
Abstract
Methicillin-resistant Staphylococcus aureus (MRSA) poses a major threat to human health and food safety, especially when bacteria form biofilms or invade host cells, which may cause recurring infections. A new solution is therefore urgently needed. The antimicrobial peptide innate defense regulator (IDR)-1018 and its derived peptide 1018M showed promising antimicrobial and antibiofilm activities. Nevertheless, their antibacterial efficacy against intracellular MRSA and protease tolerance remains to be promoted. Therefore, we synthesized D-amino acid substitution peptides D1018 and D1018M. The antimicrobial activity against MRSA of these novel peptides was increased by 1-fold (D1018) or remained constant (D1018M) compared with L-amino acids peptides. Their bactericidal mechanisms involve cell wall destruction, membrane penetration, and genomic DNA disruption. As expected, the stability of D1018 and D1018M was increased by 2–32 times against pepsin, trypsin, and cathepsin K. In addition, by D-amino acids substitution, the antibiofilm ability of D1018 was increased by 1.6 times, and the anti-intracellular bacterial activity of D1018M was improved 3.2–5.7 orders of magnitude. These data indicated that D1018M is a potential antimicrobial candidate for recurring MRSA infections.
Introduction
Staphylococcus aureus (S. aureus) is a prominent pathogen that threatens human health and food safety. Under selective pressure of antibiotics, S. aureus was provoked to evolve its own strategies to survive (Hong et al., 2016). Methicillin-resistant Staphylococcus aureus (MRSA), renowned for its resistance to the majority of β-lactams due to a mutant penicillin-binding protein (PBP-2a), could cause serious hospital- and community-, biofilm-associated and intracellular infections (Llarrull et al., 2009). Biofilms are one of the basic survival tactics associated with planktonic microorganisms in adverse environments (de la Fuente-Núñez et al., 2014). Resistant to host defense mechanisms and antibiotics, these multicellular communities pose a major risk in refractory infections. In addition, intracellular infection triggered by MRSA has been widespread concern. Once internalized, MRSA has the ability to keep inactive but persist, potentially giving rise to recurrent infections (Monack et al., 2004). Microorganisms have exhibited various resistance patterns, rendering many existing treatments ineffective at therapeutic doses (Buccini et al., 2020). Vancomycin was the critical and decisive drug for the treatment of MRSA infections among conventional antimicrobials (Levine, 2006). However, considering the complex issues caused by biofilm and intracellular bacteria, development of novel antimicrobial agents remains a major challenge (Nelson, 2003).
Antimicrobial peptides (AMPs) constitute a class of small peptides that are widely distributed across all forms of creatures (Kumar et al., 2018; Fjell et al., 2011). Carrying cationic and hydrophobic residues, AMPs display a wide spectrum of antimicrobial activity against both Gram-positive and Gram-negative bacteria and have become promising drug candidates. With multitarget antimicrobial mechanisms, such as the destruction of cell walls and membranes, and binding to DNA, etc. (Kościuczuk et al., 2012; Li et al., 2021), it is difficult for AMPs to form drug resistance (Qiu et al., 2017; Lin et al., 2023). One notable example is innate defense regulator (IDR)-1018, a derivative synthesized from natural bovine host defense peptide bactenecin, which can hinder the accumulation and expedite the degradation of stringent response signaling molecule ppGpp (Wang et al., 2018a); Wieczorek et al., 2010). ppGpp can be triggered by a range of stressful conditions including biofilm development and intracellular infection, when nutrient starvation happens, thus leading to tolerance to antibiotic exposure (de la Fuente-Núñez et al., 2014). Utilizing amino acid substitution and molecular docking, 1018M, a novel peptide was designed, which was more potent than IDR-1018 at antimicrobial and ppGpp binding activity (Jiale et al., 2021). However, their antibacterial, antibiofilm, and anti-intracellular infection effects remain to be promoted (Jiale et al., 2021; Wang et al., 2018a).
Protease is one of the important factors affecting efficacy of AMPs. Various environments, such as cells, tissues, and body fluids, are rich in proteases. In addition, proteases secreted by bacteria in biofilm and produced by phagocytes in lysosomes seriously threaten the activity of IDR-1018 and 1018M, when acting on biofilm and intracellular bacteria (Böttger et al., 2017; Tan et al., 2021; Uhlig et al., 2014). Therefore, there is a need to explore novel and viable strategies aimed at enhancing the efficiency and conquering the instability of these peptides. It is widely known that one of the strict interaction specificities between enzyme and substrate is called stereoisomerism specificity, where the enzyme shows selective activity toward a specific chirality of its substrates. Compared with the transformation of chemical bonds, substituting D-enantiomers for L-amino acids with the goal of furthering the protease stability of AMPs appears to be a more straightforward approach (Qiu et al., 2017). Moreover, previous researches have demonstrated that D-amino acid substitution could not only protect peptides from proteolysis but also improve or maintain their bioactivities (Khara et al., 2016; Li et al., 2016). Therefore, we hypothesized that D-amino acid substitution confers IDR-1018 and its derivatives 1018M enhanced proteolytic stability and anti-biofilm/intracellular bacterial activities even in the action environment full of enzymes.
In the present study, D-enantiomers of IDR-1018 and 1018M (D1018 and D1018M) were synthesized. Subsequently, their stability, bioactivity (antibacterial, antibiofilm, and anti-intracellular bacterial activity), and related mechanisms were explored.
Materials and Methods
Bacterial strains and cell lines
The bacterial strains used in this study included MRSA ATCC 43300, MRSA ATCC 6538 (procured from the American Type Culture Collection), and clinical strain MRSA 49303 (isolated from sputum samples at Ningbo Medical Center Lihuili Hospital). The RAW 264.7 cells were provided by Dr. Guo Hua (Ningbo University).
The preparation of peptides
Every residue in IDR-1018 and 1018M was substituted by D-amino acids (Table 1) (Jiale et al., 2021; Wu et al., 2022). The methodology of solid-phase synthesis was utilized to synthesize all the mentioned peptides: the peptides were synthesized from the C-terminus to the N-terminus through resin loading, stepwise coupling, and deprotection. After cleavage from the resin, the crude product was precipitated with diethyl ether, purified by high-performance liquid chromatography, and freeze-dried.
The Minimum Inhibitory Concentrations of IDR-1018, 1018M, D1018, and D1018M against MSSA ATCC 6538, MRSA ATCC 43300, and MRSA 49303

aData were previously reported (Jiale et al., 2021).
bLowercase letters refer to D-amino acids.
MIC, minimum inhibitory concentration; MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-sensitive Staphylococcus aureus.
Antimicrobial activity
Minimum inhibitory concentration
The minimum inhibitory concentration (MIC) values of peptides were determined using the microtiter broth dilution method. Bacterial suspensions (90 μL, 1 × 105 CFU/mL) were mixed with 10 μL of peptides (2.5–640 μg/mL), vancomycin, or sterile water and then incubated at 37°C for 16–24 h. The MIC was defined as the lowest concentration without visible bacterial growth (Wiegand et al., 2008).
Time-kill curves
Exponential-phase bacterial cultures (1 × 105 CFU/mL) were treated with peptides at 1×, 2×, and 4× MIC. Bacterial growth was monitored at different times by sampling, serial dilution, and colony counting on agar plates (Yang et al., 2019). Sterile water and vancomycin (2× MIC) were included.
Proteolytic stability
The enzymolysis stability of the peptides was conducted by incubating them at 37°C for 4 h with pepsin (pH 2.0, 3000 U/mg), trypsin (pH 8.0, 250 U/mg), or cathepsin K (10:1 w/w; peptide and protease concentrations were 320 μg/mL and 32 μg/mL, respectively). MIC of the treated peptides was determined (Yang et al., 2017).
Resistance
The resistance of MRSA ATCC 43300 against peptides was evaluated using MIC assays. Bacteria were mixed with peptides at various concentrations. After incubation, bacteria from the 1/4 MIC well were transferred to fresh culture. The MICs were determined again over the following 30 days.
Effects of peptides on cell wall and membrane
Scanning/transmission electron microscope observations
Log-phase MRSA ATCC 43300 (108 CFU/mL) was treated with or without peptides (4× MIC) at 37°C for 2 h. Bacteria were then fixed overnight. For scanning electron microscopy (SEM), bacteria were dehydrated, dried, coated, and imaged using SEM S4800. For transmission electron microscopy (TEM), bacteria were prefixed, dehydrated, embedded, polymerized, stained, and observed under TEM Hitachi H-7650(Yang et al., 2019).
Membrane permeabilization analysis
The propidium iodide (PI) uptake assay was determined to investigate the impact of peptides on bacterial membrane integrity. Mid-logarithmic phase cultures were treated with 1×, 2×, and 4× MIC of peptides at 37°C for 0.5 and 2 h. After coincubation with 50 μg/mL PI for 15 min, the samples were analyzed using a flow cytometer (FACS Calibur) (Yang et al., 2017).
Effects of peptides on bacterial genomic DNA
Gel retardation assay
The interaction of peptides with MRSA genomic DNA was performed using gel retardation assay. Genomic DNA was mixed with D1018 and D1018M (1.25–40 μg/mL) for 10 min, and the samples were analyzed by electrophoresis.
Circular dichroism spectroscopy
To investigate the impact of peptides on the secondary structure of DNA, peptides (40 μg/mL) were mixed with genomic DNA (150 μg/mL) for 10 min. The samples were then loaded in cuvettes (1.0-mm path length), and J-1700 CD spectrometer was employed to record spectra from 230 to 320 nm with a 10 nm/min scanning speed (Wang et al., 2016).
Antibiofilm activity of peptides
Effects on biofilm formation
The effect of peptides on biofilm formation was evaluated by coincubation bacteria (1 × 108 CFU/mL) with peptides at 37°C for 24 h. After removing nonadherent bacteria, the biofilms were fixed for 15 min, stained with 0.1% crystal violet, and dried overnight. The bound dye was dissolved, and absorbance (570 nm) was measured (de Breij A et al., 2016; de Breij A et al., 2018).
Biofilms observed by SEM
To further explore the biofilm inhibition activity of the peptides, 24-well plates containing sterile silicon slides were loaded with bacteria (1 × 108 CFU/mL) and peptides (64 μg/mL). After 24 h of incubation, planktonic bacteria were gently washed off with phosphate buffer saline (PBS). The biofilms on the slides were then observed by S4800 SEM (Yang et al., 2019).
Anti-intracellular bacteria activity
Effects on intracellular forms of MRSA
RAW 264.7 macrophages (2.5 × 105 cells/mL) were seeded into 24-well plates (375 μL/well) and cultured in antibiotic-free Dulbecco's Modified Eagle Medium (DMEM) for 24 h. Cells were infected with log-phase MRSA (375 μL, 2.5 × 107 CFU/mL) for 30 min. Extracellular bacteria were removed using 100 μg/mL gentamicin, and the cells were rinsed twice with PBS. Peptides at various concentrations were added to the cultures for 24 h. After washing, cells were treated with 0.1% Triton-X, and intracellular bacteria at 0 and 24 h were quantified (Wang et al., 2018b).
Quantification of peptides uptake
To quantify peptide cellular uptake, RAW 264.7 macrophages (2.5 × 105 cells/mL, 375 μL/well) were cultured overnight and incubated with fluorescein isothiocyanate (FITC)-labeled peptides at concentrations of 15.625, 31.25, and 62.5 μg/mL for 24 h. The cells were then washed with PBS and treated with 0.04% trypan blue for 15 min to minimize the impact of peptides bound with membranes. Uptake was measured using flow cytometer (FACS Calibur) (Kamaruzzaman et al., 2016).
Mechanism of peptides cellular uptake
To study the mechanism of peptide uptake in RAW 264.7 macrophages, cells were pretreated with endocytosis inhibitors (3 mM amiloride, 5 mM methyl-β-cyclodextrin (MβCD), 20 μM nocodazole, or 6 μg/mL chlorpromazine) for 1 h at 37°C. Then, 62.5 μg/mL of FITC-labeled peptides were added and incubated for 6 h. The cells were then treated with 0.04% trypan blue, and fluorescence intensity was measured using flow cytometer (Gomarasca et al., 2017).
Statistical analysis
All assays were performed in triplicate. Data were analyzed using GraphPad Prism 6 and are presented as mean ± standard deviation (standard error of the mean). Comparisons among multiple groups were confirmed using one-way ANOVA or Student’s t test, with significance set at P < 0.05.
Results
Antimicrobial activity
MIC
As described in Table 1, D1018 and D1018M had MICs of 2 μg/mL against MRSA ATCC 43300, showing stronger antibacterial activity than IDR-1018 and matching 1018M. For other pathogens, the derived peptides retained similar activity to the parent peptide, likely due to strain differences or drug storage conditions.
Time-kill curves
The time-kill kinetics of D1018 and D1018M are depicted in Figure 1A. Both peptides significantly reduced viable bacteria within 0.5 h at 1×, 2×, and 4× MIC (4.35 log CFU reduction). Their antimicrobial effect was stronger than vancomycin (2× MIC) within 4 h. After 4 h, bacterial growth slightly increased with 1× MIC D1018M, while rejuvenation occurred with 2×MIC vancomycin after 10 h. In contrast, 2× and 4× MIC peptides maintained their effect for 24 h. These results indicate that the novel peptides have longer-lasting effects than vancomycin and mimic their parental peptides (Jiale et al., 2021).
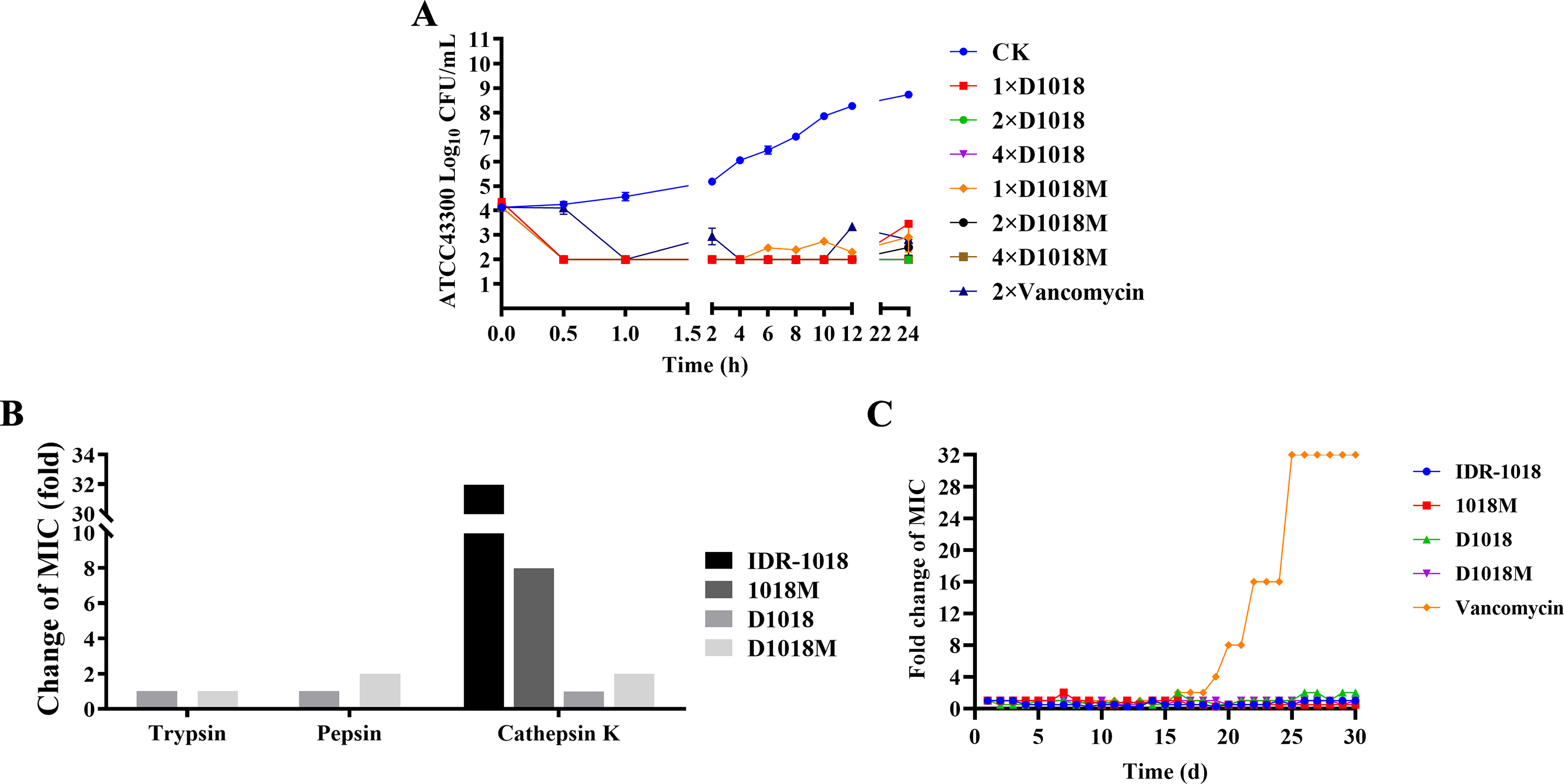
The time-kill kinetics, proteolytic stability, and resistance of peptides.
Proteolytic stability
As shown in Figure 1B, after exposed to pepsin, trypsin, and cathepsin K, the antibacterial activity remained unchanged (D1018, MIC = 2 μg/mL) or doubled (D1018M, MIC = 4 μg/mL). D1018 and D1018M showed 2–32 times increased proteolytic stability compared with parental peptides (MIC increased 16, 16, and 32 times after treatment with pepsin, trypsin and cathepsin K for IDR-1018, and MIC increased 1, 32, and 8 times after treatment with pepsin trypsin and cathepsin K for 1018M) (Jiale et al., 2021).
Resistance
After 30 serial tests, the MIC values of peptides against MRSA did not change (1–2 times of original MIC), which illustrated that no resistant mutant was generated (Fig. 1C). In contrast, vancomycin induced a 32-fold MIC increase. These results underscore the potency of peptide series as promising novel antibacterial candidate.
Effects of peptides on cell wall and membrane
SEM/TEM observations
SEM analysis revealed the bacterial morphology, integrity, and cellular structure. In the control group, MRSA ATCC 43300 showed intact cell surfaces and normal morphology (Fig. 2A, B). However, after 2 h of exposure to 4× MIC D1018 or D1018M, about 70% of bacteria exhibited shrinkage, bubbling, and severe holes. In addition, TEM could further confirm these effects, presenting normal ultrastructure and homogeneous cytoplasm of microbes in the control group (Fig. 2B). However, D1018 caused cell wall and membrane destruction, while D1018M led to grossly damaged bacteria, with over 50% appearing as ghost cells and visible cytoplasmic leakage.
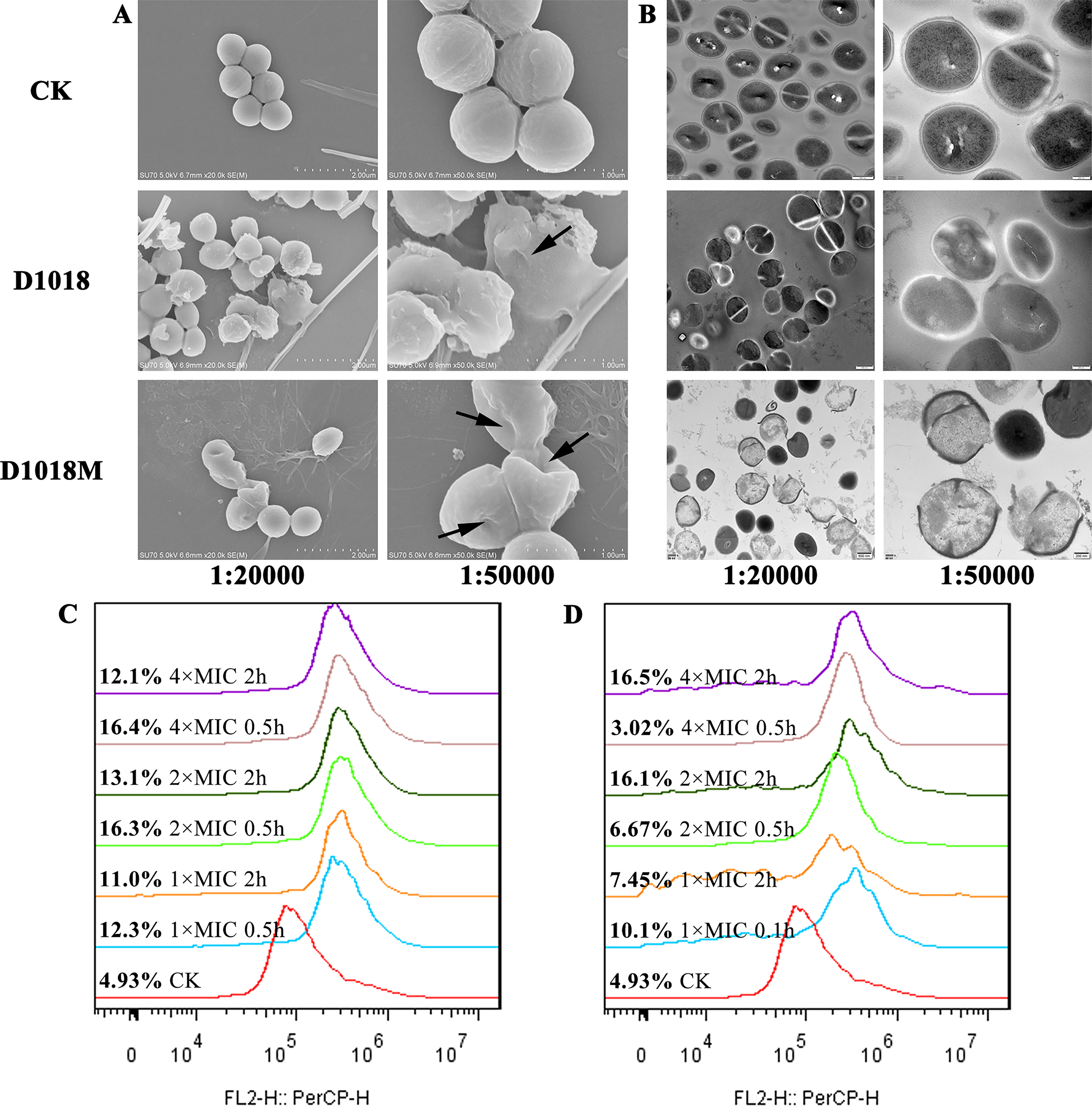
Effects of D1018 and D1018M on cell wall and membrane.
Membrane permeabilization analysis
Bacterial membrane integrity was assessed via flow cytometry with PI staining (Fig. 2C, D). Normal cells showed 4.93% fluorescence. After 0.5 h and 2 h coincubation with 1×, 2×, and 4× MIC of D1018 or D1018M, PI-permeable bacteria ranged from 11.0% to 16.4% and 3.02% to 16.5%, respectively, indicating these peptides possess a slight ability to penetrate bacterial membranes.
Effects of peptides on bacterial genomic DNA
Gel retardation assay
DNA gel retardation assay (Fig. 3A) evaluated the DNA-binding ability of D1018 and D1018M. The result showed that D1018 completely inhibited DNA migration at 5 μg/mL, while D1018M required 10 μg/mL, indicating both peptides bind bacterial DNA, with D1018 showing superior efficiency.

Interaction of D1018 and D1018M with MRSA ATCC 43300 bacterial genomic DNA.
CD spectroscopy
The destructive effects of peptides on bacterial genomic DNA structure were analyzed via CD spectroscopy (Fig. 3B). Normal DNA showed a negative peak at 250 nm and a positive peak at 270 nm. Treatment with D1018 and D1018M caused a sharp decrease in the negative peak, while D1018 nearly abolished the positive peak. These results confirm greater impact of D1018 on DNA conformation, aligning with findings from gel retardation.
Antibiofilm activity of peptides
Effects on biofilm formation
Crystal violet staining assay showed that D1018 demonstrated dose-dependent inhibition, with biofilm reduction of 26.62%, 55.27%, and 70.76% at 16, 32, and 64 μg/mL, respectively. D1018M also inhibited biofilm formation, reducing it by 50.89% at the highest concentration (Fig. 4A).
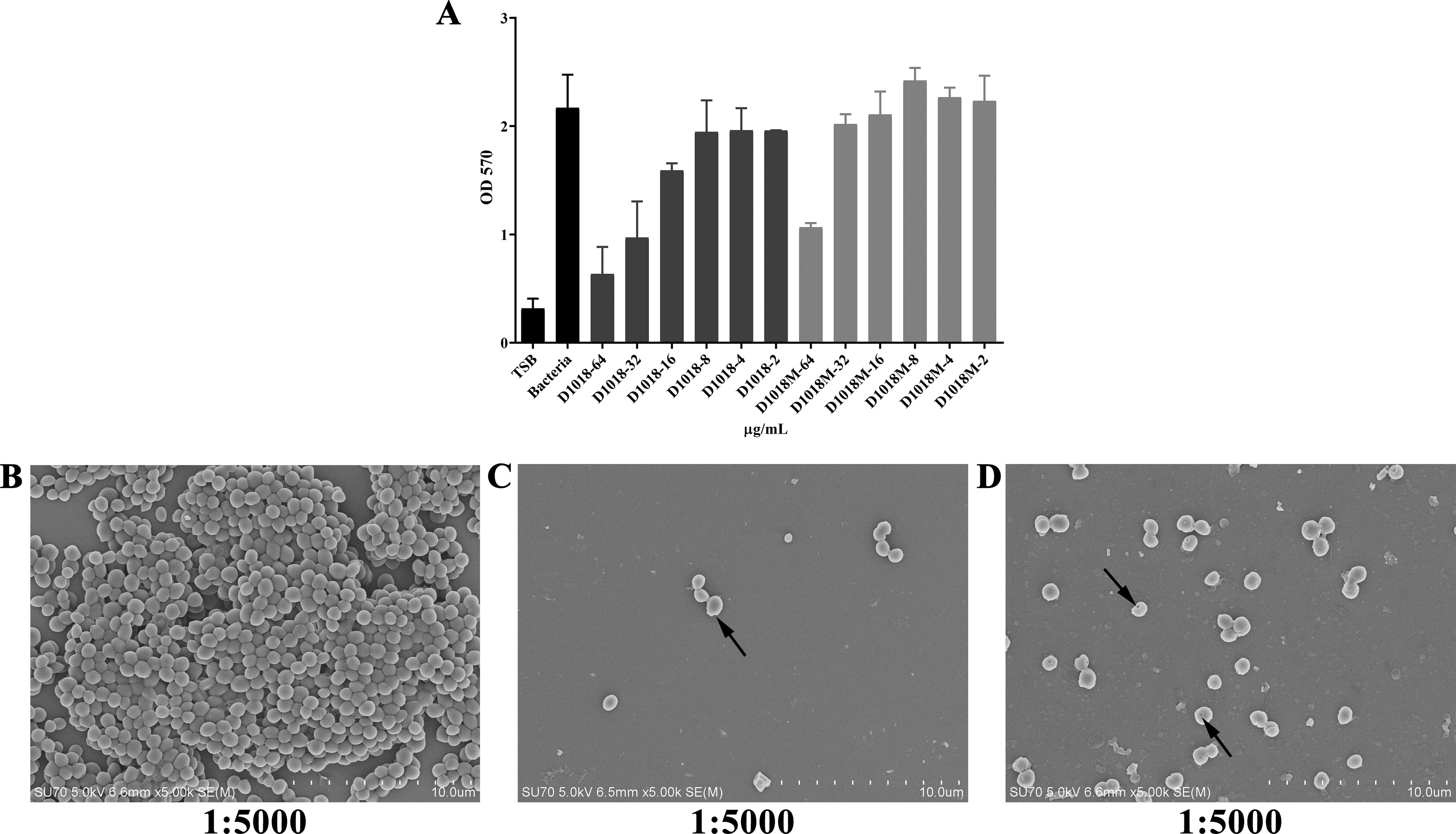
The inhibition abilities of D1018 and D1018M against MRSA biofilm formation.
Biofilms observed by SEM
The antibiofilm efficacy of peptides was further proved by SEM observation (Fig. 4B–D). Untreated bacteria formed dense biofilms with normal morphology mostly. D1018 nearly eliminated bacterial adhesion, leaving few intact cells, while D1018M partially inhibited biofilm formation, causing severe bacterial damage. These findings aligned with crystal violet staining results.
Anti-intracellular bacteria activity
Effects on intracellular forms of MRSA
The anti-intracellular effects of peptides were displayed in Figure 5A. IDR-1018 showed minimal impact, reducing bacteria by only 0.76–0.91 log CFU/mL. However, 1018M reduced bacteria by 3.89, 1.78, and 1.87 log CFU/mL at 250, 125, and 62.5 μg/mL, respectively. D1018 exhibited similar activity, with reductions of 3.35, 1.71, and 0.75 log CFU/mL at the same concentrations. Notably, D1018M was significantly more effective, reducing bacteria by 3.28 log CFU/mL at just 15.6 μg/mL and completely eradicating intracellular MRSA at 250 μg/mL. This dose-dependent efficacy highlights potent bacteriostatic effects of D1018M.

Intracellular antimicrobial activity and mechanism of IDR-1018 and its derivatives against MRSA in RAW 264.7 cells.
Quantification of peptides uptake
The proportion of cells penetrated by IDR-1018, 1018M, and D1018 increased with the increment of peptide concentration (Fig. 5B). FITC-D1018 exhibited the highest uptake, with 64.2%, 74.3%, and 90.9% of cells labeled at 15.625, 31.25, and 62.5 μg/mL, respectively. FITC-D1018M showed slightly lower uptake, with 64.5%, 58.7%, and 63.2% labeled cells. These results suggested all peptides can enter RAW 264.7 macrophages.
Mechanism of peptides cellular uptake
All endocytosis inhibitors reduced the uptake of FITC-IDR-1018, FITC-1018M, and FITC-D1018. MβCD caused the strongest suppression (7.3%−20.9%). Other inhibitors did barely impact the uptake of IDR-1018 and 1018M. For D1018, uptake decreased by 7% with chlorpromazine, while D1018M uptake was inhibited by nocodazole (7.1%) and chlorpromazine (7.6%) (Fig. 5C).
Discussion
MRSA, a notorious pathogen tactically resistant to multiple antibiotics, causes many infections incredibly difficult to treat. Moreover, intracellular lifestyle and biofilm formation enable evasion of the host immune response and antibiotic interaction, withstand hostile environments, and lead to chronic and persistent inflammation (Monack et al., 2004). The gravity of this looming threat to public health necessitates the innovation of new drugs that can meet the urgent requirements of clinical medicine. As a vital component of the organism’s innate defense system, AMPs drew worldwide attention and offer a pathway to conquer drug resistance (Zhong et al., 2020). However, their appealing therapeutic potential has largely been impeded by shortcomings, such as protease susceptibility, improvable antimicrobial activity, and cytotoxicity. Several strategies have been employed to modify the peptides to circumvent the above issues in order to improve the stability and reduce the toxicity, including the incorporation of unnatural amino acids (Ong et al., 2014). In this study, we synthesized D-enantiomers of IDR-1018 (D1018) and its novel derivative with potent antimicrobial/biofilm activities 1018M (D1018M), and then investigated the effects on the bioactivity, stability, and mechanism of action.
It is well known that unnatural amino acids rarely act as endogenous protease substrates. Therefore, D-amino acid substitution was very attractive for the design of novel proteolytically stable AMPs. Although, the impact of substitution on their antimicrobial activity is still controversial (Wątły et al., 2021). Fortunately, the substitution of D-amino acids in IDR-1018 and 1018M did not affect or even improve their antibacterial capacity (Table 1 and Fig. 1A). In terms of proteolytic stability, pepsin, trypsin, and the plasma protease tolerance of AMPs have been the hot spot of the current researches for their oral or injection use. However, when it comes to intracellular antibacterial activity, cathepsin resistance is even more important, for the reason that phagolysosome may be the main site of action for AMPs. Hence, the cathepsin tolerance and intracellular anti-MRSA activity of D1018 and D1018M, the unnatural amino acid substitution peptides, were first investigated. As expected, D1018 and D1018M exhibited excellent proteolytic stability, especially against cathepsin, improving their clinical application potential against planktonic or intracellular bacteria (Fig. 1B). In addition, no bacteria-resistant mutants to all the peptides were produced after 30 serial passages (Fig. 3C), indicating the safety of the peptides to reduce drug-resistant bacteria infection incidence.
In addition, the incorporation of D-amino acids into peptides didn’t change the bactericidal mechanism of IDR-1018 and 1018M. D1018 and D1018M were also able to disrupt the cell wall of bacteria, penetrate the cell membrane, and subsequently bind to genomic DNA afterward (Jiale et al., 2021). These multitarget antimicrobial mechanisms may be an explanation for the difficulty of drug resistance formation. Moreover, D1018 and D1018M showed more serious damage to bacterial cell walls and DNA, when compared with their parental peptides. Most of the bacterial cells were cleaved by D1018M (Fig. 2B), while the secondary structure of genomic DNA was severely destroyed by D1018 (Fig. 3B). All the differences between L- and D- peptides may attribute to the change of secondary structure or dynamics of self-assembled of these AMPs after substitution (Kapil and Sharma, 2021; Ye et al., 2018).
Bacterial biofilms contribute to the development of chronic infections due to their heightened tolerance to antibiotics and the body’s defense system (Høiby et al., 2010). Previous studies have shown that IDR-1018 and 1018M have biofilm formation inhibition ability against S. aureus. In the present study, we tested the antibiofilm activities of the derivative peptides and found that D1018 possessed more potent effects against MRSA ATCC 43300 biofilms than the parent peptides. The results were consistent with previous researches (Ben Hur et al., 2022; Casciaro et al., 2019; Guo et al., 2021). The possible explanations of this thesis include the following aspects: 1) With better antiprotease activity, the interaction time of peptides and bacteria, even the targets such as DNA, ppGpp, etc., was prolonged (Casciaro et al., 2019). 2) D-amino acid peptides may have a relatively greater affinity for biofilm extracellular polysaccharides (Yin et al., 2013). However, the biofilm inhibition ability of D1018M was significantly lower than that of 1018M. This may result from the D-isomer substitutions at R4, W5, W6, R7 and R9, which could affect the peptide’s interaction with the signaling nucleotide ppGpp, thereby reducing its antibiofilm capacity.
The intracellular anti-MRSA properties of IDR-1018 and its derivatives were first evaluated in this study. It was not surprising to see that 1018M, D1018, and D1018M displayed improved intracellular bacterial activity (Fig. 5A) due to their excellent proteolytic stability, especially D1018M. In Addition, another key to excellent intracellular antibacterial performance hinges on the intracellular accumulation of these peptides. Hence, the permeability of these peptides was explored. We found that IDR-1018, 1018M, and D1018 could enter into RAW 264.7 macrophages in a dose-dependent manner, whereas D1018M showed a lower penetration efficiency (Fig. 5B). All the data demonstrated that D-amino acid substitution could not contribute to improving the peptides’ cell internalization ability. In order to further explore the internalization mechanisms of these peptides, different endocytosis inhibitors were used for blocking specific endocytosis pathways (Lundin et al., 2008). The uptake of all the peptides was blocked by MβCD, which indicated that lipid rafts were the main pathways for the internalization of this series peptides. Moreover, the uptake of D1018 was slightly dependent on clathrin-mediated endocytosis. D1018M entered into cells partly via clathrin-mediated endocytosis and microtubule polymerization (Fig. 5C) (Lundin et al., 2008; Suzuki et al., 2016; Wadia et al., 2004). The cell internalization pathways of IDR-1018 and its derived peptides were different from our previously studied fungal defense peptides, which enter cells via clathrin-mediated endocytosis and micropinocytosis. Overall, we speculated that the internalization efficiency and mechanism of AMPs were basically not influenced by D-amino acid substitution but by the other characteristics of the AMPs.
In summary, D-amino acid substitution could significantly enhance the proteolytic stability of IDR-1018 and D1018M as well as maintain or even improve their bioactivities. The derivatives D1018 and D1018M were proved to be more potent than the parent peptides in damaging cell wall and interacting with the genomic DNA of microbes in our study. In addition, it was found that D1018M could kill the internalized bacteria, suggesting that D-amino acid modification might be a tactic to extend the range of action of AMPs. One implication of this study is the possibility that D-amino acid substitution might facilitate and hasten the transition of AMPs from bench to bedside. Furthermore, a comprehensive exploration of the antibacterial mechanisms and in vivo effects is warranted.
Authors’ Contributions
X.W. and J.J. conceived and designed experiments. Z.Z., Z.J., X.D., R.W., Q.W., H.W., and X.W. carried out all the experiments. Z.Z. and X.W. contributed in writing. X.W., Z.J., and T.L. contributed in funding acquisition.
Footnotes
Funding Information
This work was supported by Natural Science Foundation of Ningbo (2024J407), One Health Interdisciplinary Research Project of Ningbo University (HY202402), 2021 Ningbo Science and Technology Plan Project (Grant No. 2021S119), 2020 Yinzhou District Science and Technology Plan Project in Ningbo City, and the National Natural Science Foundation of China (82002190).
Data Availability
All data analyzed in this study are included in this published article.
Ethics Approval
This article does not contain any studies with human participants or animals performed by the authors.
Consent to Participate
The participant has consented to the publication of the article.
Disclosure Statement
No competing financial interests exist.