the EfEV group, the fecal culture was supplemented with a dosage of EfEV (1 μg EV protein/mL), while the control group received an equal volume of PBS.
Measurement of pH and OD630 value in fecal culture
The pH was assessed using a pH meter of the sample in 1 mL fecal culture. Then, the fecal culture was centrifuged at 9600 r/min for 5 min. After centrifugation, the supernatant was discarded and the bacterial pellet was resuspended in PBS. Finally, an appropriate volume of sample was added to a 96-well plate and the absorbance at 630 nm was measured using a microplate reader.
Microbiota analysis
After 12 h, fecal culture were collected and stored at −80°C until usage. The microbiota analysis was conducted using 16S rRNA sequencing by Novogene Technology Co., Ltd. (Beijing, China). Similar sequences were clustered into the same operational taxonomy unit (OTU) with a 97% sequence identity. To evaluate alpha diversity in each sample, the Chao index, Sobs index, and Shannon index were calculated. In addition, the chao1 and abundance-based coverage estimator (ACE) indices were used to assess the α-diversity among groups. Cluster analysis, based on Euclidean distance, was performed using the relative abundances of all OTUs. Finally, the composition of fecal bacteria was analyzed at the phylum, family, and genus levels.
Determination of metabolites
The main short-chain fatty acids (SCFAs) were determined by gas chromatography (Abdel-Rahman et al., 2020; Qi et al., 2020). The chromatographic conditions were as follows: a DB-FFAP (60 m × 0.25 mm × 0.25 μm) column was used, with a flow rate of 0.8 mL/min and an injection volume of 1 μL. The temperature program was initiated at 60°C with a 2 min hold, followed by an increase to 220°C at a rate of 20°C/min, and then held for 3.5 min. Detection was achieved using a flame ionization detector set at 250°C.
Statistical analyses
Results were analyzed using an unpaired, two-tailed Student’s t-test with SPSS 20 to calculate a p value. Significance was declared at p ≤ 0.05. Results are presented as mean ± standard deviation.
Results
Effect of different concentrations of EfEV on the viability of IPEC-J2 cells
As shown in Figure 1A, the effect of various concentrations of EfEV on the viability of IPEC-J2 cells was assessed. In the concentration range of 0.001∼1 μg EV protein/mL, a positive correlation was observed between EfEV concentration and cell viability. Specifically, cell viability increased significantly at concentrations of 0.01–0.1 μg EV protein/mL compared to the control group (p < 0.05), with the highest increase occurring at 1 μg EV protein/mL. However, at a concentration of 10 μg EV protein/mL, cell viability decreased compared to the control group, though this reduction was not statistically significant (p > 0.05).
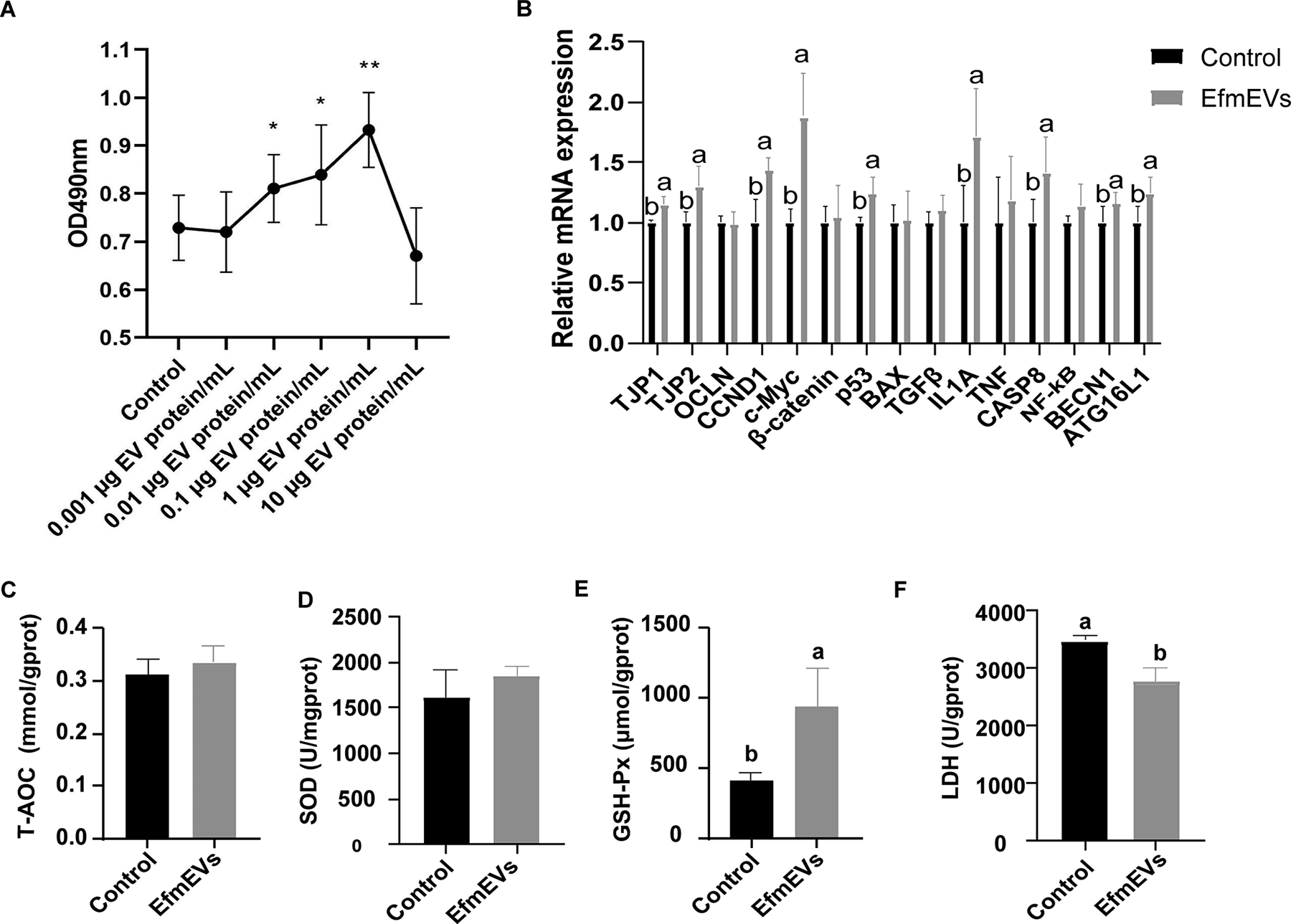
Effects of EfEV on IPEC-J2 cells.
Effect of EfEV on mRNA expression in IPEC-J2 cells
As shown in Figure 1B, EfEV treatment resulted in increased relative expression levels of TJP1, TJP2, CCND1, c-Myc, p53, IL1A, CASP8, BECN1, and ATG16L1 compared to the control group (p < 0.05).
Effects of EfEV on the antioxidant capacity of IPEC-J2 cells
As shown in Figure 1C–F, EfEV treatment resulted in a significant increase in GSH-Px level (p < 0.05) (Fig. 1E), and a significant decrease in the LDH level compared to the control group (p < 0.05) (Fig. 1F).
Effect of EfEV on fecal cultures of pH and OD630
Feces from 32-day-old weaned piglets were collected and cultured with the supplementation of EfEV to study the effect of EfEV on the fecal microbiota (Table 1). EfEV supplementation significantly increased in OD630 (p < 0.05).
Effect of EfEV Supplementation on pH and OD630 in Piglets Fecal Culture
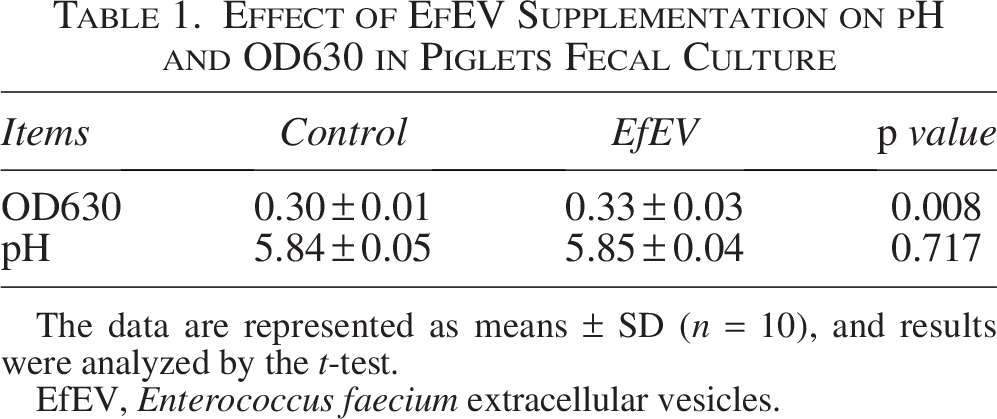
The data are represented as means ± SD (n = 10), and results were analyzed by the t-test.
EfEV, Enterococcus faecium extracellular vesicles.
Effect of EfEV on volatile fatty acid content in fecal cultures
The effects of EfEV on the volatile fatty acids in the fecal cultures of piglets are shown in Table 2. Compared to the control group, EfEV supplementation increased the levels of acetic acid, butyric acid, and isovaleric acid (p < 0.05) and decreased the level of valeric acid (p < 0.05).
Effect of EfEV Supplementation on the Concentrations of SCFAs in Piglets Fecal Culture
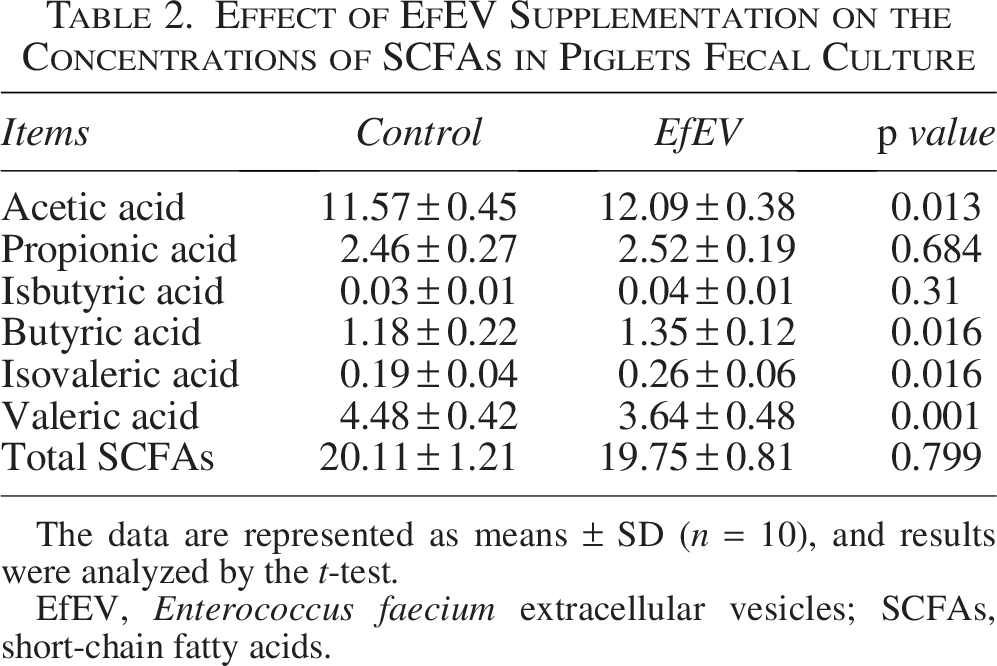
The data are represented as means ± SD (n = 10), and results were analyzed by the t-test.
EfEV, Enterococcus faecium extracellular vesicles; SCFAs, short-chain fatty acids.
Effect of EfEV on the microbiota of fecal cultures
The Venn diagram in Figure 2A showed that the control group contained 511 OTUs, including 80 unique OTUs, while the EfEV group had 532 OTUs, with 101 unique OTUs. Alpha diversity comparisons were shown in Figure 2B, demonstrating that supplementation with EfEV significantly increased both the Simpson index and Chao1 index (p < 0.05). At the OTU level, PCA analysis indicated significant differences in bacterial composition between the two groups (Fig. 2C).
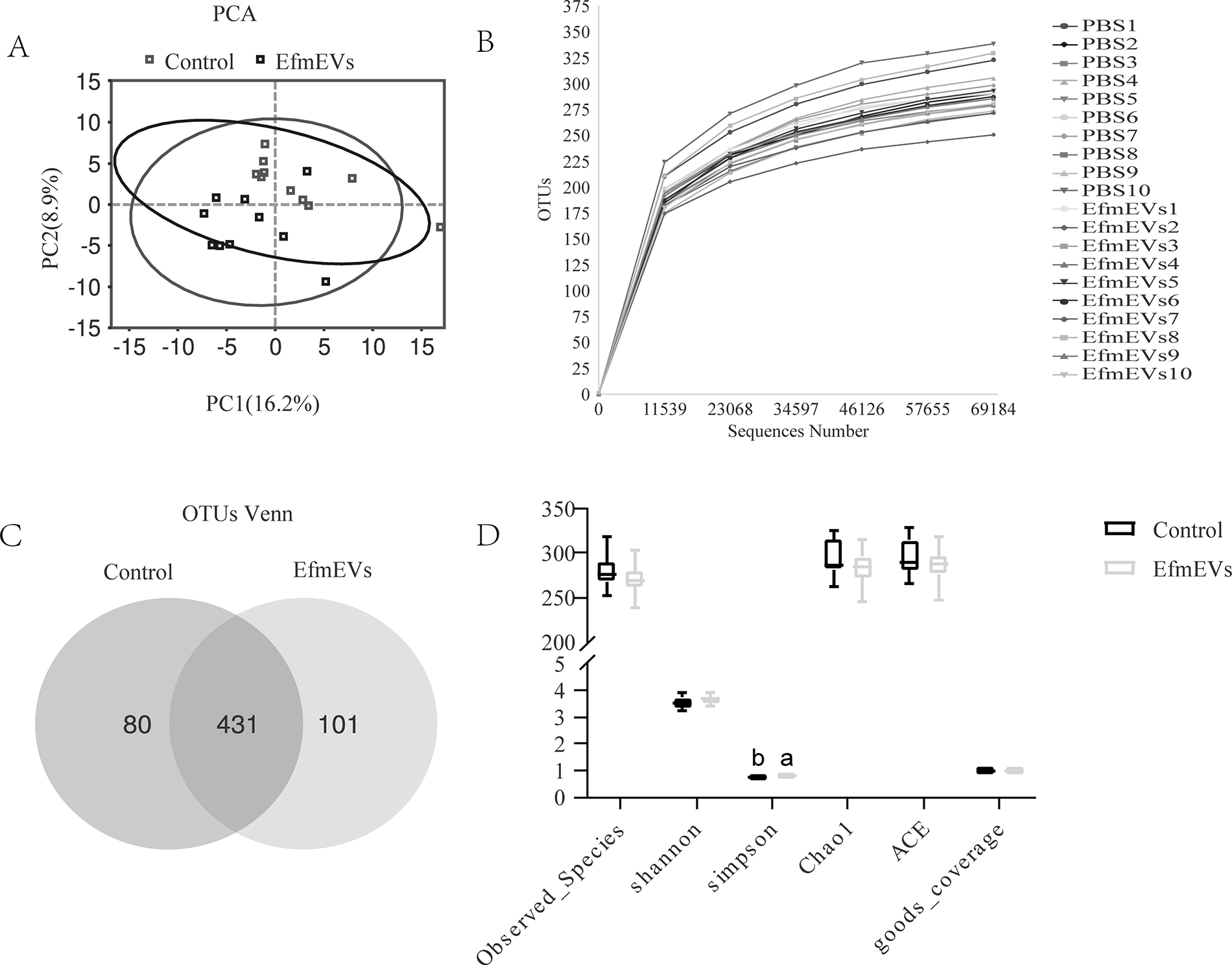
Effect of EfEV on richness and diversity of fecal microbiotas in vitro fecal culture of piglets.
Based on the species annotation, the top 10 microorganisms in terms of relative abundance at the phylum, family, and genus levels were identified in the fecal microbiota. The microbial abundance charts are shown in Figure 3A–C. At the phylum level, the dominant microbiota in both the control group and EfEV group were Proteobacteria, Bacteroidetes, and Firmicutes. Compared to the control group, the EfEV group significantly increased the relative abundances of Firmicutes and Cyanobacteria, and deseaing the relative abundances of Proteobacteria (Fig. 3D). At the family level, following the interventions, the relative abundance of [Eubacterium]_eligens_group, Unidentified_Chloroplast, unidentified_Mitochondria, Lachnoclostridium, and Oribacterium was significantly increased, while that of Escherichia-Shigella, Enterococcus, unidentified_Chloroplast, Mitsuokella, Megasphaera, Lachnosiraceae_NK3A20_group, Sutterella, Erysipelotrichaceae_UCG-009, Streptococcus, Ligilactobacillus, Erysipelatoclostridium, and Butyricicoccus was decreased in the EfEV group (Fig. 3E). At the genus level, the abundance of the Schrenkiella_parvula increased significantly after supplementation with EfEV, while the abundance of Escherihia_coli, Enteroccus_faecium, Megasphaera_elsdenii, Sutterella_parvirubra, Lactobacillus_salivarius, and Erysipelatostridium_ramosum decreased (Fig. 3F).
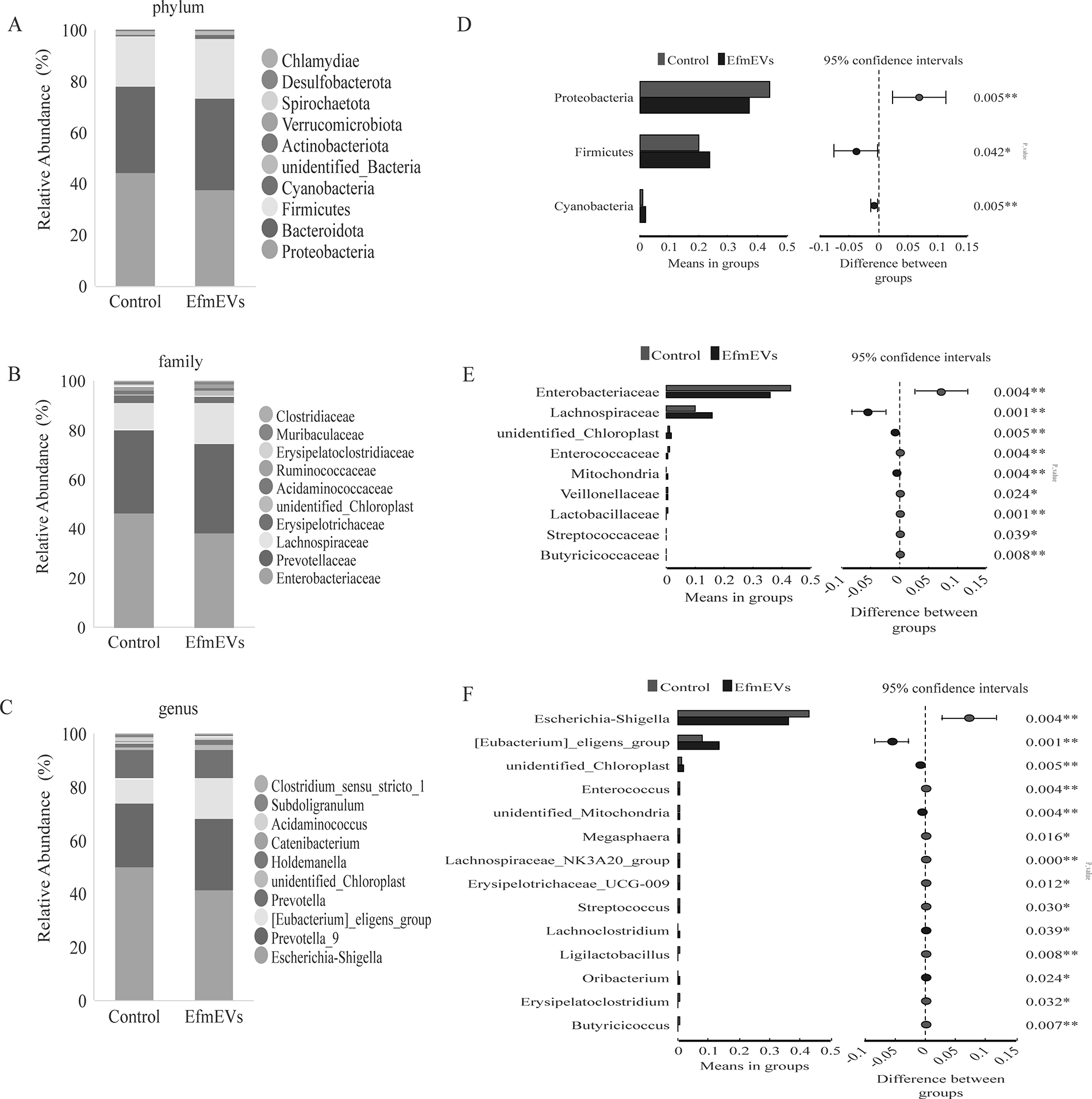
Effect of EfEV on the intestinal microbiota in vitro fecal culture of piglets.
Discussion
The stability of the intestine microecosystem is essential for the animals health, as it prevents pathogen invasion and enhances nutrient digestion and absorption (Ivanov et al., 2006). Ef, a probiotic found in the gastrointestinal tract, has been used to promote human and animal health by improving gut barrier function and microbial balance (He et al., 2021; Xu et al., 2022). Our previous studies demonstrated that EfEV exhibits a similar function in gastric injury protection as Ef itself (Luo et al., 2024b). In this study, we investigated the effects of EfEV on the luminal environment of the porcine intestine by using IPEC-J2 cells and fecal culture models. The results showed that EfEV improved cell permeability and regulated apoptosis while positively influencing the composition of the intestinal microbiota. The findings align with previous research showing that Ef treatment exerts multiple beneficial effects on IPEC-J2 cell integrity, cell permeability, and intestinal microbiota (Pézsa et al., 2022; Xu et al., 2022).
Enterocytes not only function as selectively permeable filters for nutrient absorption but also serve as the first line of defense against foreign antigens (Gu et al., 2014). The viability of IPEC-J2 cells was positively correlated with EfEV at concentrations of 0.01–1 μg EV protein/mL; however, higher concentrations of EVs inhibited cell viability. A study by Pézsa et al. (Pézsa et al., 2022) found that inoculation of Ef National Collection of Industrial, Food and Marine Bacteria (NCIMB) 10415 at a concentration of 108 CFU/mL significantly reduced the viability of IPEC-J2 cells, whereas low concentrations had no impact on cell viability. This finding is consistent with results for Ef NCIMB 10,415 (Pézsa et al., 2022). Consequently, we selected 1 μg EV protein/mL as the optimal concentration for the subsequent studies.
Autophagy, apoptosis, and permeability are key processes that influence cellular health and function (He et al., 2019; Pederzolli et al., 2018; Tian et al., 2020). In this study, we observed that EfEV supplementation increased the relative expression levels of TJP1, TJP2, CCND1, P53, IL1A, CASP8, BECN1 and ATG16L1 in IPEC-J2 cells. Previous studies have shown that Ef supplementation can protect the intestinal mucosa and tight junctions, thereby reducing intestinal damage (He et al., 2021). Cell-free extracts of Ef L-15 enhanced proliferation and self-renewal capacity in mouse skin-derived precursor cells (Kim et al., 2020). These findings suggest that probiotic EVs can influence cellular functions in ways similar to their mother cells, with enhanced autophagy potentially supporting the survival and spatial adaption of newly proliferating cells.
SOD, GSH-Px, and T-AOC are commonly used indicators of antioxidant activity (Wang et al., 2019). In this study, EfEV supplementation increased GSH-PX and reduced LDH activity in IPEC-J2 cells. Yu et al. (Yu et al., 2019) found that supplementing broiler chickens with Ef NCIMB 10,415 improved meat quality and enhanced muscle antioxidant capacity, including increased activity of CAT, SOD, T-AOC, and GSH-Px in muscle tissue. Probiotic EVs and probiotics may enhance their antioxidant capacity and help maintain intestinal function by regulating the expression of intestinal antioxidant enzymes.
Based on the observed positive effect of EfEV on IPEC-J2 cells, we can conclude that the role of EfEV in the intestine further investigation. To explore this, we examined their impact on the fecal microbiota of weaned piglets using in vitro fecal culture. Weaning often disrupts intestinal microbiota composition, characterized by a decline in Lactobacillus and an increase in Escherichia coli, Shigella, and Acetobacteria (Frese et al., 2015; Su et al., 2008). Previous studies have shown that Ef can regulate intestinal microbiota composition and stimulate the mucosal immune response, thereby enhancing resistance to enteric pathogens such as Salmonella, Escherichia coli, and Campylobacter spp. (V et al., 2017). Consistent with these findings, this study demonstrated that EfEV reduced the abundance of Escherichia coli-Shigella species, indicating that EfEV exhibits similar antimicrobial properties to Ef in inhibiting pathogenic microorganisms. In addition, our analysis highlighted changes in beneficial gut bacterial populations.
SCFAs, as products of intestinal microbiota, are playing an increasingly important role (Chang et al., 2022). The gut microbiota ferments to produce SCFAs, mainly acetic acid, propionic acid, and butyric acid, which are quickly absorbed by the host’s gut lining to supply energy and enhance the host’s metabolic capacity (Zhao et al., 2023). Lachnoclostridium of the Lachnospiraceae family may enhance the absorption of volatile fatty acids, thereby increasing the energy utilization rate of the recipients, increasing the diameter of muscle fibers, and reducing drip loss (Lei et al., 2022). Megasphaera, belonging to the Firmicutes phylum, can convert lactic acid to butyric acid (Rey et al., 2014; Wang et al., 2022). Consistent with these microbial changes, our metabolite analysis detected an increase in acetic acid and butyric acid levels, aligning with the observed enrichment of the [Eubacterium]_aligens_group, Lachnoclostridium, Butyricicoccus, and Megasphaera. These findings underscore the potential of EfEV to modulate microbiota composition and metabolic outputs, contributing to improved intestinal health in weaned piglets.
Conclusions
In conclusion, we reveal for the first time that EfEV is beneficial to the intestinal environment. It reduces the abundance of SCFA-producing bacteria, thereby affecting SCFA production. The increase in SCFAs may also influence cell viability and have a positive impact on the intestinal barrier. Our study reveals another possible mechanism by which Ef affects the intestinal environment.
Animal Welfare Statement
All animal care and treatment procedures were approved by the Animal Care and Use Committee of Foshan University, which meets the ethical standards in Laboratory animal—Guidelines for ethical review of animal welfare (The National Standard of the People’s Republic of China GB/T 35892–2018).
Authors’ Contributions
Q.Q. contributed to the study conceptualization, data curation, formal analysis, validation, visualization. Methodology, investigation, and writing (original draft) were performed by M.L., J.S., and S.L. Funding acquisition were performed by Q.Q. Writing (review and editing) were performed by X.F., H.Z., and Q.Q. All authors have read and agreed to the published version of the article.
Footnotes
Funding Information
This research was funded by the National Natural Science Foundation of China (Grant No. 31902228), Special Fund for Science and Technology Innovation Cultivation of Guangdong University Students (Grant No. pdjh2024b397).
Disclosure Statement
They have no conflict of interest.