
Abstract
Antimicrobial-resistant Salmonella has posed a huge threat to food safety and public health, and tetracycline is the commonly used antibiotic for treating salmonellosis. In this study, eight Salmonella Typhimurium isolates from diarrhea patients (n = 7) and clam (n = 1) exhibited high-level tetracycline resistance (minimum inhibitory concentration = 128 μg/mL), and these isolates were further resistant to ampicillin and sulfonamides, formatting the R-type ASuT (ampicillin, sulfonamides, and tetracycline). Then, these eight isolates were sequenced using PacBio platform, revealing the presence of tetracycline resistance gene tet(B), along with sulfonamide-resistance gene sul2, extended-spectrum-β-lactamase gene blaTEM-1B, and aminoglycoside resistance genes aph(3″)-Ib and aph(6)-Id on the chromosome, which was associated with R-type ASuT. Four types (A, B1, B2, and C) of genetic arrangement for chromosomally encoding tet(B) were found, inserted into fljBA operon. Type C (fljBA operon-tet(B)-sul2-blaTEM-1B-aph(3″)-Ib-aph(6)-Id-merACDEPTR) was the most common type and was accompanied by various insertion sequences (ISs) (IS26, IS1, and ISVsa5) and recombinases. Pairwise sequence alignment showed that type C arrangement likely resulted from stepwise acquisitions and rearrangements facilitated by the actions of ISs, followed by integration into the chromosome by prophages. Phylogenomic analysis showed that all eight Salmonella Typhimurium isolates from China in this study, along with a human-borne Salmonella Typhimurium isolate (DA34821) from Germany and a foodborne Salmonella Typhimurium isolate (CFSA629) from China, clustered into a single clade, sharing ≤67 SNPs, which suggested that clone spread occurred. These findings underline the emergence of R-type ASuT in Salmonella Typhimurium, which is attributed to the presence of an antimicrobial resistance gene cluster (tet(B), sul2, blaTEM-1B, aph(3″)-Ib, and aph(6)-Id) encoded on the chromosome.
Introduction
The multidrug-resistant (MDR) Gram-negative bacteria are increasingly posing a huge threat to global food safety and public health. It was estimated that by 2050, ∼10 million people will die annually due to drug-resistant bacteria, unless a global response to the problem of antimicrobial resistance (AMR) is mounted (O’NeillJ, 2014). In 2019, antibiotic-resistant infections directly caused about 1.27 million deaths worldwide and indirectly caused about 4.95 million deaths (Vos et al., 2020). Among more than 2600 serovars of Salmonella, Salmonella Typhimurium was a leading cause of human gastroenteritis worldwide (Sana et al., 2016). Especially in China, Salmonella Typhimurium accounted for 69.1% of acute gastroenteritis among all ages of people (Liang et al., 2019). In recent years, the outbreaks of Salmonella Typhimurium causing gastroenteritis were frequently reported to originate from food contaminations such as meats, eggs, and chocolate products (Friesema et al., 2022; Larkin et al., 2022; Zhang et al., 2019). In April 2024, the outbreak of Salmonella Typhimurium linked to fresh basil resulted in 36 cases in the United States (https://www.cdc.gov/Salmonella/outbreaks/basil-04-24/?CDC_AAref_Val, available on January 26, 2025). The outbreak of MDR monophasic Salmonella Typhimurium (1,4,[5],12:i:-) associated with chocolate products resulted in at least 150 reported cases (by April 10, 2022) in EU/EEA countries (Germany, Sweden, France, Netherlands, Luxembourg, Norway, Ireland, Belgium, Sweden, and Spain) and the United Kingdom (Larkin et al., 2022). Tetracycline is commonly used to treat Salmonella infections. However, it was noted that high-level tetracycline resistance and the MDR had emerged in Salmonella Typhimurium. In China, 89.8% of Salmonella Typhimurium isolates exhibited resistance to tetracycline, and 91.1% of isolates exhibited MDR profiles (Dong et al., 2020). The classical MDR pattern in Salmonella Typhimurium isolates was ACSSuT (ampicillin, chloramphenicol, streptomycin, sulfonamides, and tetracycline), which ranged from 21.6% to 22.8% (Wang et al., 2017; Wei et al., 2019). The increase of tetracycline-resistant and MDR Salmonella Typhimurium isolates could pose formidable and growing menace for clinical treatments, and it is thus vital to uncover their development in Salmonella Typhimurium.
Currently, eight tet genes including tet(A), tet(B), tet(C), tet(G), tet(O), tet(M), tet(W), and tet(X) have been found to mediate tetracycline resistance in bacteria, of which, tet(A), tet(B), tet(C), and tet(G) code for energy-dependent efflux proteins, tet(O), tet(M), and tet(W) for ribosomal protection proteins, and tet(X) for a tetracycline-inactivating enzyme (Chopra and Roberts, 2001; Roberts, 2005). Furthermore, tet(A) and tet(B) were the most common ones in Salmonella Typhimurium isolates, accounting for 64.0% (Khan et al., 2019). In addition to tetracycline resistance, tet(B) also played an important role in the tigecycline resistance in Gram-negative isolates (Khlaif and Hussein, 2022). A previous study showed that 100% tigecycline-resistant Klebsiella pneumoniae, Acinetobacter baumannii, and Pseudomonas aeruginosa isolates harbored the tet(B) gene (Khlaif and Hussein, 2022). More importantly, the dissemination of tet(B) was primarily mediated by multiple types of transferred plasmids, including IncF2:A1:B1, IncFⅡ, IncA/C in Salmonella Typhimurium (Glenn et al., 2011; Guerra et al., 2002; Herrero et al., 2006; Liu et al., 2020). However, antimicrobial resistance genes (ARGs) were also frequently reported to be embedded into the chromosome (Partridge et al., 2018). For example, tet(M) has been also reported in the chromosome of marine bacteria, including Vibrio spp., Lactococcus garvieae, Bacillus spp., Lactobacillus spp., and Paenibacillus spp. (Neela et al., 2009).
In the present study, it was found that tet(B) was located on the chromosome as well as other ARGs (sul2, blaTEM-1B, aph(3″)-Ib, and aph(6)-Id) in Salmonella Typhimurium isolates from patients. To our knowledge, there is little information regarding how tet(B) horizontally incorporated into the chromosome of Salmonella spp. Here, we first found the emergence of chromosome-encoded ARG tet(B) in Salmonella Typhimurium, and characterized tet(B)-contained chromosome sequences to explore its embedding mechanism. In addition, whole genomes of Salmonella Typhimurium across the globe were selected to gain insights into the phylogenomic features of chromosome-encoded tet(B) isolates.
Materials and Methods
Bacterial isolates
The eight Salmonella Typhimurium isolates (SJTUF10231, SJTUF10057, SJTUF10169, SJTUF10484, SJTUF10250, SJTUF10330, SJTUF10648, and SJTUF11216) with high-level tetracycline resistance used in this study were available from our previous study (Zhang et al., 2024). SJTUF10484 was recovered from clam, and the rest were recovered from food and the stool of outpatients and inpatients in hospitals for diarrhea treatment. All the above Salmonella isolates were identified via API20E test strips (BioMerieux, France) and serotyped via commercial antiserum (Statens Serum Institute, Copenhagen, Denmark) according to the manufacturer’s guidelines.
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed on tetracycline-resistant Salmonella Typhimurium isolates using the agar dilution method provided by the Clinical and Laboratory Standard Institute (CLSI, 2019). The following antibiotics were selected: amikacin (AMK), ampicillin (AMP), ceftriaxone (CRO), cefoxitin (FOX), nalidixic acid (NAL), ciprofloxacin (CIP), ofloxacin (OFX), chloramphenicol (CHL), kanamycin (KAN), gentamicin (GEN), streptomycin (STR), sulfamethoxazole/trimethoprim (SXT), sulfisoxazole (FIS), azithromycin (AZM), fosfomycin (FOS), meropenem (MEM), and imipenem (IMP). Bacterial susceptibility to colistin (CT) was performed with the broth microdilution method recommended by the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2019). Escherichia coli ATCC 25922 and Enterococcus faecalis ATCC 29212 were used as quality control strains.
Whole-genome sequencing and assembly
These eight tetracycline-resistant Salmonella Typhimurium isolates were submitted to whole-genome sequencing (WGS) using the PacBio RS II platform. Salmonella Typhimurium cells were grown in 250 mL of liquid Luria–Bertani (LB) medium overnight at 37°C with agitation at 200 rpm. Following incubation, the cells were harvested and stored under specific conditions to maintain their integrity prior to DNA extraction. The storage conditions for the cell samples involved placing them in sterile containers at −80°C, which helps to preserve the cells and prevent degradation of their genomic material.
For DNA extraction, the QIAamp DNA mini kit (Qiagen, CA) was utilized following the manufacturer’s instructions with some additional details. Initially, the cell pellets were thawed on ice and resuspended in the appropriate buffer provided in the kit. Lysis of the cells was achieved through a combination of mechanical and chemical methods as per the kit protocol, ensuring complete release of the genomic DNA. The lysate was then processed through the provided columns, which bind the DNA while removing impurities such as proteins, RNA, and cellular debris.
After washing the columns to remove any residual contaminants, the genomic DNA was eluted in a buffer supplied with the kit. The eluted DNA was then quantified using spectrophotometric methods to assess its concentration and purity. Finally, the extracted genomic DNA was stored in aliquots at −20°C to prevent degradation.
The sequencing service was provided by the Personal Biotechnology Company (Shanghai, China). A 10-kbp DNA library was constructed and sequenced using single-molecule real-time sequencing technology. Sequence data from the PacBio RS II platform was assembled using the Canu software (Koren et al., 2017). Finally, the consensus genome sequence was determined using the Pilon software (Walker et al., 2014).
Annotation of the genome was performed using RAST (Overbeek et al., 2014), BLASTn, and BLASTp programs. The encoding genes in the genome were predicted by Glimmer (Delcher et al., 2007) and GeneMarkS (Besemer et al., 2001). The tRNAs, rRNAs, and repeated sequences in the genome were predicted by tRNAscan-SE v2.0 (http://trna.ucsc.edu/software/), Barrnap (https://github.com/tseemann/barrnap), and Tandem Repeats Finder (http://tandem.bu.edu/trf/trf.html), respectively.
Identification of antimicrobial resistance determinants, STs, plasmid types, and prophages
ResFinder 4.1 was used to identify ARGs and chromosomal mutations mediating antibiotic resistance in the genome (Bortolaia et al., 2020). MLST 2.0 was used to identify the sequence typings (STs) of bacteria (Larsen et al., 2012). Plasmidfinder was used to identify replicon types of plasmids (Carattoli et al., 2014). ISfinder (https://www-is.biotoul.fr/) was used to analyze the insertion sequences (ISs) and transposons in the genome. Prophage regions were identified using the PHASTER tool (http://phaster.ca).
Phylogenomic analysis
The genomic analysis was conducted using both de novo assemblies for our isolates and publicly available genomes retrieved from NCBI. We submitted the sequences (Region: 3789555.3841020 from SJTUF10231 genomes containing tet(B)-mer-sul2-blaTEM-1B-aph(3″)-Ib-aac(6′)-Iaa-aph(6)-Id) to NCBI public database, 55 genomes (≥45.00% coverage; ≥99.97% identity) were selected. A total of 63 genomes (Supplementary Table S1), including 8 genomes in this study and 55 genomes from NCBI, with a reference genome of Salmonella Typhimurium LT2 (Accession number NC_003197.2) were used for phylogenomic analysis to investigate their evolutional traces. Single-nucleotide polymorphisms (SNPs) were extracted using Snippy (https://github.com/tseemann/snippy) to generate core genomic alignment. Gubbins (Croucher et al., 2015) was then used to remove recombination regions. The core SNP alignment was used to generate a maximum-likelihood phylogeny using RAxML v8.1.23 (Stamatakis, 2014) with the GTR nucleotide substitution model. Furthermore, 100 random bootstrap replicates were conducted to assess the node support. The phylogenetic tree was visualized together with metadata using Interactive Tree Of Life (ITOL) stool (Letunic and Bork, 2021).
Data availability
The genome sequence data of eight Salmonella Typhimurium isolates have been deposited in the NCBI, which were available from BioProject ID: PRJNA484101 as follows: SJTUF10231 (Accession number CP047553), SJTUF10057 (Accession number CP047550), SJTUF10169 (Accession number CP047548), SJTUF10484 (Accession number CP047531), SJTUF10250 (Accession number CP047544), SJTUF10330 (Accession number CP047542), SJTUF10648 (Accession number CP047527), and SJTUF11216 (Accession number CP047522).
Results
Antimicrobial susceptibility results
The minimum inhibitory concentrations (MICs) of tetracycline (TET) in these eight isolates were 128 μg/mL (Table 1). In addition to tetracycline resistance, these isolates also exhibited resistance to ampicillin (AMP) (MIC ≥128 μg/mL), trimethoprim–sulfamethoxazole (SXT) (MIC ≥32/608 μg/mL), and sulfisoxazole (SUL) (MIC ≥1024 μg/mL). All these isolates displayed resistance to AMP, SUL, and TET (R-type ASuT).
Antimicrobial-Resistant Characteristics of Tet(B)-Producing Salmonella Typhimurium Isolates from Foods and Patients in Shanghai, China
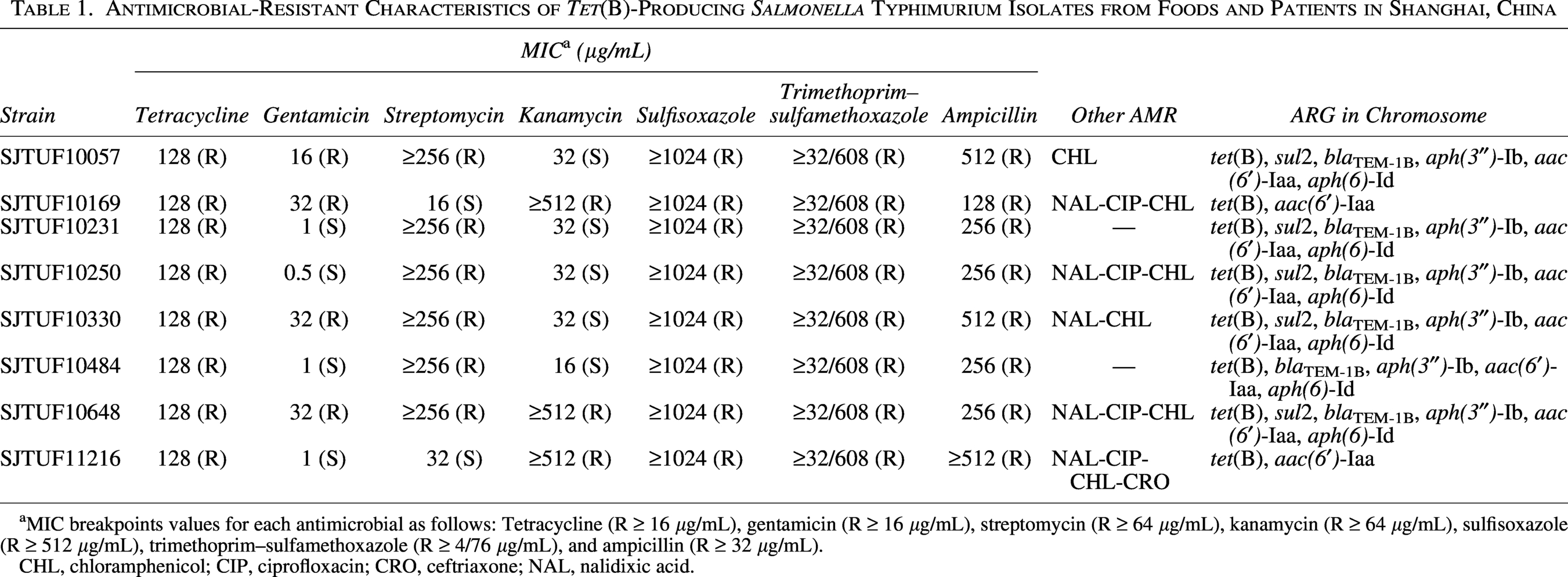
MIC breakpoints values for each antimicrobial as follows: Tetracycline (R ≥ 16 μg/mL), gentamicin (R ≥ 16 μg/mL), streptomycin (R ≥ 64 μg/mL), kanamycin (R ≥ 64 μg/mL), sulfisoxazole (R ≥ 512 μg/mL), trimethoprim–sulfamethoxazole (R ≥ 4/76 μg/mL), and ampicillin (R ≥ 32 μg/mL).
CHL, chloramphenicol; CIP, ciprofloxacin; CRO, ceftriaxone; NAL, nalidixic acid.
Furthermore, resistances to streptomycin, gentamicin, kanamycin, ciprofloxacin, chloramphenicol, and ceftriaxone were also observed among various isolates. For instance, five isolates were resistant to streptomycin, four to gentamicin, and three to kanamycin. Additionally, four isolates exhibited resistance to ciprofloxacin, six to chloramphenicol, and one to ceftriaxone. Beyond its high-level AMR in Salmonella, higher resistance to certain antimicrobial agents was also observed in China, such as cephems, quinolones, fluoroquinolones, lipopeptides, penicillins, and aminoglycosides (Hu et al., 2023). These findings highlight the growing problem of antimicrobial resistance in Salmonella Typhimurium and underscore the need for continued surveillance and monitoring of antibiotic resistance patterns.
ARGs in chromosome
These tetracycline-resistant isolates were then submitted to WGS. It was noted that four classes of ARGs were identified in chromosome. Tetracycline resistance gene tet(B) was identified in all these isolates. In addition to tet(B), sulfonamide-resistance gene sul2 and extended-spectrum-β-lactamase (ESBL) gene blaTEM-1B were identified in five and six isolates, respectively. Aminoglycoside resistance genes aac(6′)-Iaa (n = 8), aph(3″)-Ib (n = 6), and aph(6)-Id (n = 6) were also identified.
Four types (A, B1, B2, and C) of genetic arrangement of chromosomally encoding tet(B)
The chromosome sequences of these Salmonella Typhimurium isolates were separated from whole sequences using PacBio RS II platform. Four types (A, B1, B2, and C) of genetic arrangement of tet(B) were found (Fig. 1). A-type arrangement profile (flj-tet[B]) was composed of fljBA operon and ARG tet(B), which was identified in SJTUF11216 chromosome sequence (Fig. 1). B1-type arrangement profile (flj-tet(B)-mer) was composed of fljBA operon, tet(B), and heavy metal-resistant genes merACDEPTR, which was identified in SJTUF10169 chromosome sequence. B2-type arrangement profile (flj-tet(B)-aph-sul-bla) was composed of fljBA operon, tet(B), and blaTEM-1B, aph(3″)-Ib, and aph(6)-Id, which was identified in SJTUF10484 chromosome sequences. C-type arrangement profile (flj-tet(B)-aph-sul-bla-mer) was composed of fljBA operon, tet(B), blaTEM-1B, aph(3″)-Ib, aph(6)-Id, and merACDEPTR, which was identified in chromosome sequences of five isolates (SJTUF10231, SJTUF10057, SJTUF10250, SJTUF10330, and SJTUF10648).
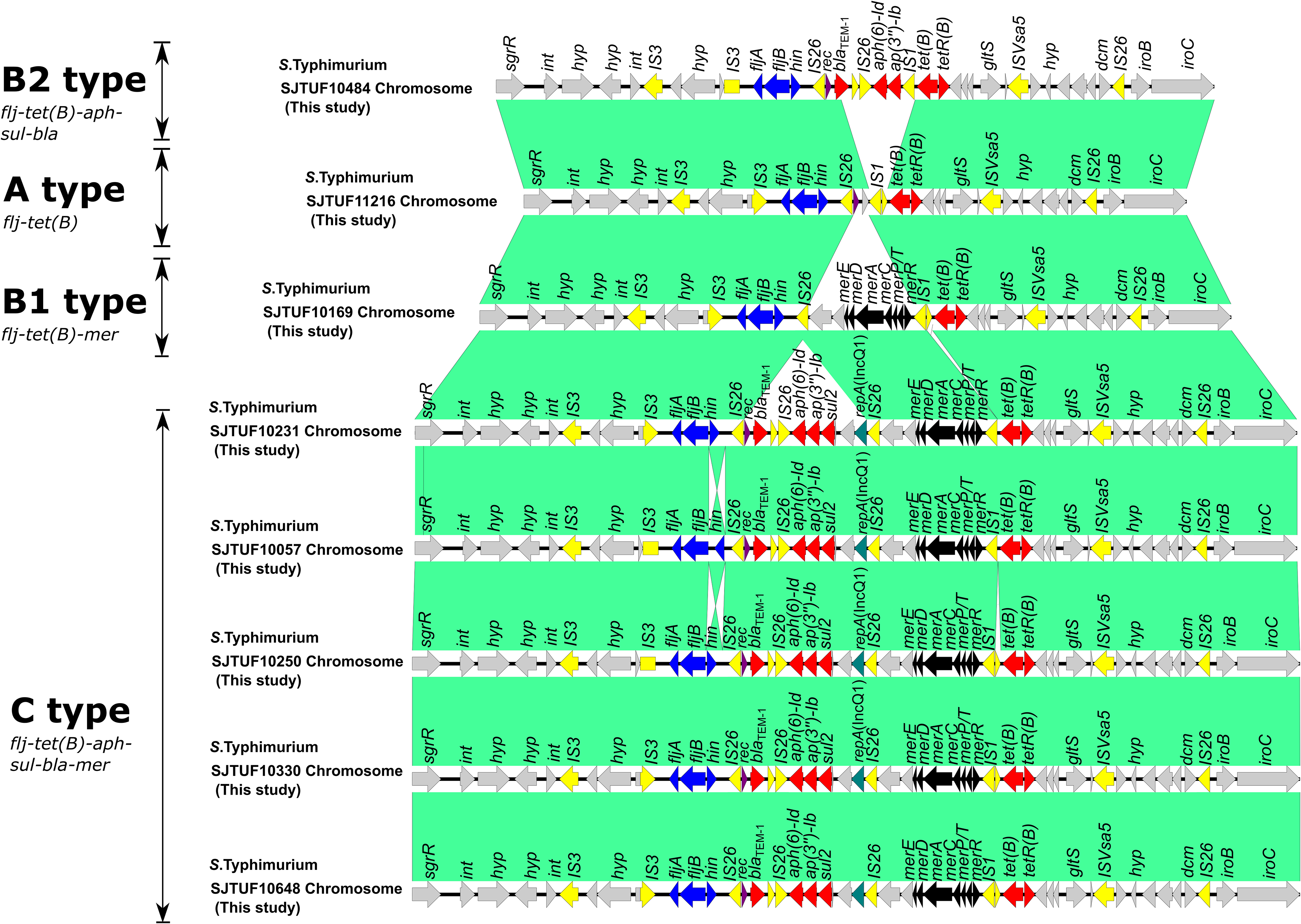
Four types (A, B1, B2, and C) of genetic arrangement of chromosomally encoding tet(B) in this study. Areas shaded in green indicate homologies between the corresponding genetic loci on each plasmid. Boxes or arrows represent the open reading frames (ORFs). Red represents antibiotic resistance genes; yellow, IS/transposase; black, heavy metal-resistant genes; purple, recombinant gene; gray, other genes. IS, insertion sequence.
Comparison of chromosome sequences containing tet(B)
We further compare chromosome sequences of SJTUF10484, SJTUF11216, SJTUF10169, SJTUF10231, and Salmonella Typhimurium LT2 isolates. One of the differences among them was the insertions of region I (~18.2 kb; SJTUF10484), II (~15.0 kb; SJTUF11216), III (~20.1 kb; SJTUF10169), and IV (~27.8 kb; SJTUF10231) (Fig. 2A). In particular, the ARGs, and/or heavy metal-resistant genes were contained in regions I–IV. Furthermore, these regions (I–IV) were inserted into the upstream of fljBA operon, which was associated with the expression of phase-2 flagellar antigen (Barco et al., 2014). Compared with that in Salmonella Typhimurium LT2 isolate, both ends of region I-IV were surrounded by inserted sequence IS26 element, and recombinase gene rec linking with ARG unit was further observed in SJTUF10169, SJTUF10231, SJTUF10057, SJTUF10250, SJTUF10330, and SJTUF10648 (Figs. 2A and 1), which suggested that insertion and recombination event might occurred in these isolates in this study. Region I composed of three ARG units containing IS26-blaTEM-1B-IS26, IS26- aph(3″)-Ib- aph(6)-Id -IS1, and IS1-tet(B)-ISVsa5. Region Ⅱ composed of one ARG unit containing IS1-tet(B)-ISVsa5. Region III composed of one ARG unit containing IS1-tet(B)-ISVsa5 and one heavy metal-resistant gene unit containing IS26-merACDEPT-IS1. Region IV composed of three ARG units containing IS26-blaTEM-1B-IS26, IS26- aph(6)-Id- aph(3″)-Ib-sul2-IS1, and IS1-tet(B)-ISVsa5, and one heavy metal-resistant gene unit containing IS26- merACDEPT-IS1. These results indicated that these regions containing ARG and heavy metal-resistant genes might evolve from independent resistant determinant through stepwise acquisitions and rearrangements by the actions of ISs and recombinase.

Refined comparison of antimicrobial resistance clusters in chromosome.
Particularly, region IV shared a homologous module with an Salmonella Typhi IncQ1-like plasmid pHCM1 (Accession number NZ_CP029939.1) and Salmonella 4,[5],12:i:- IncHI1 plasmid pB71 (Accession no NZ_KP899806.1). Furthermore, IncQ1 replicon gene repA was also found in SJTUF10231, SJTUF10057, SJTUF10250, SJTUF10330, and SJTUF10648 (Figs. 2A and 1). These results implied that these ARGs and heavy metal-resistant gene module in the chromosome originated from those on the plasmids. In addition, it was interesting that both the end of ARG module in pHCM1 and pB71 was IS1, suggested that homologous recombination might occur in IS1 as the common components of antibiotic resistance determinants.
We also identified an ~35.6 kb putative intact phage-containing ARG unit of blaTEM-1B, aph(6)-Id, aph(3″)-Ib, sul2, and tet(B) in the downstream of transcriptional regulator sgrR in the SJTUF10231 (Fig. 2B). Furthermore, integrase/transposase/recombinase was also identified in putative intact phage. Similar results were found in the chromosome of SJTUF11216. Thus, the ARG unit of blaTEM-1B, aph(6)-Id, aph(3″)-Ib, sul2, and tet(B) was probably integrated into the chromosome of the Salmonella Typhimurium by the prophages.
Phylogenomic analysis of Salmonella Typhimurium isolates carrying chromosomally encoding tet(B)
To further determine the phylogenetic characteristics of Salmonella Typhimurium isolates carrying chromosomally encoding tet(B), phylogenomic analysis was performed on 63 genomes from 14 countries during 1942–2018 (Fig. 3). The ARGs in the genomes were identified by ResFinder software. Phylogenomic results indicated that these Salmonella isolates could be divided into five clades (I–V). Clade I was composed of 10 Salmonella Saintpaul ST50 isolates, which showed a big genetic distance with other clades (Fig. 3). Only one isolate in clade I was identified to carry chromosomally encoding tetracycline resistance gene tet(A), not tet(B), and also possess sulfonamide-resistance gene sul1 as well as aminoglycoside resistance genes aac(6′)-Iaa, aph(3)-VIa, and aadA1. Clade Ⅱ was composed of 9 Salmonella Typhimurium isolates and one Salmonella Typhimurium var.5-isolates, and all these isolates belonged to ST19 (Fig. 3). It was noted that six isolates in clade Ⅱ were identified to carry chromosomally encoding tet(G), not tet(A) and tet(B). Furthermore, ESBL genes blaCARB-2 and AmpC β-lactamases gene blaCMY-2, as well as flufenicol resistance gene floR were also found in some isolates in clade Ⅱ. In addition to those in clade Ⅱ, floR was also found in one isolate from clade III. Clade IV was composed of 7 Salmonella Typhimurium isolates and 4 Salmonella Typhimurium var.5-isolates, and all these isolates belonged to ST19 (Fig. 3). It was noted that five Salmonella Typhimurium ST19 isolates in clade IV were identified to carry chromosomally encoding tet(A), sul1, aac(6′)-Iaa, aph(3)-VIa, and aadA1. AmpC β-lactamases gene blaCMY-2 was identified in four Salmonella Typhimurium var.5-isolates from clade IV.
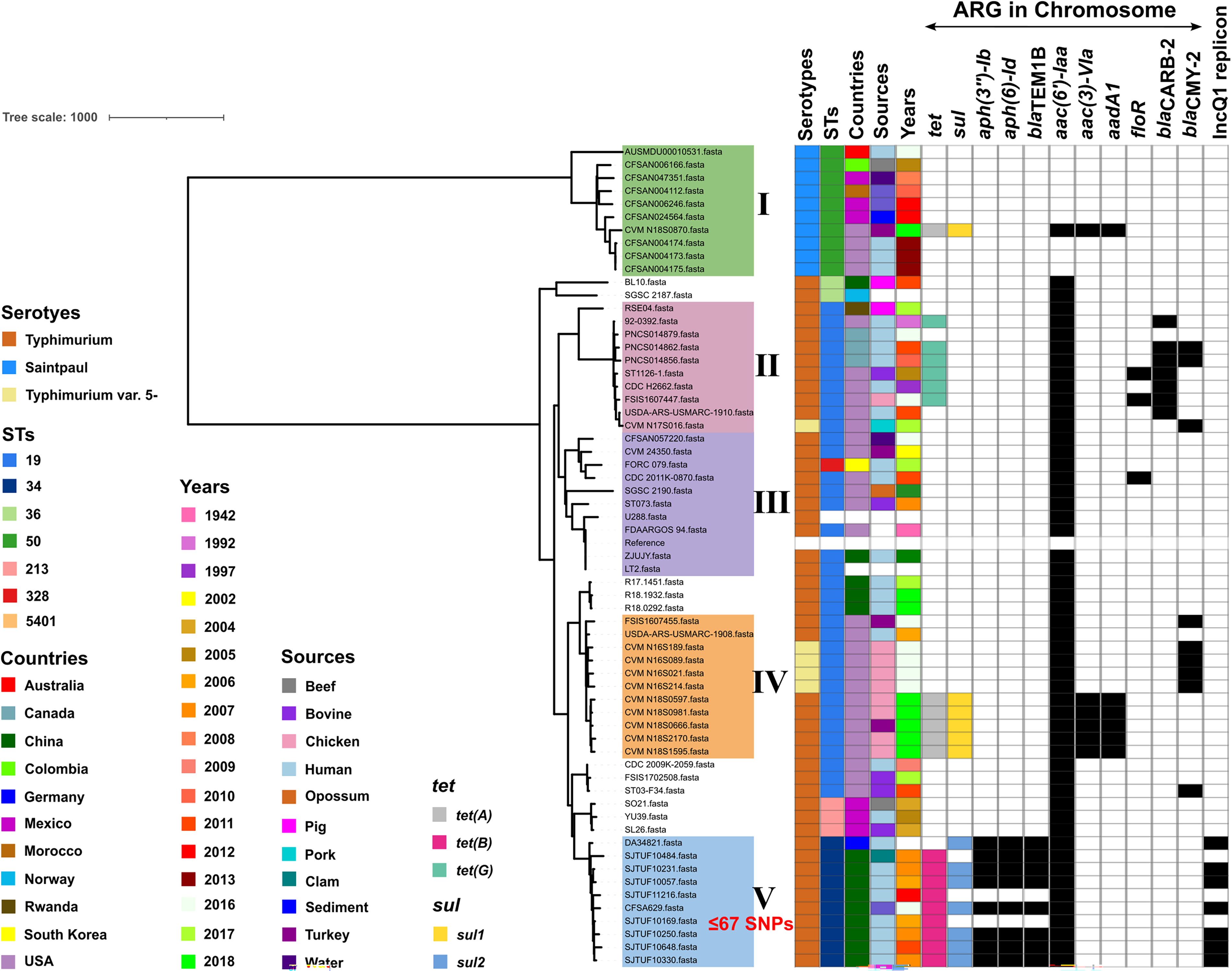
Phylogenomic analysis of Salmonella Typhimurium isolates bearing chromosome-encoded tet(B) in this study with other Salmonella genomes. The colors of isolate tips represent metadata columns including serotypes, STs, countries, sources, years, and ARG in chromosome (as shown in the inset legend). Light color shading shows Clades I–V. ARG, antimicrobial resistance gene.
Furthermore, all these eight Salmonella Typhimurium isolates in this study fell into clade V together with a human-borne Salmonella Typhimurium isolate (DA34821) from Germany and a foodborne Salmonella Typhimurium isolate (CFSA629) from China. All Salmonella Typhimurium isolates in clade V belonged to ST34, and shared ≤67 SNPs, suggesting their close genetic relationship, and the possibility of clone spread in isolates from foods and humans. Furthermore, Salmonella Typhimurium isolate DA34821 from Germany, which was the base of clade V, was not carried chromosomally encoding tet(B), but then tet(B) emerged in the rest isolates of clade V, which indicated that tet(B) might be captured later to enhance their antibiotic resistance for survive under the environmental pressure. It was interesting that IncQ1 replicon genes were identified in these chromosomally encoding tet(B) Salmonella Typhimurium isolates, but was absent in the other isolates, which suggested that IncQ1 replicon was special for Salmonella Typhimurium isolates carrying chromosomally encoding tet(B). In addition to tet(B), aminoglycoside resistance genes aph(6)-Id, aph(3)-VIa, and aadA1, sulfonamide-resistance gene sul2, as well as ESBL gene blaTEM-1B were also identified in isolates from clade V, which was also absent in those from other clades. Therefore, the emergence of Salmonella Typhimurium isolates carrying chromosomally encoding tet(B) as well as aph(6)-Id, aph(3)-VIa, aadA1, sul2, and blaTEM-1B occurred mainly in Shanghai, China, representing the regional uniqueness.
Discussions
Plasmids are primarily considered as a lead cause of the spread of ARG in Enterobacteriaceae isolates, including Salmonella spp. While ARGs are frequently integrated into chromosomes, the role of mobile genetic elements such as transposons in facilitating horizontal gene transfer warrants further investigation. In this study, these eight Salmonella Typhimurium isolates exhibited high-level resistance to tetracycline (MIC = 128 μg/mL); but any tetracycline resistance genes were not found in the plasmid, although that plasmids in our previous study were found to carry multiple ARGs such as sulfonamide-resistance gene sul1/3, extended-spectrum-β-lactamase (ESBL) gene blaOXA-1, and aminoglycoside resistance genes aac(6′)-Ib-cr, aac(3)-IV and aadA1/2 (Shang et al., 2021). Then it was noted that tetracycline resistance gene tet(B) was identified in the chromosome, and further joined to form a ARG cluster together with sulfonamide-resistance gene sul2, extended-spectrum-β-lactamase (ESBL) gene blaTEM-1B, and aminoglycoside resistance genes aph(3″)-Ib and aph(6)-Id, which has not been reported before to our knowledge. It was interesting that all these Salmonella Typhimurium isolates exhibited R-type ASuT (ampicillin, sulfonamides, and tetracycline), which was consistent with the AMR mediated by the chromosomal ARG cluster. The public health implications of our findings regarding the spread of R-type ASuT Salmonella Typhimurium are indeed significant. The discovery of a chromosomal ARG cluster containing multiple resistance genes in these isolates underscores the complexity and severity of the antimicrobial resistance issue. Such highly resistant strains pose a considerable threat to public health, as they can evade multiple classes of antibiotics, limiting therapeutic options. To prevent further dissemination of these resistant strains, it is crucial to implement strict antimicrobial stewardship programs in hospitals and clinics and enhance food safety practices to prevent contamination of food with resistant bacteria. This includes strict adherence to hygiene protocols during food production, processing, and handling.
Blast results showed that ARG cluster tet(B), aph(6)-Id, aph(3)-VIa, sul2, and blaTEM-1B was found in Salmonella Typhimurium ST34 isolates from China (7 human isolates and 2 food isolates) and Germany (a human isolate) (August 15, 2022). Phylogenomic results showed that these isolates fell in to a single clade V, and shared ≤67 SNPs, suggesting that clone spread occurred in isolates from foods and humans. The eight isolates had a high degree of genetic similarity, indicating that they are closely related genetically. This could be due to a recent common ancestor or shared genetic mutations. The close genetic relationship in these eight isolates might be the reason for their similar antimicrobial resistance phenotype. Some isolates (SJTUF10169, SJTUF10484, and SJTUF11216) also exhibited resistance to sulfisoxazole and trimethoprim–sulfamethoxazole despite lacking sul genes, which might be resulted from ARGs on the plasmids (Supplementary Table S2) reported in the previous study (Shang et al., 2021). The widespread use of antibiotics in animal farming to promote growth or prevent infections can select for resistant strains. These resistant clones can then enter the food chain through contaminated meat, dairy products, or eggs (Samtiya et al., 2022). Poor hygiene practices during food handling and processing can lead to cross-contamination (Zhang and Shi, 2024). This includes inadequate cleaning and sanitization of surfaces, equipment, and utensils (Zhang and Shi, 2024).
All these Salmonella Typhimurium isolates were submitted to PacBio RS II platform to characterize WGS, and then four genetic arrangement types of A (flj-tet(B)), B1 (flj-tet(B)-mer), B2 (flj-tet(B)-aph-sul-bla), and C (flj-tet(B)-aph-sul-bla-mer) were identified. These ARGs and heavy metal-resistant genes merACDEPTR inserted into the upstream of fljBA operon, but not disrupt it. Previous study showed that the genetic change of fljBA operon region like IS26 insertion would pose an effect on the expression of the phase 2 flagellin, and then generate multiple monophasic variants of Salmonella Typhimurium (Boland et al., 2015). For example, the disruption of fljBA operon by IS26 insertion linking to ARGs blaTEM-1, strAB, sul2, and tet(B) could result in the serovar 4,[5],12:i:- (Mu et al., 2022). Furthermore, recombinase and multiple ISs were identified in these four types of genetic arrangement regions, and it was apparent that C-type MDR genetic arrangement originated from ARG and heavy metal-resistant determinant region of A/B1/B2-type region through stepwise acquisitions and rearrangements with the help of recombinase and ISs such as IS26 and IS1.
Particularly, IncQ1-bearing MDR region IV containing multiply ISs shared by chromosomes (SJTUF10231, SJTUF10057, SJTUF10250, SJTUF10330, and SJTUF10648) and plasmids (Salmonella Typhi IncQ1-like plasmid pHCM1 and Salmonella 4,[5],12:i:- IncHI1 plasmid pB71), implying that this MDR region recombined occurred between chromosome and plasmids. Furthermore, an recombinase gene was found in the left end of ARG cluster tet(B), aph(6)-Id, aph(3)-VIa, sul2, and blaTEM-1B, and an ~35.6 kb region containing ARG cluster was identified as a putative intact phage. In the Acinetobacter, prophages can mediate the transfer of ARGs located on chromosomes between cells through transduction mechanism, providing a new perspective for understanding the dissemination of ARGs among Gram-negative pathogens (Wachino et al., 2019). The property of generalized transduction of temperate phages not only can bring antibiotic resistance to the host, but also can benefit the phage itself, presenting a mutualistic symbiotic relationship (Fillol-Salom et al., 2019). These relationships help the survival of bacteria and lead to the growth of the bacteriophage population (Fillol-Salom et al., 2019). Therefore, the MDR region IV into the chromosome of the Salmonella Typhimurium might be mediated by the prophages, and the direct evidence needs to be provided in the future.
Conclusions
In summary, our study highlighted the emergence of chromosome-encoded tet(B) with diverse ARGs, including blaTEM-1B, aph(6)-Id, aph(3″)-Ib, and sul2 in the Salmonella Typhimurium, which might be responsible for the formation of R-type ASuT. The formation of ARG cluster tet(B), aph(6)-Id, aph(3)-VIa, sul2, and blaTEM-1B was likely to be gradually integrated from a single ARG determinant unit by recombinase and ISs, and embedded into the chromosome with the help of the prophage. Great concerns should be taken for the co-spread of tet(B), aph(6)-Id, aph(3)-VIa, sul2, and blaTEM-1B.
Authors’ Contributions
Z.Z. contributed to conceptualization, methodology, data curation, formal analysis, writing—original draft, writing—review and editing, and funding acquisition. M.H. and K.A. contributed to writing—review and editing. C.S. contributed to resources, writing—original draft, writing—review and editing, supervision, project administration, and funding acquisition.
Footnotes
Funding Information
This research was supported by the Science and Technology Commission of Shanghai Municipality, China (grant number 21N31900200), and the National Natural Science Foundation of China (grant numbers 32202193 and 32472458).
Disclosure Statement
No interests to disclose.
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.