
Abstract
This study developed and optimized loop-mediated isothermal amplification (LAMP) assays for rapid detection of Salmonella and L. monocytogenes in fresh beef. LAMP primers targeting the invA gene of Salmonella and hly gene of L. monocytogenes were used. Reaction conditions including temperature, dNTP concentration, Mg2+ concentration, primer ratio, and Bst DNA polymerase amount were optimized for each pathogen. Differences were observed between the two pathogens in optimal temperature, Mg2+ concentration, and Bst DNA polymerase requirements, highlighting the importance of pathogen-specific optimization. The optimized assays demonstrated high sensitivity with detection limits of 120 fg/μL for Salmonella and 130 fg/μL for L. monocytogenes, achieving detection within 40 and 60 min, respectively. Specificity tests confirmed both assays were highly specific for their target pathogens in fresh beef samples with no cross-reactivity observed. Addition of hydroxynaphthol blue enabled simple visual detection of positive results, eliminating the need for specialized equipment. The developed LAMP assays offer rapid, sensitive, and specific detection of these important foodborne pathogens.
Introduction
Foodborne pathogens pose significant risks to public health and food safety globally. Among these, Salmonella and L. monocytogenes are major concerns in meat products, including fresh beef (Ali and Alsayeqh, 2022; Warmate and Onarinde, 2023). Salmonella is a Gram-negative bacterium responsible for salmonellosis, causing symptoms such as diarrhea, abdominal cramps, and fever (Ehuwa et al., 2021). L. monocytogenes is a Gram-positive bacterium that can cause listeriosis, a severe illness with high mortality rates, especially in vulnerable populations (Dos Reis et al., 2024). The prevalence of L. monocytogenes in beef and beef products can be substantial, with some studies reporting contamination rates of up to 60% (Dos Reis et al., 2024).
The incidence of foodborne diseases has been increasing in recent years, particularly in industrialized countries, due to changing eating habits and rising demand for ready-to-eat foods (Zhao et al., 2014). Fresh beef, being a nutrient-rich medium, provides an ideal environment for the growth of these pathogens. Contamination can occur at various stages of the production chain, from slaughter to processing and packaging, making it crucial to have effective detection methods in place (Rhoades et al., 2009).
Traditional culture-based methods for pathogen detection are labor-intensive, require skilled personnel, and are time-consuming, often taking 5–7 d for confirmation, although they do provide viable cell counts (Dwivedi and Jaykus, 2011). Immunological methods, such as enzyme-linked immunosorbent assay (ELISA), provide high specificity and faster detection compared with culture methods, but may have lower sensitivity than molecular methods such as polymerase chain reaction (PCR), and can be time-consuming for certain techniques (Law et al., 2014). Among molecular methods, PCR offers high sensitivity and rapid results, capable of detecting low pathogen concentrations, but requires specialized equipment (Chen et al., 2022). Quantitative real-time PCR has further advanced detection by enabling real-time monitoring and quantification without post-amplification processing (Salihah et al., 2016), whereas digital PCR provides absolute quantification through sample partitioning, eliminating standard curves and showing greater tolerance to food matrix inhibitors (Gutiérrez-Aguirre et al., 2015). Despite these advantages, all PCR-based methods require thermal cycling equipment, skilled personnel, and complex sample preparation, limiting their application in resource-constrained settings or field-testing scenarios.
Loop-mediated isothermal amplification (LAMP) is a novel nucleic acid amplification technique that offers several advantages over traditional PCR and other rapid detection methods (Law et al., 2014). LAMP operates at a constant temperature, typically between 60°C and 65°C, eliminating the need for thermal cycling equipment (Yang et al., 2018). It is highly specific due to the use of four to six primers recognizing six to eight distinct regions on the target gene (Yang et al., 2018). This unique amplification mechanism results in the production of large amounts of DNA, allowing for easy detection by turbidity or fluorescence (Yang et al., 2018). For visualization in LAMP assays, several colorimetric indicators have been developed, including SYBR Green I, calcein, berberine, EvaGreen, and hydroxynaphthol blue (HNB) (Fischbach et al., 2015). In this study, we selected HNB as the colorimetric indicator due to its ability to provide clear visual discrimination between positive (sky blue) and negative (violet) results (Fischbach et al., 2015).
LAMP is also more tolerant to biological substances that typically inhibit PCR, making it suitable for complex food matrices (Francois et al., 2011). The technique has shown promise in detecting various foodborne pathogens, such as Staphylococcus aureus, Vibrio parahaemolyticus, and Escherichia coli (Lakshmi and Kim, 2021). However, the application of LAMP for simultaneous detection of multiple pathogens in fresh beef has not been extensively studied.
The invA gene, encoding a protein essential for Salmonella invasion of epithelial cells, is highly conserved among Salmonella species and serves as an ideal target for detection (Mohammed, 2022). For L. monocytogenes, the hly gene encoding listeriolysin O is a well-established virulence factor and detection target (Gasanov et al., 2005). Both genes have been successfully used in previous PCR and LAMP-based detection methods (Fiore et al., 2023; Yang et al., 2018).
This study aimed to develop and optimize LAMP assays for the rapid detection of Salmonella and L. monocytogenes in fresh beef. We used LAMP primers targeting the invA and hly genes, optimized reaction conditions, and evaluated the sensitivity and specificity of the assay. In addition, we incorporated a colorimetric detection method using HNB to enable visual result interpretation without specialized equipment.
Materials and Methods
Bacterial strains and DNA extraction
Salmonella enterica (CICC 21482), L. monocytogenes (ATCC 19115), S. flexneri (CMCC 51302), Staphylococcus aureus (CICC 21600), and Escherichia coli (CICC 10389) were used in this study. All strains were cultured in Luria-Bertani (LB) broth at 37°C with shaking at 180 rpm for 18 h. Bacterial DNA was extracted using a commercial DNA extraction kit (Sangon Biotech Co., Ltd., Shanghai, China) according to the manufacturer’s instructions. All extracted DNA concentrations and purity were determined using spectrophotometry. DNA samples were stored at −20°C until use.
LAMP primers
LAMP primers (Jiang et al., 2013) were designed based on the invA gene of Salmonella enterica (National Center for Biotechnology Information [NCBI] Reference Sequence: NC_003197) (McClelland et al., 2001) and the hly gene of L. monocytogenes (NCBI Reference Sequence: NC_003210) (Toledo-Arana et al., 2009). The invA gene was selected as it encodes an invasion protein essential for Salmonella pathogenicity and is highly conserved. The hly gene, encoding listeriolysin O, was chosen as it is a well-characterized virulence factor specific to L. monocytogenes. For each target, four primers were used: two outer primers (F3 and B3) and two inner primers (FIP and BIP). Primer sequences are listed in Supplementary Table S1.
Optimization of LAMP reaction conditions
LAMP reactions were optimized separately for Salmonella and L. monocytogenes. Starting from an initial reaction condition containing F3 and B3 (10 μM, 0.5 μL each), FIP and BIP (40 μM, 1.0 μL each), dNTP (10 mM, 2.5 μL), MgSO4 (100 mM, 2.5 μL), 10× buffer (2.5 μL), Bst DNA polymerase (8U, 1.0 μL) (Sangon Biotech Co., Ltd.), and template DNA (1.0 μL) in a 25 μL reaction volume, several parameters were systematically optimized. These included reaction temperature (60°C, 61°C, 62°C, 63°C, 64°C, and 65°C), dNTP concentration (0.6 mmol/L, 0.8 mmol/L, 1.0 mmol/L, 1.2 mmol/L, and 1.4 mmol/L), Mg2+ concentration (6 mmol/L, 8 mmol/L, 10 mmol/L, 12 mmol/L, and 14 mmol/L), inner to outer primer ratio (4:1, 8:1, 16:1, and 32:1), and Bst DNA polymerase amount (0.8 μL, 0.9 μL, 1.0 μL, 1.1 μL, and 1.2 μL). The optimized conditions were established for each pathogen separately and are detailed in the section “Result.” LAMP reactions were incubated in a water bath at specified temperature. Results were assessed by visual inspection for turbidity and confirmed by agarose gel electrophoresis with 4S GelRed nucleic acid staining (Sangon Biotech Co., Ltd.).
Sensitivity testing
The sensitivity of the optimized LAMP assays was determined using 10-fold serial dilutions of genomic DNA from Salmonella and L. monocytogenes, ranging from 120 ng/μL to 12 fg/μL and 130 ng/μL to 13 fg/μL, respectively.
Application to artificially contaminated samples
This part of the experiment was conducted based on previously established methods with modifications (Fiore et al., 2023). Fresh beef samples (25 g) were artificially contaminated with bacteria. After homogenization, 10 μL of each homogenate was inoculated into LB broth and cultured at 37°C for 14 h. After enrichment, DNA was extracted from the samples using the DNA extraction kit (Sangon Biotech Co., Ltd.) for LAMP analysis. Non-contaminated samples served as negative controls, and samples contaminated with non-target bacteria, including S. flexneri, S. aureus, and E. coli, were used as additional controls.
Colorimetric LAMP detection
HNB was added to the LAMP reaction at a final concentration of 19.2 μmol/L to enable visual detection. A color change from violet to sky blue indicated a positive reaction. The color change was observed by the naked eye and photographed.
Results and Discussion
Optimization of Salmonella LAMP assay
The experimental design of this study is shown in Figure 1. In the Salmonella LAMP assay, the effect of reaction temperature on amplification was investigated using a range from 60°C to 65°C (Fig. 2A). No visible turbidity was observed in any reaction tubes at 30 min. At 40 min, visible turbidity appeared in tubes 2 (61°C) and 5 (64°C), followed by tubes 1 (60°C), 3 (62°C), 4 (63°C), and 6 (65°C) showing turbidity at 50 min. No turbidity was observed in the negative control throughout the reaction. Although both 61°C and 64°C showed early turbidity at 40 min, 64°C was selected as the optimal temperature. This choice was based on two considerations: first, 64°C is closer to the optimal working temperature of Bst DNA polymerase (65°C), which ensures maximum enzyme activity; second, higher temperatures generally provide better specificity in nucleic acid amplification by reducing non-specific primer binding (Jang and Kim, 2022). The observation that amplification occurred across all tested temperatures (60–65°C) demonstrates the robust nature of LAMP reactions, which is advantageous for practical applications where precise temperature control might be challenging (Cai et al., 2018).

The flowchart for the experimental design of this study.
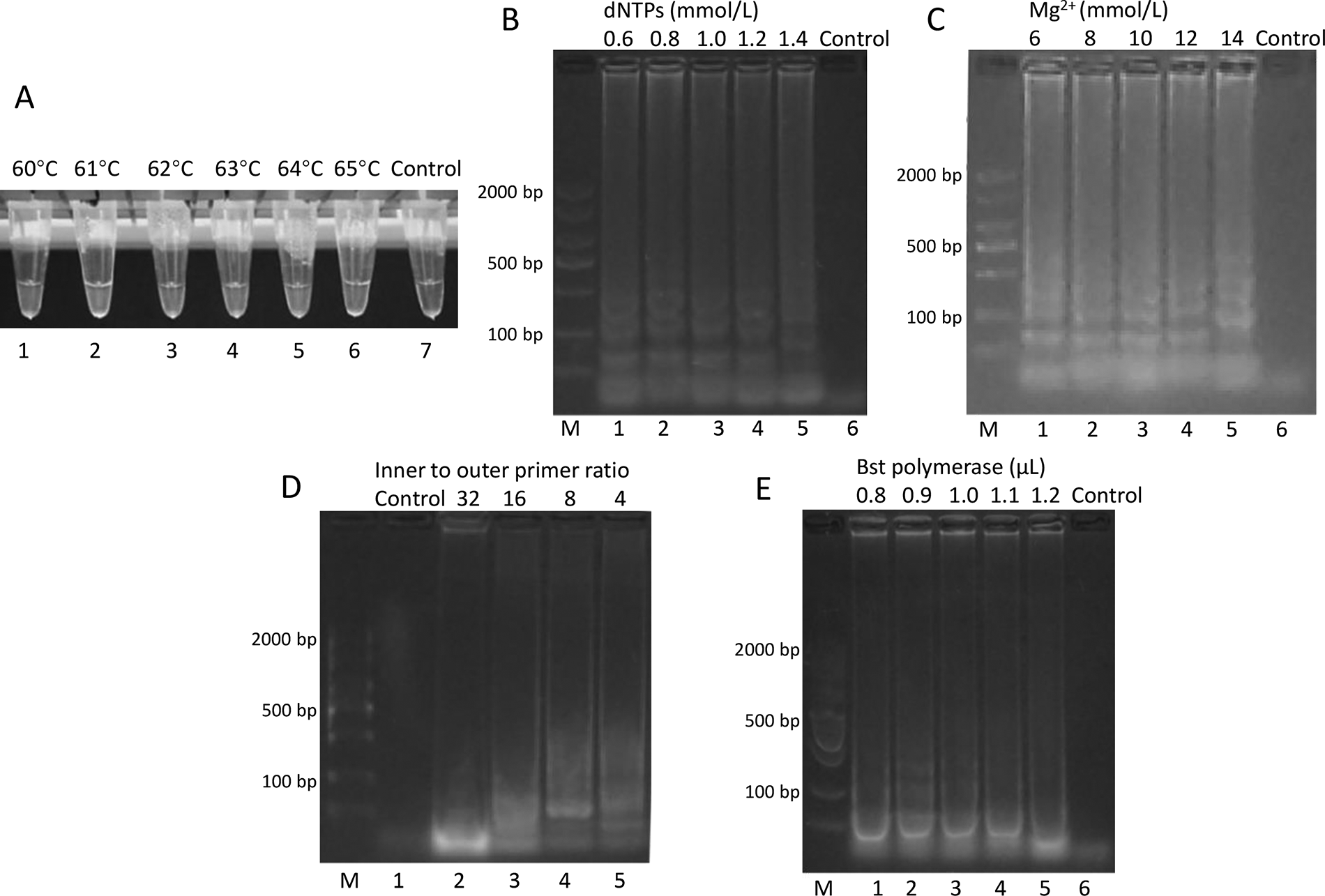
Optimization of LAMP reaction conditions for Salmonella detection.
The optimization of other reaction parameters was evaluated by agarose gel electrophoresis. For dNTP concentration (Fig. 2B), although amplification occurred across all tested concentrations (0.6–1.4 mmol/L), ladder-like patterns were most obvious at 1.0 mmol/L. Mg2+ concentration was tested from 6 to 14 mmol/L (Fig. 2C), with 10 mmol/L showing the strongest ladder-like patterns. The inner to outer primer ratio optimization (Fig. 2D) revealed 8:1 as optimal, producing the most obvious ladder-like bands. For Bst DNA polymerase amount (Fig. 2E), 0.9 μL yielded the best amplification results among the tested volumes (0.8–1.2 μL). Therefore, the optimal conditions for the Salmonella LAMP assay were determined to be 64°C reaction temperature, 1.0 mmol/L dNTPs, 10 mmol/L Mg2+, 8:1 inner to outer primer ratio, and 0.9 μL Bst DNA polymerase. Under these conditions, positive results were obtained within 40 min (Supplementary Table S2).
Optimization of L. monocytogenes LAMP assay
In the L. monocytogenes LAMP assay, reaction temperature was first optimized from 60°C to 65°C (Fig. 3A). No visible turbidity was observed in any reaction tubes at 50 min. At 60 min, tube 4 (63°C) showed turbidity, followed by tubes 3, 5, and 6 (62°C, 64°C, and 65°C) at 80 min. Although multiple temperatures supported amplification, 63°C was selected as the optimal temperature due to its earliest detection time.
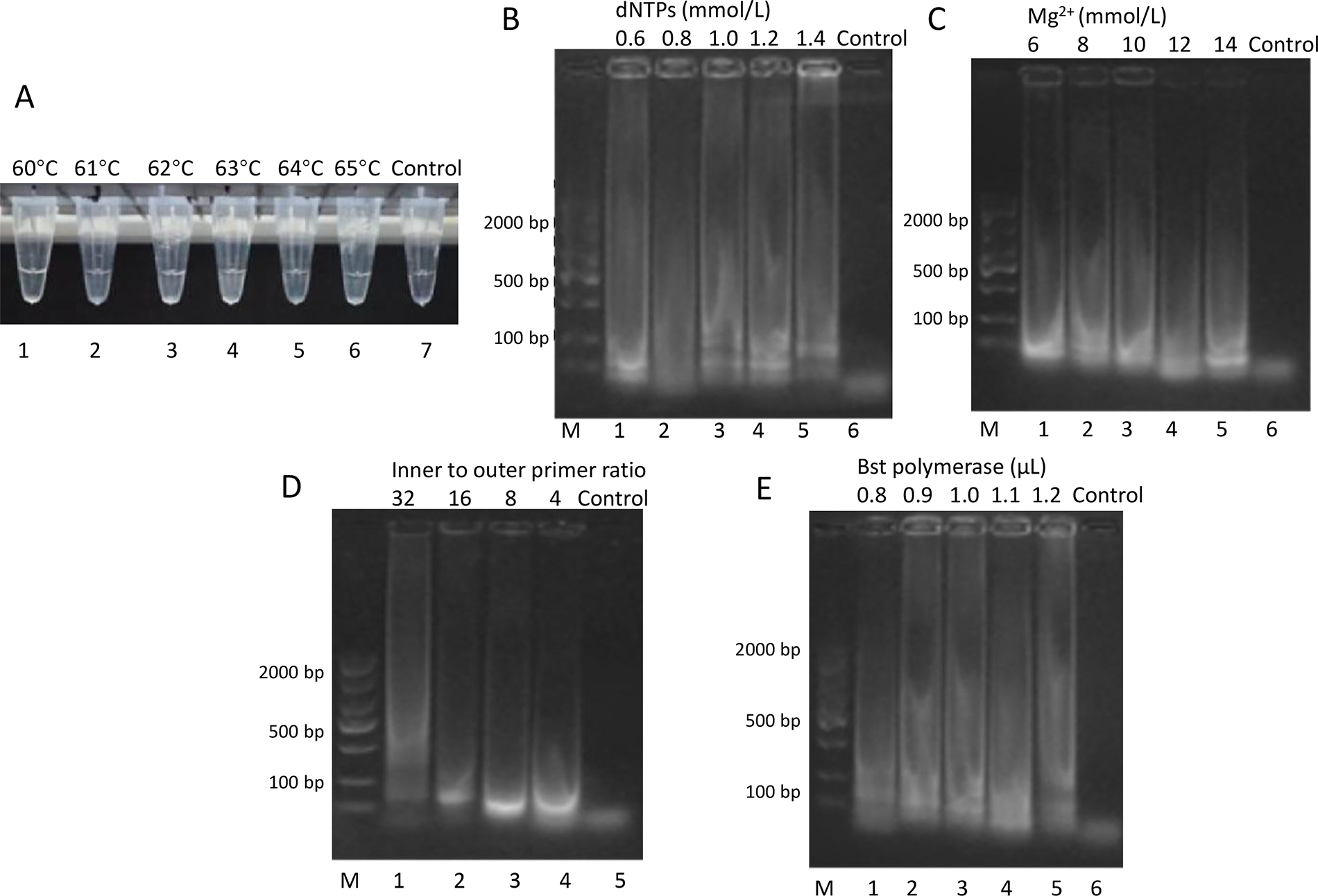
Optimization of LAMP reaction conditions for Listeria monocytogenes detection.
The optimization of other reaction parameters was evaluated by agarose gel electrophoresis. For dNTP concentration (Fig. 3B), ladder-like patterns of LAMP amplification were observed, with the strongest amplification at 1.0 mmol/L. Similar ladder-like patterns were seen in Mg2+ optimization (Fig. 3C), where 8 mmol/L produced the most intense amplification. The inner to outer primer ratio of 8:1 (Fig. 3D) and Bst DNA polymerase amount of 1.0 μL (Fig. 3E) were selected based on the intensity of the LAMP amplification patterns. Therefore, for the L. monocytogenes LAMP assay, the optimal conditions were 63°C reaction temperature, 1.0 mmol/L dNTPs, 8 mmol/L Mg2+, 8:1 inner to outer primer ratio, and 1.0 μL Bst DNA polymerase. Positive results were obtained within 60 min under these conditions (Supplementary Table S3).
The observed variations in amplification timing at different temperatures may be explained by the temperature-dependent nature of LAMP reactions (Shirshikov and Bespyatykh, 2022). Small temperature changes can affect enzyme kinetics and DNA structural dynamics (Shirshikov and Bespyatykh, 2022), leading to the non-linear relationship between temperature and amplification timing observed in our study. Standard LAMP reactions typically utilize temperatures between 60°C and 65°C (Shirshikov and Bespyatykh, 2022), and our optimal temperatures (64°C for Salmonella and 63°C for L. monocytogenes) fall within this range. The optimized Mg2+ concentrations were determined to be 10 mmol/L for Salmonella and 8 mmol/L for L. monocytogenes, whereas the optimal inner to outer primer ratio was 8:1 for both assays. These findings demonstrate that each LAMP assay may require specific optimization of reaction parameters for maximum efficiency and specificity.
Sensitivity and specificity of LAMP assays
The sensitivity and specificity of the optimized LAMP assays were evaluated (Fig. 4). For Salmonella, the ladder-like pattern of LAMP amplification was observed down to 120 fg/μL (Fig. 4A). No amplification was detected at 12 fg/μL or in the negative control. Similarly, for L. monocytogenes, positive amplification was observed down to 130 fg/μL (Fig. 4B), whereas no amplification occurred at 13 fg/μL or in the negative control.
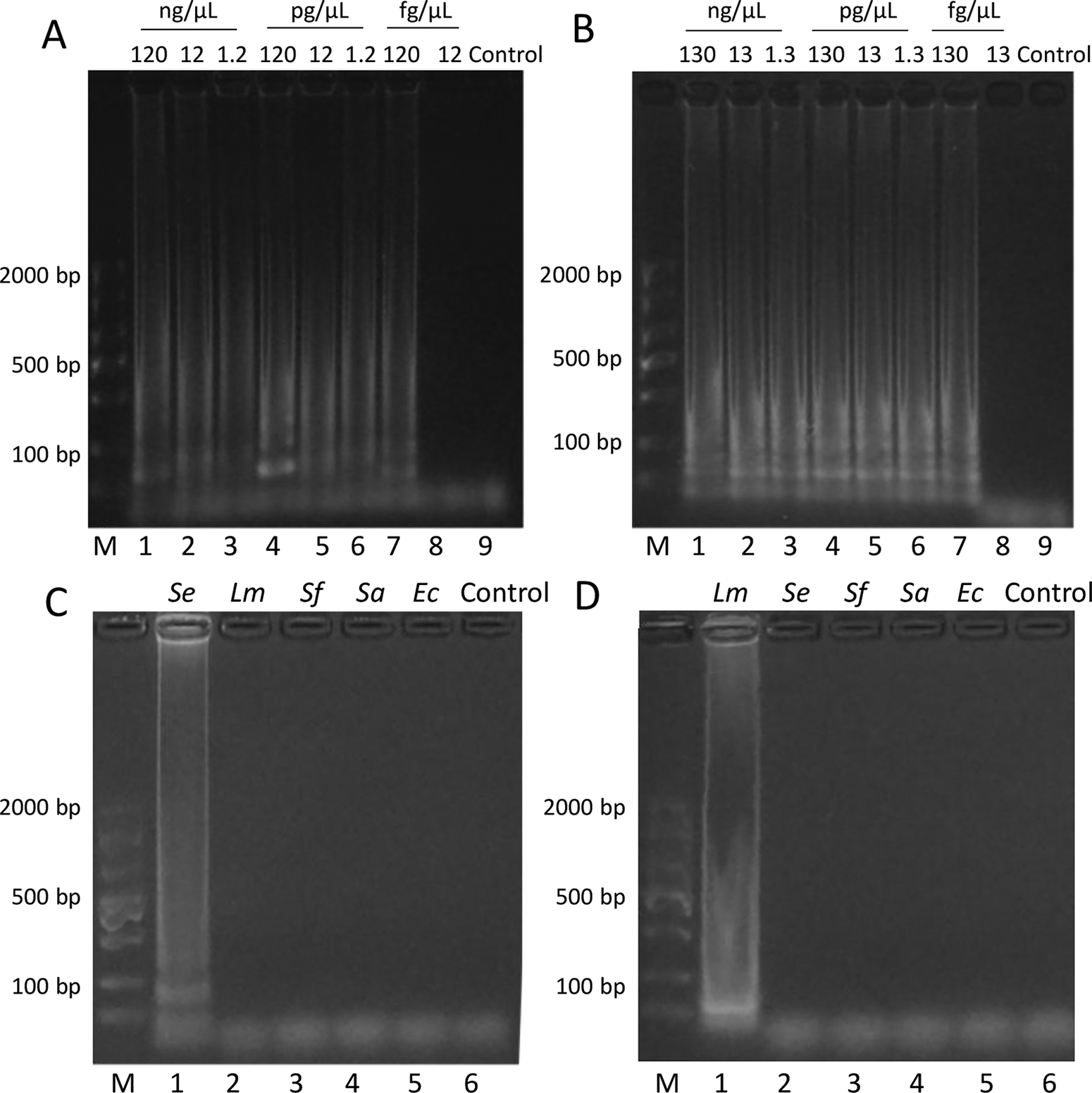
Sensitivity and specificity of LAMP assays.
The specificity of both assays was evaluated using DNA extracted from artificially contaminated beef samples. For Salmonella, amplification was only observed in samples contaminated with Salmonella (Fig. 4C, lane 1), whereas no amplification was detected in samples containing L. monocytogenes, S. flexneri, S. aureus, and E. coli (lanes 2–5). Similarly, the L. monocytogenes assay showed positive amplification only with L. monocytogenes-contaminated samples (Fig. 4D, lane 1), with no cross-reactivity observed with other tested pathogens (lanes 2–5).
These results demonstrate the sensitivity and specificity of our LAMP assays. The detection limits achieved (120–130 fg/μL) are comparable with those reported in previous LAMP-based detection methods (Wang et al., 2008; Wang et al., 2015). The high specificity, evidenced by the absence of cross-reactivity with other common foodborne pathogens, can be attributed to the use of multiple primers recognizing distinct regions of the target genes (Wong et al., 2018). Furthermore, the successful detection in beef samples indicates the robustness of these assays against potential inhibitors present in food matrices, making them suitable for practical food safety applications.
The gel electrophoresis images in Figures 2–4 display the characteristic ladder-like pattern typical of successful LAMP amplification. This pattern results from the formation of stem-loop DNA structures with inverted repeats of the target sequence, producing amplicons of various sizes (Notomi et al., 2000). For both the Salmonella and L. monocytogenes LAMP assays (Figs. 2B–E and 3B–E, respectively), the most distinct ladder-like patterns were observed under the optimized conditions. In the sensitivity analysis (Fig. 4A and B), clear ladder patterns are visible down to the detection limits of 120 fg/μL for Salmonella and 130 fg/μL for L. monocytogenes, with no amplification bands observed below these concentrations or in the negative controls. The specificity gels (Fig. 4C and D) show clear ladder patterns only in lanes containing the target pathogen, with no amplification bands in non-target pathogens or negative controls, confirming assay specificity.
Colorimetric detection
The visual detection capability of the LAMP assays was evaluated using HNB as a colorimetric indicator (Fig. 5). For both Salmonella and L. monocytogenes assays, positive reactions showed a distinct color change from violet to sky blue (Fig. 5A and B, tube 1), whereas negative controls remained violet (tube 2). HNB functions as a metal ion indicator, changing color in response to the concentration of free Mg2+ in the reaction mixture (Goto et al., 2009). During DNA synthesis, dNTP consumption leads to chelation of magnesium ions, reducing free Mg2+ concentration. This alters the HNB environment, causing a visible color change from violet to sky blue in positive reactions. The clear visual distinction between positive and negative results demonstrates the potential of these assays for rapid, equipment-free detection of target pathogens.
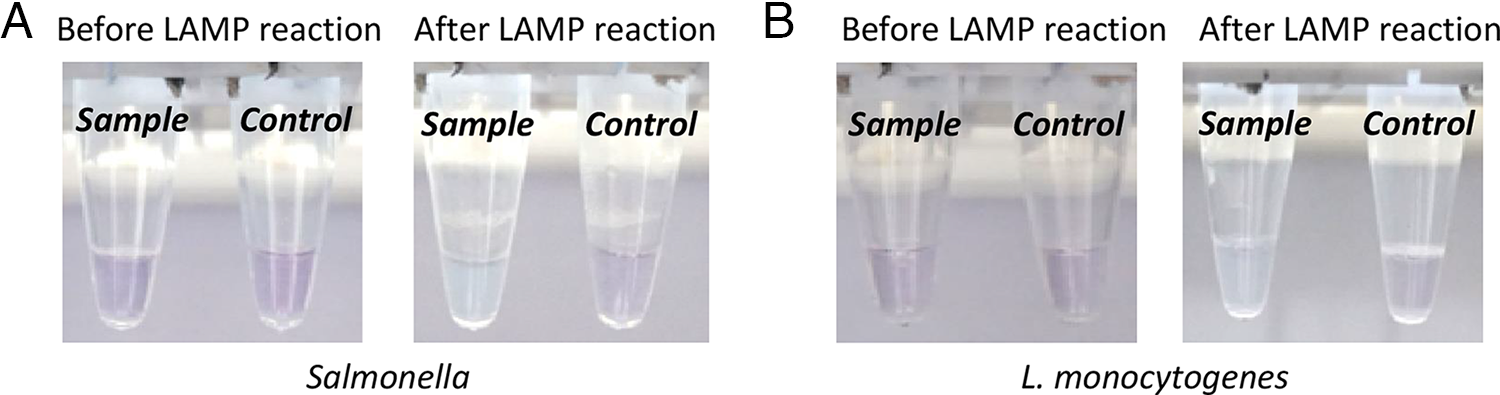
Colorimetric detection of LAMP products using HNB.
While this study focused on detecting single pathogens, future research could explore the development of multiplex LAMP assays for simultaneous detection of multiple foodborne pathogens. Such multiplex assays could further improve the efficiency and cost-effectiveness of food safety testing. In addition, further validation with a larger number of naturally contaminated samples would be beneficial to fully assess the performance of the assays in applications. Another potential area for improvement is the integration of these LAMP assays with rapid sample preparation methods. While the current study used a commercial DNA extraction kit, the development of simpler and faster DNA extraction protocols could further reduce the overall time required for pathogen detection.
Conclusions
The LAMP assays developed in this study provide a rapid, sensitive, and specific method for detecting Salmonella and L. monocytogenes in fresh beef. With optimized reaction conditions and colorimetric detection, these assays offer a promising tool for food safety monitoring.
Authors’ Contributions
X.W.: Methodology, software, validation, formal analysis, data curation, and writing—original draft preparation. K.J.: Methodology, software, validation, formal analysis, data curation, and writing—original draft preparation. J.R.: Methodology, software, validation, formal analysis, data curation, and writing—original draft preparation. X.Y.: Writing—review and editing. X.C.: Writing—review and editing. L.P.: Writing—review and editing. W.C.: Conceptualization and resources. R.F.: Supervision, project administration, funding acquisition, and writing—review and editing. Z.L.: Conceptualization, methodology, formal analysis, resources, project administration, funding acquisition, writing—original draft preparation, and writing—review and editing. All authors have read and agreed to the published version of the article.
Footnotes
Acknowledgment
The authors gratefully acknowledge all of the people who have contributed to this article.
Funding Information
This study was supported by the Fundamental Research Funds for the Central Universities (Project number: SWU-KR24016), National Center of Technology Innovation for Pigs (Project number: NCTIP-XD/C17), and Chongqing Modern Agricultural Industry Technology System (Project number: CQMAITS202312).
Data Availability
Data might be available upon request to the corresponding author.
Disclosure Statement
The authors have no conflicts of interest to declare.
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.