
Abstract
Staphylococcus aureus (S. aureus) is a major zoonotic pathogen. To investigate CRISPR carriage in S. aureus isolates from cows with mastitis and the role of the CRISPR system and efflux pumps in antibiotic resistance. We analyzed antibiotic resistance genes and CRISPR loci, sequenced spacers, and assessed correlations between CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) presence and antibiotic resistance in 234 S. aureus isolates. The changes in CRISPR sequences were examined by continuous passage of 360 generations without antibiotic pressure. Subsequently, variations in CRISPR loci and transcript levels were measured under ciprofloxacin (CIP) exposure. In addition, an S. aureus-25-mepA was constructed to evaluate changes in antimicrobial sensitivity and mepA transcript levels in both planktonic and biofilm states. Our results revealed a CRISPR loci detection rate of 7.69% among the 234 S. aureus isolates, with significantly lower rates of the antibiotic resistance genes gyrA, grlA, norA, and tet(M) in CRISPR-positive isolates compared to those in CRISPR-negative isolates (p < 0.05). CIP-resistant strains exhibited loss of repeat and spacer sequence in CRISPR loci, and the transcript abundance of these loci gradually decreased under CIP pressures, indicating that CRISPR loci deletion or transcript level downregulation under antibiotic stress may be a potential regulatory mechanism of antibiotic resistance. Correlation analysis linked CIP resistance in both planktonic and biofilm S. aureus to mepA transcript levels and biofilm integrity. Our study provides insight into the mechanism by which S. aureus develops antibiotic resistance via the CRISPR system and the MepA efflux pump, offering a theoretical foundation for monitoring the prevalence and resistance of pathogenic bacteria.
Introduction
The high frequency of horizontal gene transfer in Staphylococcus aureus (S. aureus)has led to the rapid emergence of multidrug-resistant strains. The CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) system in bacteria acts as an adaptive immune defense against foreign mobile elements, such as plasmids, making it the only known adaptive immune system in prokaryotes (Hur et al., 2014; Peng et al., 2017; Roy et al., 2021). Marraffini and Sontheimer (2008) found that S. epidermidis utilizes CRISPR interference to block plasmid conjugation, thereby preventing plasmid transfer. Dang et al. (2013) found a tight correlation between CRISPR deletion and increased antibiotic resistance. These studies highlight the critical role of the CRISPR system in bacterial defense mechanisms, with its absence linked to heightened antibiotic resistance (Mayorga-Ramos et al., 2023; Roy et al., 2021).
The efflux pump mechanism is a critical means by which bacteria evade the effects of antibiotics (Darby et al., 2023). Fluoroquinolones are vital broad-spectrum antibiotics primarily used to treat severe invasive infections. These antibiotics work by inhibiting type II-DNA gyrase and type IV-topoisomerase, which are involved in bacterial DNA synthesis and replication (Bhatt and Chatterjee, 2022). The MepA efflux pump, part of the multidrug and toxin efflux family, is chromosomally encoded and plays a critical role in fluoroquinolone resistance (Huang et al., 2023; Nishino et al., 2021).
We collected milk samples from mastitis cows in the Ningxia Hui Autonomous Region to investigate the CRISPR carriage of S. aureus and the relationship between the CRISPR system and efflux pump in terms of antibiotic resistance. Our study provides novel insights for developing strategies to mitigate antibiotic resistance in S. aureus.
Materials and Methods
Source of isolates
A total of 234 strains of S. aureus were isolated from milk samples of mastitis cows from 16 farms in Ningxia, China. The whole genome sequencing of 14 methicillin-resistant S. aureus (MRSA) isolates was done at Annoroad Gene Technology, Beijing, China.
Detection of CRISPR loci and antibiotic resistance genes
Genomic DNA was extracted from 234 S. aureus isolates using bacterial genome extraction kits. CRISPR loci and associated antibiotic-resistance genes were identified through polymerase chain reaction (PCR) amplification, and the PCR primer sequences are listed in Supplementary Table S1. The amplified CRISPR fragments were sent to Sangon Biotech (Shanghai, China) for sequencing. The sequencing data were then compared and analyzed using the online tool National Center for Biotechnology Information Basic Local Alignment Search Tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and CRISPR Finder (https://crisprcas.i2bc.paris-saclay.fr/CrisprCasFinder/Index).
Resistance gene information for spacer homologous plasmids
Plasmids with spacer homologous sequences were identified using the CRISPR Target tool (http://crispr.otago.ac.nz/CRISPRTarget/crispr_analysis.html). Information on resistance genes associated with these homologous plasmids can be found at the Center for Genomic Epidemiology (https://genomicepidemiology.org/). CRISPR loci and CRISPR-Cas system types in whole-genome sequenced S. aureus were predicted using the online tools CRISPR Finder (https://crisprcas.i2bc.paris-saclay.fr/CrisprCasFinder/Index) and CRISPR Miner (http://www.microbiome-bigdata.com/CRISPRminer/). RNA secondary structure and minimum free energy (MFE) were predicted using the RNAfold web server (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi).
Molecular docking of MepA protein with ciprofloxacin (CIP)
The tertiary structure of the MepA protein was obtained through homology modeling using AlphaFold (Q2G1X0, identity: 99.7%) and I-TASSER (Iterative Threading ASSEmbly Refinement). The structures of enrofloxacin, norfloxacin, and CIP were downloaded from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). Semi-flexible molecular docking of the receptor proteins was performed using AutoDock 4.0 and AutoDock Vina. The final docking results were evaluated based on scoring and conformational fit and visualized and analyzed using PyMOL and Discovery Studio 2020.
Antibiotic susceptibility and CRISPR loci change in S. aureus under CIP pressure
Bacterial fluids were inoculated in Mueller–Hinton broth (MHB) with 1/4 minimum inhibitory concentration (MIC) CIP, passed to 2–4 generations, and then transferred into MHB with 1/2 MIC CIP and continued to pass 2–4 times until the induced S. aureus could grow in MHB with 4 µg/mL of CIP. The induced resistant S. aureus was passaged in a medium without CIP for five generations and then tested for antibiotic resistance using the standard broth dilution method according to the guidelines of the Clinical and Laboratory Standards Institute. The total mRNA of induced S. aureus was extracted using the Trizol method, and the CRISPR transcript levels were detected by quantitative PCR (qPCR) after reverse transcription. The results were calculated by the 2-ΔΔCT method. The fluorescence quantification primers are shown in Supplementary Table S1, and the reaction system is template DNA 1 µL, forward prime (10 µM) 1 µL, reverse primer 1 µL, 2 × Taq PCR MasterMix 10 µL, and ddH2O 7 µL.
Antibiotic susceptibility and CRISPR loci change in S. aureus after successive passages without antibiotic pressure
The bacterial solution incubated for 18 h was transferred to MHB medium at a ratio of 1:500 and incubated at 37°C and 200 rpm for 6–7 h until the colony count reached 107 colony-forming unit (CFU)/mL. Repeated this step twice, and then repeat the above steps three times a day for 120 days. Finally, 16S rDNA analysis was employed to determine whether the isolates were the same strain. Then the antibiotic susceptibility, CRISPR loci, and CRISPR transcript level changes of S. aureus were determined.
pBT2-mepA recombinant plasmid construction and transformation of S. aureus-25
Primers were designed based on GenBank sequence number AY661734.1, with protective bases added at both ends and SalI and BamHI restriction sites incorporated. The primer sequences are listed in Supplementary Table S1. The amplified mepA DNA fragment was purified and ligated to the pMD19-T (plasmid MD19-T) vector to create the pMD19-T-mepA. The pMD19-T-mepA plasmid was then double-enzymatically digested with SalI and BamHI, and transformed into the S. aureus-25. The pMD19-T-mepA plasmid and the pBT2 plasmid (6.9 kb) were double digested with SalI and BamHI. The mepA fragment was purified, recovered with the digested pBT2 plasmid, and ligated to obtain recombinant pBT2-mepA. The recombinant plasmid, pBT2-mepA, was identified through double digestion with SalI and BamHI and then sent to Sangon Biotech (Shanghai, China) for sequencing.
S. aureus RN4220 competent cells were prepared using the sucrose method. A volume of 6 µL of recombinant plasmid pBT2-mepA (100 ng/µL) was gently mixed with 100 µL RN4220 competence cells. After 25 min of ice bath, the mixture was transferred to a 2 mm rotor and subjected to two electrical shocks (2.9 kV, 25 µF, 200 Ω) with an energization time of 2.5 ms. After 0.9 mL tryptic soy broth was added and cultured at 37°C for 3 h, 100 µL was plated on a tryptone soy agar (TSA) plate containing chloramphenicol (20 µg/mL) and incubated at 37°C for 24 h. The positive colonies were selected and cultured to extract the plasmid, identified by double digestion with SalI and BamHI.
Analysis of S. aureus sensitivity to CIP and mepA transcript abundance in different states
Single colonies of S. aureus were picked and placed in MHB and incubated for 12–14 h(at 37°C, 180 r). A total of 15 mL of bacterial solution was taken, and 5 mL of MHB was added, mixed, and evenly divided into two parts. Equal volumes of CIP and sterile distilled water were added separately and incubated at 37°C with shaking. Samples of 1.5 mL were taken every hour, centrifuged to discard the supernatant, resuspended in phosphate‐buffered saline (PBS), and washed twice. The bacterial solution was diluted to 103 with PBS and 50 µL was spread on a 90 mm Mueller–Hinton Agar (MHA) plate and incubated at 37°C for 18 h. The number of viable bacteria was counted and repeated three times.
The overnight culture of S. aureus was centrifuged and resuspended in MHB, diluted to 106 CFU/mL, and 5 µL was inoculated on a 0.2 µm polycarbonate filter membrane (diameter 25 mm) on a TSA plate and incubated at 37°C for 48 h. The filter membrane and biofilm were transferred to a new plate every 12 h. The biofilm bacteria were transferred to MHA plates containing CIP (25 µg/mL) and cultured for 4 h, with samples taken every hour. The biofilm samples were placed in 9 mL PBS, vortexed at high speed for 5 min to disperse the bacteria, and then the viable bacteria were counted, and repeated three times. The biofilm bacteria cultured for 48 h were collected in 10 mL PBS, vortexed for 5 min, centrifuged, resuspended diluted, and counted for viable bacteria, and repeated three times. Planktonic and biofilm bacteria without CIP were used as controls, and the mepA gene expression before and after CIP treatment was detected by qPCR.
Statistical analysis
The software SPSS 21.0 (IBM Corp., Armonk, NY, USA) was used to process the data, and the chi-square test or Fisher’s exact probability test was performed. p < 0.05 was regarded as a statistically significant difference.
Results
Correlation analysis between antibiotic resistance genes and CRISPR loci in S. aureus
Among the 234 S. aureus isolates, 18 strains carried CRISPR loci, with a positive rate of 7.69% (Table 1). Notably, eight strains (3.42%) simultaneously contained CRISPR 1, CRISPR 2, and CRISPR 3. A total of 43 spacer sequences were identified across all CRISPR loci, of which 28 spacer sequences showed potential homology with plasmids or phages, but only three plasmids with homologous spacer sequences were found to carry resistance genes (Table 2).
Distribution of CRISPR Loci in Staphylococcus aureus Isolates
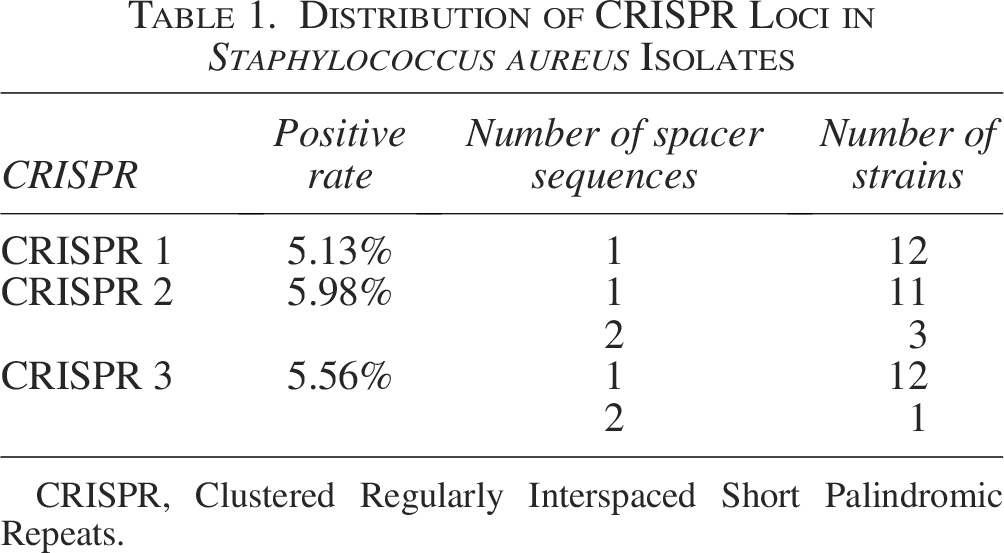
CRISPR, Clustered Regularly Interspaced Short Palindromic Repeats.
Homologous Plasmids of Spacer Sequences Carrying Antibiotic Resistance Gene

A total of 13 resistance genes in seven categories were detected in 234 S. aureus isolates (Table 3). The highest positive rates were gyrA (97.85%) and norA (97.85%), followed by grlA (95.72%). Conversely, genes such as tet(O), optrA, cfr, sul1, sul2, and sul3 were not detected. Among CRISPR-positive S. aureus strains, gyrA (88.89%), grlA (83.33%), and norA (83.33%) exhibited the highest detection rates, which were significantly lower than those of CRISPR-negative isolates (p < 0.05) (Fig. 1A, Table 3). Among CRISPR-positive isolates, penicillin G showed the highest resistance rate, followed by ampicillin (AMP) and amoxicillin/clavulanic acid (Fig. 1B).
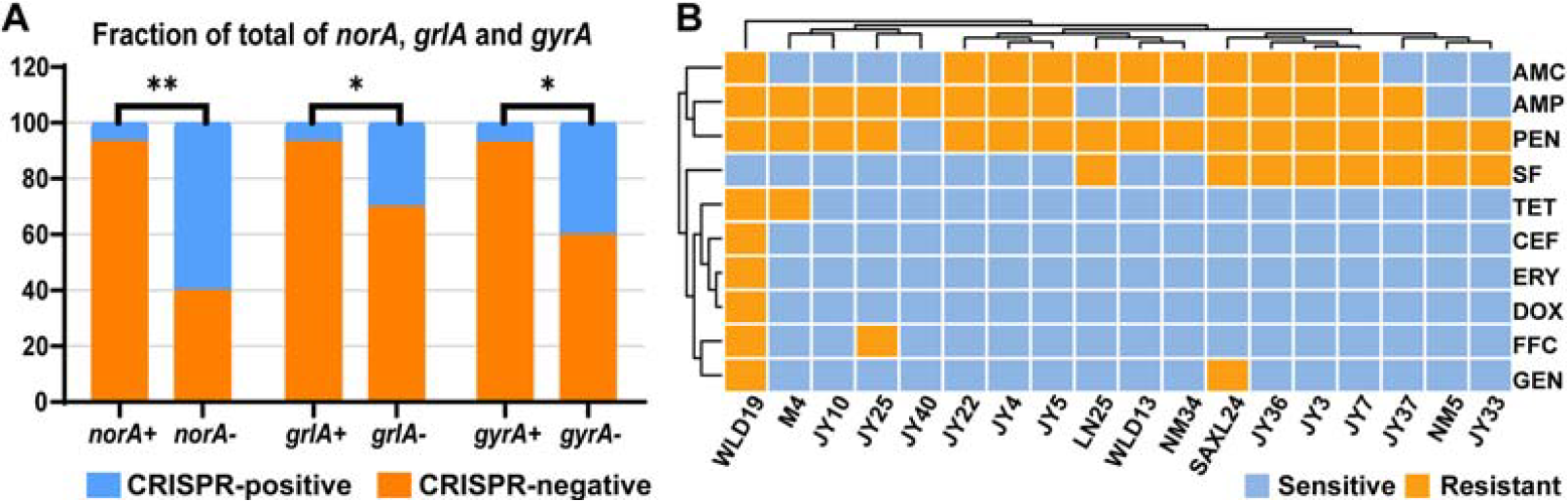
Antibiotic resistance genes and phenotypes spectrum of Staphylococcus aureus (S. aureus).
Association Between Antibiotic Resistance Gene Distribution and CRISPR Loci
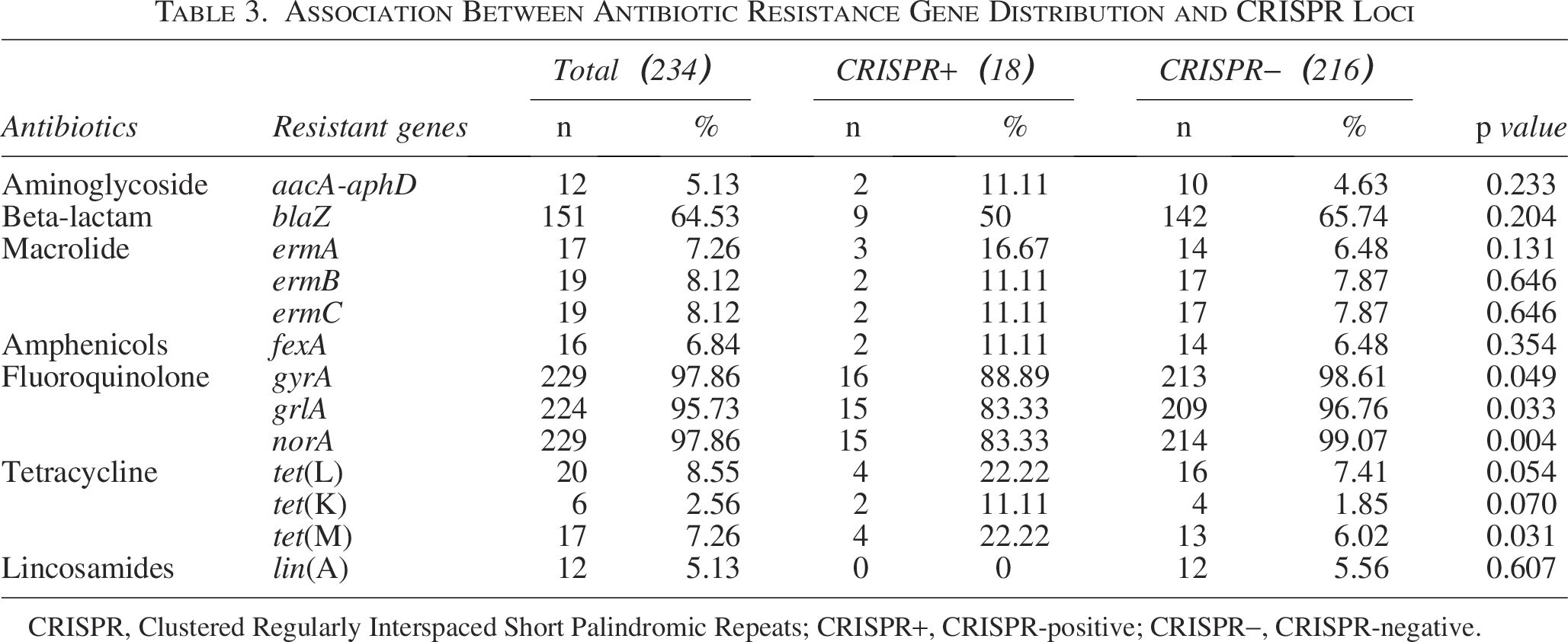
CRISPR, Clustered Regularly Interspaced Short Palindromic Repeats; CRISPR+, CRISPR-positive; CRISPR−, CRISPR-negative.
Analysis of repeat and spacer sequences in MRSA isolates
CRISPR sequences were detected in all 14 MRSA strains. Of which three strains belonged to the CRISPR-Cas-Ι system. The repetitive sequences within the same CRISPR locus exhibited high similarity. After redundancies were removed, 26 distinct consensus direct repeat sequences (CDRs) were identified (Supplementary Table S2), with an average length of 27 bp. All 26 CDRs were able to form secondary structures (Fig. 2). A total of 229 spacer sequences were identified in the 14 MRSA strains, with lengths ranging from 26 to 55 bp. Of these, 74 (32.31%) were homologous to phage sequences, 36 (15.72%) were homologous to plasmid sequences, and 110 (48.03%) were homologous to non-Staphylococcus gene sequences. Some spacer sequences matched multiple foreign gene sequences simultaneously, with 18 spacer sequences matching both plasmid and phage gene sequences.
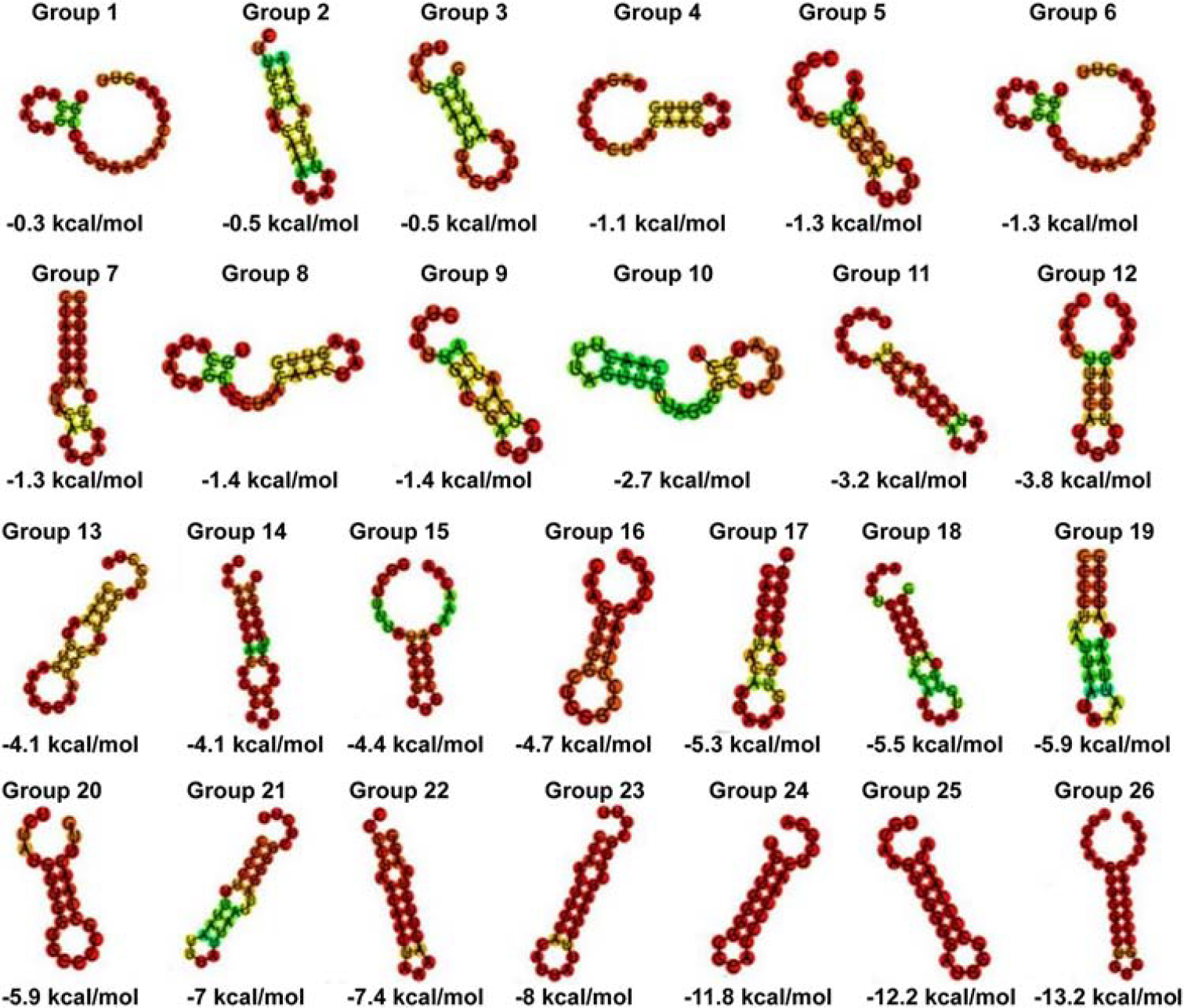
RNA secondary structure and minimum free energy (MFE) of consensus direct repeat sequences (CDRs). Note: Predicted RNA secondary structures are color-coded based on base pairing probability, with red indicating high and green indicating low probability of repetitive sequence structure formation. RNA secondary structure maps and MFE values can predict the degree of sequence stability and conservation of CDRs. Lower MFE values indicate more stable and conserved RNA secondary structures. Increased stem length or higher GC (guanine-cytosine) content contributes to more excellent RNA stability. MFE, minimum free energy.
MepA protein docking with ligand small molecules
The molecular properties of enrofloxacin, norfloxacin, and CIP are summarized in Supplementary Table S3. Following multiple rounds of molecular docking and conformational clustering analysis, the binding free energies of the best conformations for enrofloxacin, norfloxacin, and CIP were found to be −8.92 kcal/mol, −8.81 kcal/mol, and −8.63 kcal/mol, respectively (Fig. 3). Of these, CIP exhibited the lowest binding affinity to MepA. CIP predominantly interacted within a pocket formed by amino acid residues ILE278, ILE40, ARG279, ARG281, PHE345, LEU344, THR345, and THR347.
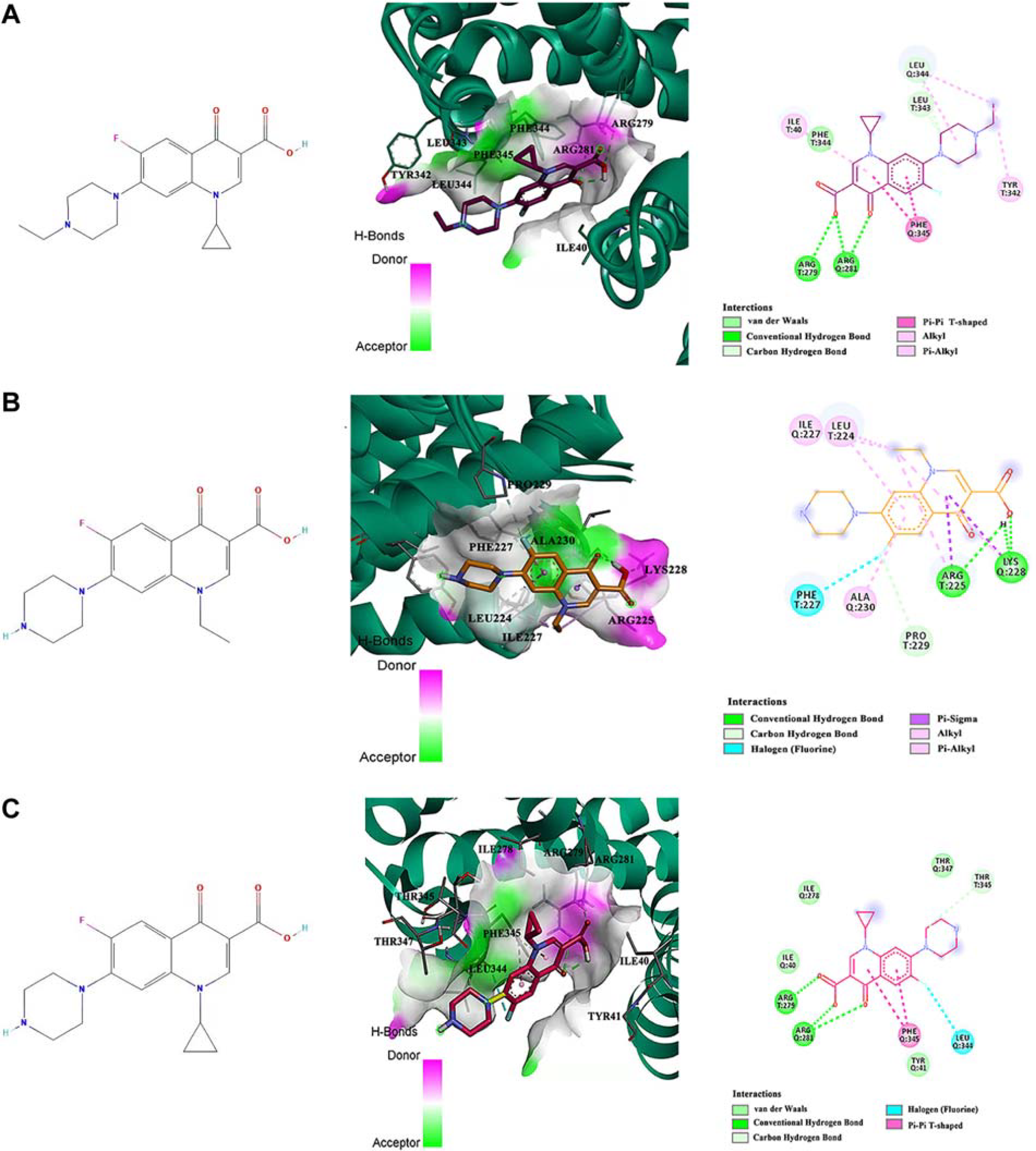
Optimal binding modes of MepA protein with ligand small molecules.
Analysis of antibiotic sensitivity and CRISPR loci changes of S. aureus under CIP pressure
Three CIP-susceptible S. aureus strains, namely SAXL24, WLD19, and LN25, were selected and induced to develop CIP resistance (Fig. 4A). Post-induction, three strains exhibited increased resistance to penicillin, AMP, and tetracycline (Fig. 4B–D). Furthermore, strains SAXL24/YD and WLD19/YD showed enhanced resistance to ceftiofur and sulfamethoxazole (Fig. 4E and F), while LN25/YD demonstrated increased resistance to gentamicin (Fig. 4G). The resistance levels against erythromycin, florfenicol, clindamycin, and streptomycin remained unchanged post-induction (Fig. 4H–K). Multiple sequence alignment revealed that both WLD19/YD and SAXL24/YD strains lost the first repeat and spacer sequences of their CRISPR loci (Supplementary Fig. S1A and B). Moreover, the relative mRNA expression levels of the CRISPR loci decreased after inducing antibiotic-sensitive S. aureus into resistant strains under antibiotic pressure (Supplementary Fig. S1C).
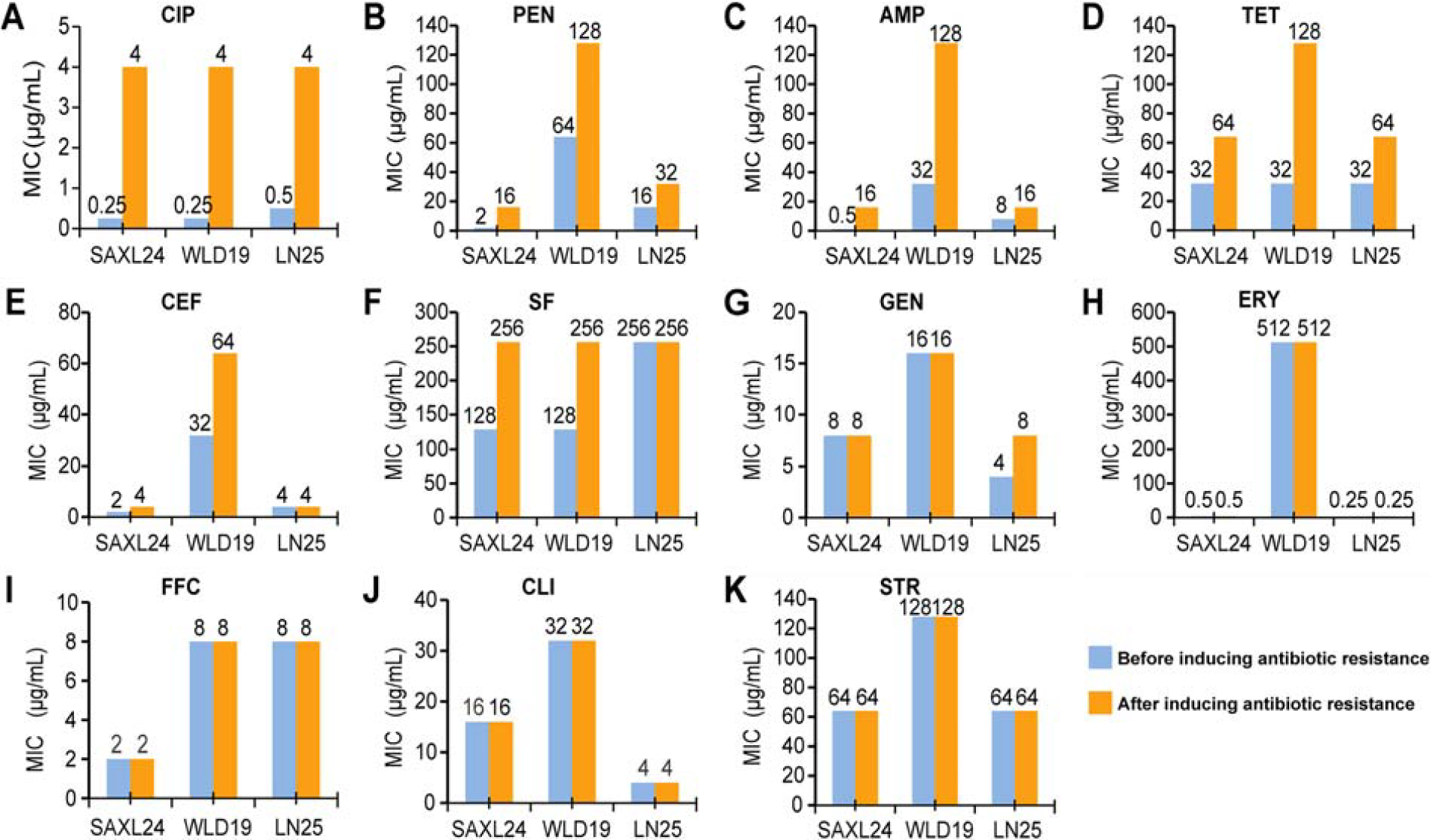
Minimum inhibitory concentration changes to various antibiotics after CIP-induced resistance in S. aureus strains. CIP, ciprofloxacin; S. aureus, Staphylococcus aureus.
CRISPR loci change in S. aureus after passage without antibiotic pressure
No significant changes were observed in the CRISPR-associated loci of S. aureus strains JY4 and M4 after 360 passages without antibiotic exposure (Supplementary Fig. S1D). The resistance of JY4/CD and M4/CD to ceftiofur, gentamicin, streptomycin, lincomycin, and florfenicol decreased, while their resistance to penicillin, tetracycline, and sulfamethoxazole remained highly stable (Table 4).
MIC Changes of S. aureus After Passage Without Antibiotic Pressure (µg/mL)

MIC, minimum inhibitory concentration.
Construction of pBT2-mepA and S. aureus 25-mepA
The mepA gene (1373 bp) was amplified from S. aureus-4 and successfully cloned into the pBT2 vector (6970 bp). Verification of the recombinant plasmid, pBT2-mepA, through SalI and BamHI digestion, yielded DNA fragments of 6970 bp and 1373 bp, confirming its successful construction (Supplementary Fig. S2A). Subsequently, the plasmid was extracted from RN4220 and electroporated into S. aureus-25, forming S. aureus-25-mepA. Enzyme digestion and electrophoresis confirmed fragment sizes of 6970 bp and 1373 bp, respectively (Supplementary Fig. S2B).
Sensitivity of S. aureus in different states to CIP
Planktonic, biofilm-associated, and resuspended biofilm bacteria of S. aureus-25-mepA, S. aureus-25, and S. aureus-4 were exposed to CIP for 1–4 h. The bacterial log reduction was more pronounced in planktonic and resuspended biofilm bacteria than in untreated controls, as shown in Figure 5A–C. Notably, after 4 h of treatment with 25 µg/mL CIP for the three strains, the log reduction in biofilm bacteria was significantly lower compared to planktonic bacteria. However, the reduction in resuspended biofilm bacteria was similar to that in planktonic bacteria, indicating a markedly reduced sensitivity of biofilm bacteria to CIP (p < 0.05). To further elucidate whether the reduced CIP sensitivity in biofilm bacteria is related to the efflux pump MepA, we assessed the transcriptional abundance of the mepA gene. Our findings revealed that the transcriptional abundance of mepA in S. aureus-4 and S. aureus-25-mepA biofilm bacteria did not significantly change under CIP pressure compared to controls (p > 0.05). In addition, the transcriptional abundance of mepA in biofilm bacteria of S. aureus-4 and S. aureus-25-mepA was significantly lower than that in planktonic bacteria (p < 0.01), indicating the pivotal role of biofilms in S. aureus resistance to CIP (Fig. 5D).
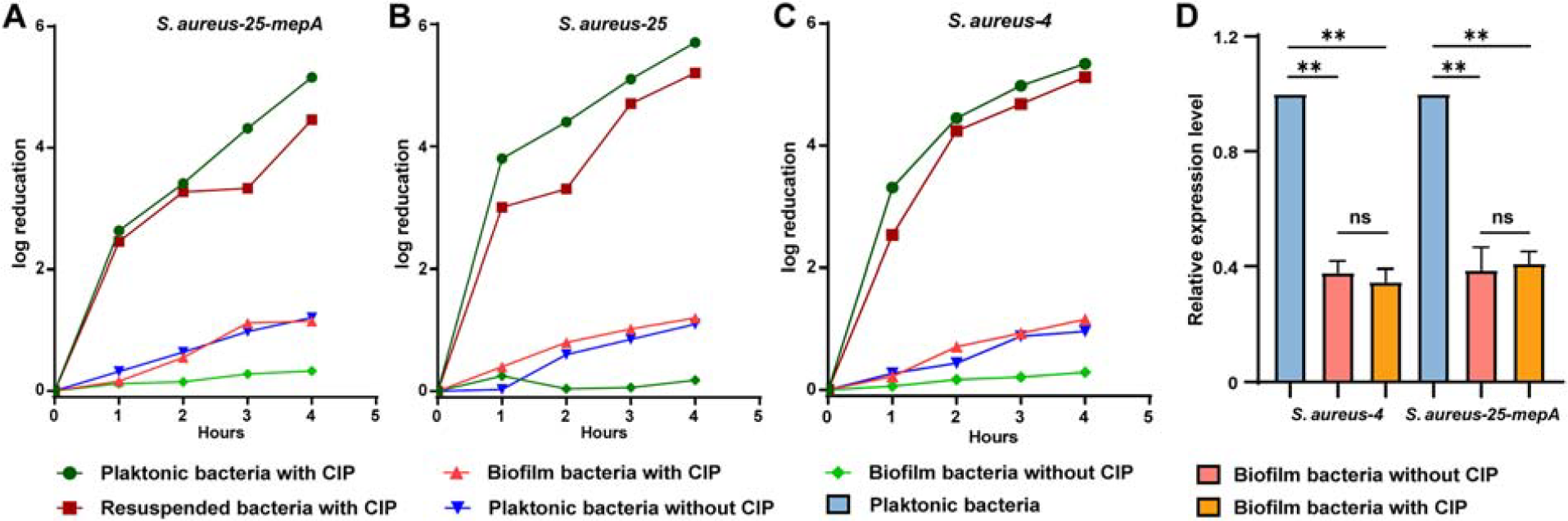
Bacterial reduction and mepA expression of biofilm-associated and planktonic S. aureus under CIP (25 µg/mL) stress.
Discussion
The CRISPR system, known for its high specificity and adaptability, utilizes various mechanisms to counteract antibiotic resistance, such as disrupting resistance-related genes (Mayorga-Ramos et al., 2023; Roy et al., 2021). In this study, only 7.69% of the 234 S. aureus isolates examined exhibited CRISPR loci, which is lower than that observed in other bacterial species (Makarova et al., 2015). This low detection rate may be due to S. aureus’s ability to suppress or lose CRISPR loci under environmental pressures essential for survival. Homology analysis of spacer sequences indicated that the spacer sequences of the CRISPR system in S. aureus were derived from exogenous genes. The length and diversity of spacer sequences can influence the activity of the CRISPR/Cas system (Tatiossian et al., 2021; Zhao et al., 2018). In addition, as a form of genetic memory, spacer sequences can provide evidence that mobile elements (such as phages and plasmids) have invaded bacteria in the past (Xie et al., 2020). This study identified 68 unique spacer sequences in 14 MRSA isolates, 32.31% of the spacer sequences targeted phages, suggesting that these MRSA had been assaulted by phages, which is evidence that bacteria resisted external invasion.
Previous research has indicated that the CRISPR sequence can remain invariant for 103–105 years (Dutta and Pan, 2002). Our study found that the CRISPR loci of multidrug-resistant S. aureus remained unchanged after 360 passages without antibiotic pressure. However, we observed the loss of the first repeat sequence and spacer sequences in the CRISPR loci of CIP-induced resistant S. aureus, along with decreased mRNA expression of CRISPR loci across all induced strains. Previous studies have reported that bacteria gain antibiotic-resistant genes by losing CRISPR loci, thereby enhancing antibiotic resistance and improving survival rates (Bikard et al., 2012; Mayorga-Ramos et al., 2023; Tatiossian et al., 2021). In our study, bacterial resistance to other antibiotics under CIP stress may be related to stress-induced mutagenesis, increased mRNA transcription of efflux pump genes, biofilm formation, and antibiotic cross-resistance (Zhai et al., 2023; Ballén et al., 2022; Nasrollahian et al., 2024).
Overexpression of MepA leads to resistance to fluoroquinolones and enhances bacterial survival in the antibiotic environment (Makarova et al., 2015; Vali et al., 2008). The present study found a high positive rate of the mepA gene (88.46%) in S. aureus, indicating that the mepA gene was prevalent in S. aureus in this region. Our results suggested that mepA mRNA expression was significantly higher in planktonic bacteria than in biofilm bacteria, potentially due to the reduced metabolic activity of biofilm bacteria (Guo et al., 2022). Bacteria in biofilms often mitigate the therapeutic effect of antibiotics by entering a dormant state or becoming persistent cells (Khan et al., 2020). In our study, the antibiotic sensitivity of S. aureus biofilm bacteria to CIP was lower than that of planktonic bacteria. Similar patterns were observed in Pseudomonas aeruginosa and S. epidermidis (Patel et al., 2022; Walters et al., 2003). However, there was no significant disparity in mepA expression level in biofilm bacteria before and after CIP exposure, indicating that S. aureus in the biofilm was lower sensitive to CIP. Similar results were identified in E. coli (Maira et al., 2000). In summary, planktonic S. aureus up-regulated the transcription level of the efflux pump gene mepA, thereby enhancing its resistance to CIP. Meanwhile, bacterial biofilms can protect against the bactericidal effect of CIP and enhance antibiotic resistance.
Conclusions
Our study found that the CRISPR carriage rate was 7.69% in 234 strains of S. aureus isolated from mastitis cows in Ningxia. Fluoroquinolone sensitivity may be associated with the loss of CRISPR repeats and spacers sequences and the downregulation of CRISPR mRNA transcription levels. This study explored the effects of the CRISPR system and the MepA efflux pump on antibiotic resistance in S. aureus under different conditions and elucidated the associated resistance mechanisms.
Authors’ Contributions
G.Q.W. and N.L. conceived and designed the experiments. G.J.W., P.P.L., and N.L. performed the experiments, analyzed the data, and wrote the original article. G.Q.W., X.H.C., and X.Q.H. participated in commenting, editing, and supervision of the article; All authors have finally read and approved the submission of the article.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded by the National Natural Science Foundation of China (Grant No. 32160852).