
Abstract
Using 10 Y-chromosome short tandem repeat allelic and haplotypic frequencies, we examined genetic variation within the population of Corsica and its relationship with other Mediterranean populations. The most significant finding is the high level of genetic differentiation within Corsica, with strong evidence of an effective barrier to male-mediated gene flow between the south and the rest of the island. This internal differentiation most probably results from low exogamy among small isolated populations and also from the orography of the island, with a central mountain chain running the length of the island restricting human movement. This physical barrier is reflected not only in present-day intraisland linguistic and genetic differences but also in the relatedness of Corsican regions to other Mediterranean groups. Northwest and Central Corsica are much closer to West Mediterranean populations, whereas South Corsica is closer to Central-North Sardinia and East Mediterranean populations.
Introduction
C
This study investigates the genetic structure of Corsica and its relationship with other Mediterranean populations using male-specific markers. Corsica was selected because: (1) its island status makes it a suitable model to study internal processes of differentiation, (2) its orography has determined the major geographical, linguistic, and cultural barriers, and (3) its “genetic” location with respect to the wider Mediterranean Basin is still controversial. To achieve this aim we analyzed Y-chromosome-specific diversity using 10 microsatellites or short tandem repeats (STRs).
Materials and Methods
Blood samples were obtained after informed consent from 62 unrelated Corsican males from three regions of the island: Northwest Corsica (NWC), Central Corsica (CC), and South Corsica (SC). Ten Y-STR loci (DYS19, DYS389I, DYS389II, DYS390, DYS391, DYS392, DYS393, DYS437, DYS438, and DYS439) were genotyped using methods previously described (Ghiani et al., 2004). To certify the correct designation of the specific alleles, we applied and obtained a GEDNAP Proficiency Test for forensic DNA typing. Allelic and haplotypic frequencies were estimated by counting. Gene and haplotype diversity values, and estimates of the sampling variance, were computed with Arlequin software (Excoffier et al., 1992). STR haplotypes were assigned into haplogroups by means of the algorithm implemented in the Haplogroup Predictor (www.hprg.com/hapest5/) (Athey and Nordtvedt, 2005). We also compared 10-locus haplotypic frequency distributions between our samples and available data on Mediterranean Basin populations (Ghiani and Vona, 2002; Cakir et al., 2004; Ghiani et al., 2004; Lopez et al., 2004; Martin et al., 2004; Spiroski et al., 2005; Frigi et al., 2006; Turrina et al., 2006). Pairwise genetic distances among Corsica and compared populations were computed as a linearization of the Rst (or Φst) values (Slatkin, 1995), with consideration of the molecular differences between individual haplotypes, in addition to differences in haplotypic frequencies. A multidimensional scaling was constructed from genetic distances by Statistica 7 (www.statsoft.com).
Results and Discussion
The distribution of allelic frequencies in the three Corsican regions was all highly polymorphic, and gene diversity values differed considerably. Sixty-two Y-chromosomes scored for 10 STRs reveal 53 different haplotypes (Table 1). The most significant finding is the high level of genetic microdifferentiation within Corsica, with strong evidence of a remarkably effective barrier to male-mediated gene flow between SC and the rest of the island. Only two haplotypes are shared between the CC and NWC regions, and no Y-haplotype is shared between SC and the other regions. The haplotype diversity observed is shown in Table 1. One explanation for this sharp internal differentiation might, at least in part, relate to the marriage structure, which characterized even the recent past in Corsica. Ranque et al. (1961) analyzed intraisland and extraisland matrimonial movement and determined that the exogamy rates between the NWC and SC were zero. A similar north-south difference is evident from a study of autosomal STRs by Tofanelli et al. (2001). The authors attributed the regional difference to the subdivision of Corsica into small isolates and to the presence of a NE-SW barrier to gene flow, resulting from a combination of geographic, historic, and linguistic barriers. A surname analysis over the whole island (Morelli et al., 2002) reached similar conclusions. The isonymy kinship values showed two main clusters, one comprising northern villages and one encompassing villages of the south. Corsica is divided into two distinct physical regions separated by a mountainous range: Banda di Fuori, to the west, and Banda di Dentro, to the east.
To identify which external Mediterranean populations, if any, could have influenced this internal differentiation of Corsica, we compared our data with those from other Mediterranean populations. Significant Rst values were observed between SC and all compared populations, whereas CC and NWC were undifferentiated from the populations of Italy, the Basque country, Spain, and the Pyrenees. The multidimensional scaling plot (Fig. 1) shows first the strong separation of NWC/CC from SC, and second it suggests the presence of two main groups in the Mediterranean: (i) a Western cluster, including NWC, CC, and South Sardinia and (ii) a cluster comprising Central/North Sardinia, SC, Turkey, and Sicily. These results receive support from the analysis of Y-haplogroup prediction (Table 1) that found R1b, the most frequent in NWC/CC (71% and 65%, respectively). This haplogroup is thought to have originated in Southwestern Europe, where it reaches its present-day highest frequencies (Semino et al., 2002). Conversely G2a was found (Table 1) to be the most frequent (43%) in SC. The distribution of Hg G in Europe may reflect the migration of agricultural Anatolians into the Mediterranean (Cinnioğlu et al., 2004). Contact with other populations was more frequent for inhabitants of SC, which historically lies more centrally in the path of the major commercial trade routes of the Mediterranean, than for other inhabitants of the island. Whether they came as invaders or traders, Greeks, Romans, and Byzantines in the early stages of Corsica's colonization most probably brought and then increased the frequency of the Hg G2a.
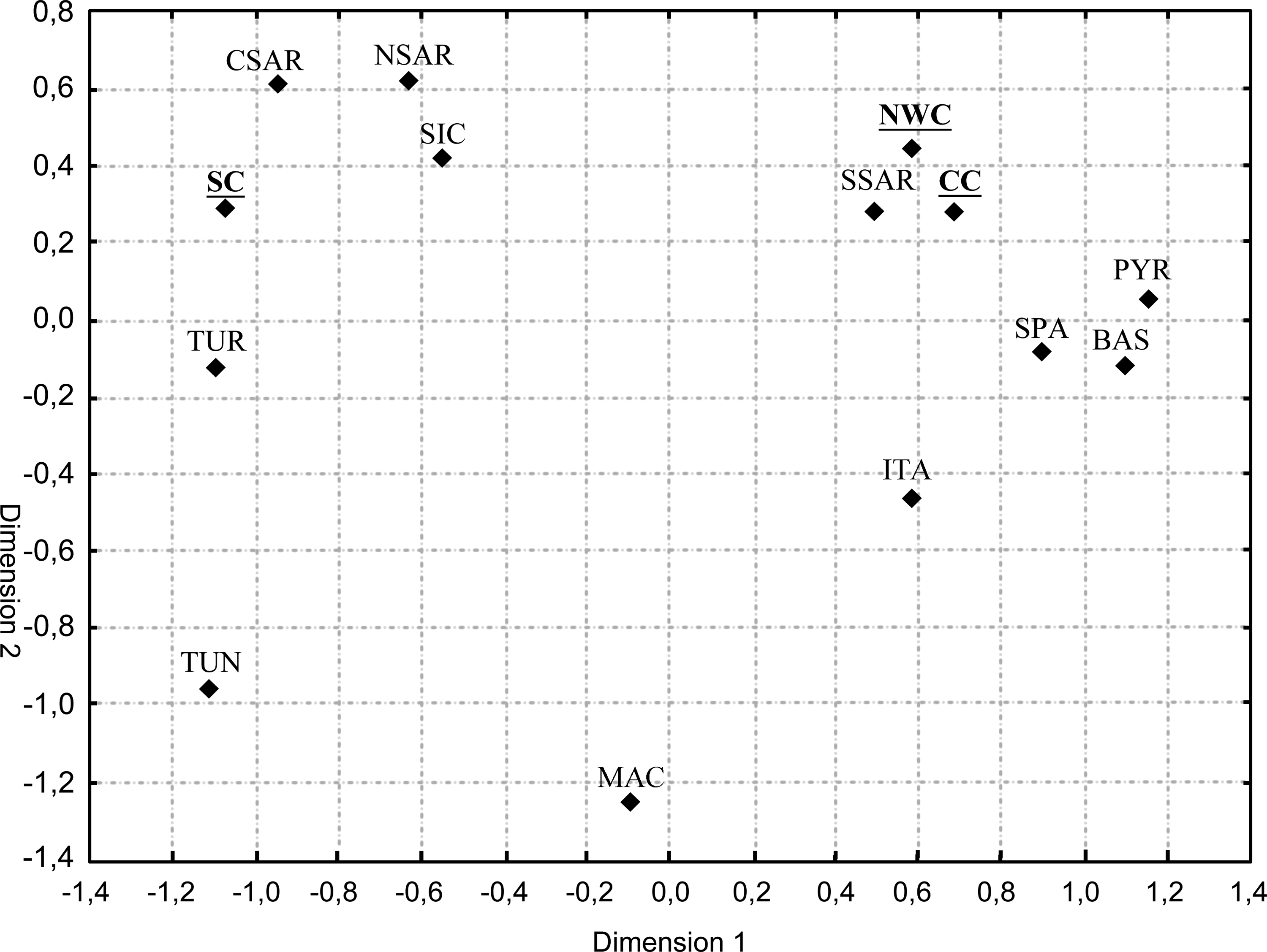
Multidimensional scaling, based on linearized Φst distances connecting Corsican and compared populations. SC, South Corsica; CC, Central Corsica; NWC, Northwest Corsica; CSAR, Central Sardinia; NSAR, North Sardinia; SSAR, South Sardinia; SIC, Sicily; TUN, Tunisia; ITA, Italy; BAS, Basques; TUR, Turkey; SPA, Spain; PYR, Pyrenees; and MAC, Macedonia.
In conclusion, our analysis of the Corsican male population indicates that the contemporary intraisland genetic differentiation most probably arose as a result of two factors. First, the long-term subdivision of the population into small isolates with very low levels of exogamy and gene flow, between even quite close settlements. Second, the effect of the NW-SE mountain chain which imposed a significant barrier to human movement and also influenced the geography of historical and administrative boundaries and linguistic differentiation.
Footnotes
Acknowledgments
This research was supported by grants from the University of Cagliari 60% (G.V.) and by the program INTERREG III (G.V. and L.V.)
Disclosure Statement
No competing financial interests exist.