
Abstract
Aims: The present study was conducted on two tribal communities, the Oraon and Munda, inhabiting the Ranchi district of Jharkhand state, India. The study was designed to elucidate genetic similarity, if any, shared between these tribes as they belong to the common Proto-Australoid stock but bear different linguistic affiliations. For this, a total of 98 intravenous blood samples (48 Oraon and 50 Munda) were collected from unrelated individuals of either sex up to first cousins, with their prior informed written consent. The DNA was extracted and studied for a total of 20 autosomal markers, including 7 Alu Indels, 3 DRD2 TaqI sites, 3 β-globin sites, and 7 restriction site polymorphisms.Results: All the 20 studied molecular markers were found to be polymorphic in both the tribal population groups and showed similarities with respect to allele frequencies, with a low coefficient of gene differentiation (GST) value. Moreover, sharing and distribution patterns of haplotypes of the β-globin gene cluster suggest that the Oraon and Munda share a common ancestry. However, small differences between them with reference to the linkage disequilibrium (LD) pattern indicate that the Munda might have emerged as a result of admixture between Proto-Australoids and Austro-Asiatic-speaking Mongoloids as supported by the principal co-ordinate analysis, wherein the Munda are closely placed with the Dravidian-speaking Proto-Australoid tribes of India.Conclusion: A common genetic substratum (Proto-Australoid stock) of the Oraon and Munda was evident in the present study, although these tribes are distinct linguistically.
Introduction
I
Human evolution has often been found at cross-roads in narrowing down to a specific phylogenetic pathway that is universally accepted from the two propositions, namely “out-of-Africa” (Cavalli-Sforza et al., 1988; Cavalli-Sforza, 1991; Vigilant et al., 1991; Wilson and Cann, 1992; Aiello, 1993) and “multiregional” (Thorne and Wolpoff, 1992; Frayer et al., 1993) theories. Keeping India in view, two major routes responsible for its initial peopling have been projected; first, from Africa, via Central Asia, to Northeast Asia, which then expanded toward Southeast Asia and to the islands beyond, that is, “the northern route” (Cavalli-Sforza et al., 1994), and the second from Africa through India to Southeast Asia and beyond, that is, “the southern route” (Nei and Ota, 1991; Chu et al., 1998; Su et al., 1999; Majumder, 2001; The HUGO Pan-Asian SNP Consortium, 2009). It has been proposed by Ballinger et al. (1992), Diamond (1997), and Gadgil et al. (1998) that the Austric language originated in China, from where it entered the Indian subcontinent through the northeast corridor and then diverged out its paths toward the islands. This is corroborated by the scattered distribution of the Austro-Asiatic speakers in India, particularly as pockets in the central and eastern regions.
The present study was designed to gain an insight into the peopling of India, specifically in relation to the several contending theories as to whether the Austro-Asiatic- or Dravidian-speaking tribal groups were the original inhabitants of the country.
For this, a total of 20 autosomal molecular markers were studied to provide a comprehensive view of genetic diversity and differentiation among Austro-Asiatic-speaking Munda and Dravidian-speaking Oraon inhabiting the eastern Indian state of Jharkhand.
Materials and Methods
The Oraon and Munda, the two tribal population groups of the Ranchi district (Jharkhand), were selected for the present study; the former belongs to Dravidian linguistic family, speaking the Kurukh dialect, whereas the latter belongs to the Austro-Asiatic linguistic family, speaking the Mundari dialect. For this work, 5 mL intravenous blood sample was drawn by a trained medical technician from each of a total of 98 unrelated individuals (40 Oraon and 50 Munda) up to first cousins of either sex, with their prior informed written consent. The fieldwork was conducted in 20 villages from three blocks, namely Kanke, Mandar, and Ratu of the district (Fig. 1). As the study involved blood collection from humans, prior approval for conducting the study was obtained from the Ethical Committee of the Department of Anthropology, University of Delhi, Delhi.

Map of India showing the geographical location of Ranchi district, Jharkhand.
Laboratory analysis
Genomic DNA was isolated from the collected blood samples using the salting-out method of Miller et al. (1988). Each DNA sample was typed for a total of 20 polymorphic autosomal sites, 7 Alu indel markers (PV92, FXIIIB, D1, APO, ACE, CD4, PLAT), 3 TaqI sites (A, B, D) of Dopamine D2 Receptor gene (DRD2), 3 sites of β-globin gene cluster [HBψβ (HB7), HB3′ψβ (HB8), HB5'β (HB9)], and 7 restriction site polymorphisms (ESR, LPL, T2, NAT, PSCR, ALAD, ADH2). The protocols used for Alu indel markers have been described by Stoneking et al. (1997), Majumder et al. (1999), and Watkins et al. (2001). The chromosomal locations, primer sequences, and polymerase chain reaction conditions for each restriction fragment length polymorphism studied are given in the Genome Base and listed by the Eccles Institute of Human Genetics (www.genetics.utah.edu/∼swatkins/pub/RSP_links.html). The protocols for DRD2 and β-globin markers are as mentioned by Jorde et al. (1995), Kidd et al. (1998), Majumdar et al. (1999), and Mukherjee et al. (2000). All the restriction site polymorphism markers were digested using 5 U of the appropriate restriction enzyme. The final product was then subjected to electrophoresis at 100 V on agarose gel. The results were noted and documented using a gel documentation system.
Statistical analysis
The allele frequencies of the selected markers were calculated by the gene counting method. The statistical software POPGENE version 1.32 (Yeh et al., 1999) was used to obtain heterozygosity and average heterozygosity estimates along with their standard deviations. Evaluation of the Hardy-Weinberg equilibrium was done by means of the chi-square (χ2) goodness-of-fit test and its statistical significance was considered at 5% level. Genetic differentiation was computed using DISPAN (Ota, 1993). The maximum likelihood estimates of haplotype frequencies were calculated from the multisite marker typing data using HAPLOPOP (Majumdar and Majumder, 1999). The coefficient of standardized pairwise linkage disequilibrium D′ (Lewontin, 1964) and the corresponding χ2 values were calculated with the LD software (Hill, 1974). Principal co-ordinate (PCO) analysis was performed using data on 17 common markers (PV92, FXIIIB, D1, APO, ACE, PLAT, 3 beta globin sites [HB7, HB8, HB9], 3 DRD2 TaqI sites [A, B, D], ESR, LPL, T2, NAT, PSCR). The software PCO version 2 (www.cse.naro.affrc.go.jp/iwatah/others/pco/index.html) was used to graphically represent the extent of genetic affinities among the population groups under study and those of the other reported Indian population groups (Chakrabarti et al., 2002; Vishwanathan et al., 2003; Saraswathy et al., 2008, 2009a). The relative amount of gene flow experienced by the Indian populations was estimated using the island model of Harpending and Ward (1982) and data from other populations on 17 common markers as mentioned before (Chakrabarti et al., 2002; Vishwanathan et al., 2003; Saraswathy et al., 2008, 2009a).
Results
All the 20 autosomal markers were found to be polymorphic in both the tribal population groups. Of these, markers each in the Oraon (FXIIIB, D1, CD4, PLAT, DRD2 TaqI D, HB9, ALAD) and Munda (FXIIIB, ACE, CD4, DRD2 TaqI A, HB7, T2, ADH2) exhibited a significant deviation from the Hardy-Weinberg equilibrium (Table 1). The frequencies of the autosomal markers have been tabulated along with their standard errors, which do not exceed the value of±2.0, indicating that the present tribes were in genetic equilibrium and deviations observed were just by chance (Table 1). In general, similar heterozygosity values were found at different loci for both the Oraon and Munda, albeit the latter tribe was found to have a relatively higher average heterozygosity (0.427) compared with the former tribe (0.407).
p-Value is indicative of the significant deviation from Hardy-Weinberg equilibrium.
To study genic differentiation, GST values were calculated, which show great heterogeneity over 20 marker loci. The average GST value obtained (0.01805) was low, which may be attributed to little genetic differentiation between the Oraon and Munda tribes (Table 2).
Of the eight possible β-globin haplotypes (prepared in the sequence HB9-Hb8-Hb7, each site having two alleles, with “+” allele indicating presence of site and “−” allele indicating absence of site), six were found among the Oraon and seven among the Munda (Table 3). The +−− haplotype was the most preponderant among both the Munda (48.2%) and Oraon (38.6%). For three of the haplotypes, +++, ++−, and +−+, the Oraon have nearly the same percentage of distribution, that is, 15.5%, 15.0%, and 15.4%, respectively. The standardized pairwise linkage disequilibrium (D′) values measured for the three sites, namely HB7, HB8, and HB9, of the beta globin gene (Table 4) were found to be low, that is, below both the tribal population groups.
Haplotypes were prepared with beta-globin sites in the order HB9-HB8-HB7.
Each site has two alleles—“+” allele indicating presence of site and “−” allele indicating absence of site.
Statistical significance at 5% level.
The PCO scatterplot (Fig. 2) shows both the Oraon and Munda in relatively close proximity, but the former tribe tends to form a cluster with the Dravidian populations of Andhra Pradesh and Nilgiri Hills (Tamil Nadu). The plot also shows that the populations from Northeast India form a separate cluster, although the Meitei of Manipur are found to be closer to the bigger cluster comprising the Oraon, Munda, and populations from Andhra Pradesh and Nilgiri Hills. Both the Mizo and Toda can be found at the opposite ends of the plot, none of them being close to any of the two major clusters.

Principal co-ordinate scatterplot based upon 17 autosomal markers exhibiting genetic affinities among the Oraon and Munda and another 16 population groups from South India and Northeast India.
Discussion
The Oraon and Munda tribes show similarity with respect to allele frequencies of the molecular markers studied with a very low GST value. Moreover, sharing and distribution patterns of haplotypes of β-globin gene cluster and also of DRD2 locus (Sachdeva et al., 2010) suggest that they share a common genetic ancestry. The difference found between them is only in LD patterns at both β-globin and DRD2 loci—the Oraon show statistically nonsignificant low LD with respect to three β-globin sites and three DRD2 TaqI sites (Sachdeva et al., 2010), whereas the Munda exhibit a statistically significant LD at all these sites. According to Sawyer et al. (2005), subpopulations of the same geographical region are expected to show similar LD patterns. However, the observed differences between the present two tribes can be conveniently attributed to the admixture scenario among the Munda (Van Driem, 2006). This tribe with slight Mongoloid features and belonging to the Austro-Asiatic linguistic group is thus an example of the fact that languages (cultural attributes) travel faster with greater intensity when compared to the genes.
It has been postulated that India has acted as a major corridor for human migration from Africa to Southeast Asia through the northeast border as the major land route (The HUGO Pan-Asian SNP Consortium, 2009). However, a contrary view was proposed by Cordaux et al. (2004), Thangaraj et al. (2005), Macaulay et al. (2005), and Saraswathy et al. (2009a) that the northeastern border was more of a barrier than corridor, which allowed inflow of people from Southeast Asia but not the back migration. This finding is basically hinged on the genetic discontinuity in Northeastern Mongoloid and other non-Mongoloid Indian population groups (Saraswathy et al., 2009b). The northeastern land route seems not to be a very convenient and feasible one, with the Himalayas spanning the major portion of the route save the Manipur-Myanmar border. Moreover, the movement of people from South and Central India toward the northeastern border also had geographical (hills) and climatic (cold climate) limitations. Thus, the present data indicate that the Mundari-speaking Munda, with very small traces of Mongoloid features, was found to be genetically similar to the Dravidian-speaking Oraon. This suggests that Austro-Asiatic-speaking people entering India through the northeast border might have substantially contributed their language to certain groups of people in the Indian subcontinent, as is evident by the distribution of Austro-Asiatic speakers in small pockets of the central and eastern parts of India. In contrast, their genetic contribution seems to be limited as seen in the case of the Munda in the present study.
Interestingly, the Austro-Asiatic-speaking Khasi tribe of Meghalaya lives closer to the northeastern border of India and exhibits exclusively Mongoloid features. This could have been possible only when Austro-Asiatic speakers from Central China entered India through northeast border and maintained identity as Khasi or else they substantially contributed both language and genes to already existing people, resulting in the Khasi tribe. Thus, the Austro-Asiatic language, spoken widely in Southeast Asian countries by the people with Mongoloid features, is expected to have reached India later. On the contrary, an ethnic element, that is, the Proto-Australoid, is not only found in the majority of primitive and advanced tribal groups but also among some of the high-caste groups of India, indicating a basal genetic substratum of Indian subcontinent as also proposed by Saraswathy et al. (2010).
The Oraon, with typical Proto-Australoid features and speaking a Dravidian tongue, are expected to be closer to the Dravidian speakers of South India. The Munda present a very interesting situation that despite belonging to a different linguistic group, they are placed in between Dravidian and Austro-Asiatic speakers but are relatively closer to the Oraon, indicating their genetic proximity with Proto-Australoid Dravidian-speaking tribes of India. Another Austro-Asiatic speaking tribe, the Ho, is found to be closer to Dravidian populations as against Austro-Asiatic or Tibeto-Burman speakers. It is also evident that the Tibeto-Burman-speaking Meitei group of Manipur (Northeast India), with mongoloid features, also lies in genetic proximity of the major cluster as they inherit the Proto-Australoid stock (Saraswathy et al., 2009a). In contrast to previous studies that support the idea that linguistic affiliations coincide with genetic similarities (Barbujani and Sokal, 1990; Barbujani, 1991; Dupanloup de Ceuninck et al., 2000; Thangaraj et al., 2005; Majumder, 2010), the PCO analysis (Fig. 2) in the present study reveals that the Tibeto-Burman-speaking Meitei tribe, the Austro-Asiatic-speaking Munda tribe, and Dravidian-speaking tribes form a major cluster, suggesting that geographic placement and ethnicity bear greater impact on genes compared to language. In the centroid analysis, all Dravidian-speaking populations along with Austro-Asiatic-speaking Munda and Indo-European-speaking Rana Tharu indicate gene flow among them with higher heterozygosity (Fig. 3). This is suggestive of larger effective sizes of these populations in India. This conforms well with the already reported fact that South Indian populations have diversified sources of gene flow (Saraswathy et al., 2008).
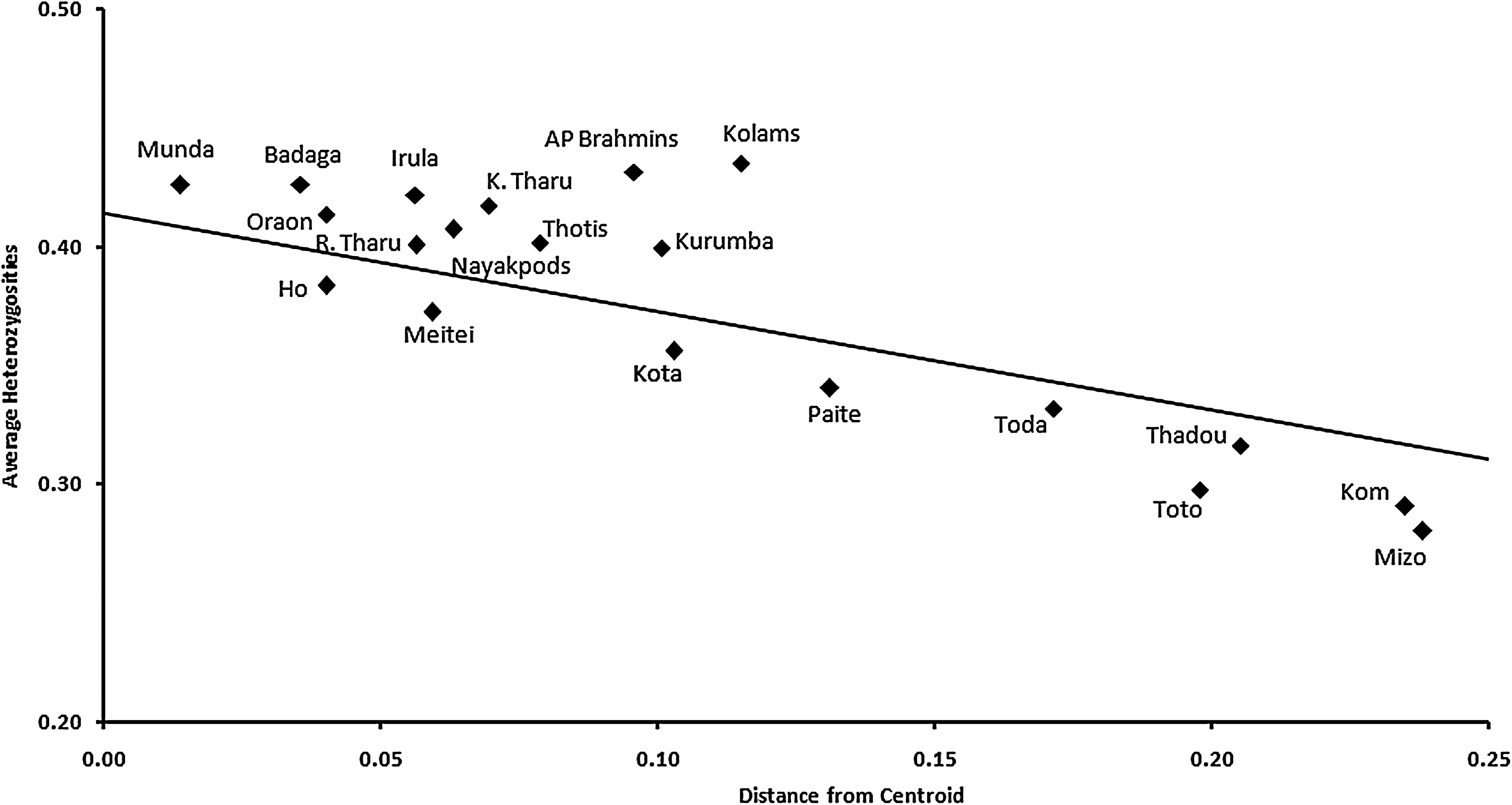
Centroid analysis using common 17 markers on the studied populations with other Indian populations.
Isolated populations showing lower heterozygosity values include the Austro-Asiatic-speaking Ho tribe, Tibeto-Burman-speaking Meitei, Paite, Thadou, Kom, Mizo, and Dravidian-speaking Kota and Toda tribes. From this, one can infer that isolation with respect to geography, language, and culture brought out homogeneity among populations, leading to distinctive gene pools. Thus, peopling of India may be attributed not only to autochthonous origin, and as a result of migration and admixture, but also to social rules such as strict endogamy prevailing in certain communities and areas.
Conclusion
Linguistically distinct Oraon and Munda of the Ranchi district are found to be genetically similar with respect to 20 autosomal genetic markers. Small differences between them with reference to the LD pattern indicate that the Munda might have emerged as a result of the admixture between Proto-Australoids and Austro-Asiatic-speaking Mongoloids. This is also evident from PCO analysis, wherein the Munda were closely placed with the Dravidian-speaking Proto-Australoid tribes of India. The present study also indicates a longer history of the Oraon in the region when compared with the Munda. Thus, a common genetic substratum (i.e., Proto-Australoid stock) of the two tribes is conspicuous in the present study for 20 autosomal loci.
Footnotes
Acknowledgments
The authors are thankful to the University Grants Commission for providing the financial support to conduct the present study. The authors are also grateful to the Department of Anthropology, University of Delhi, for providing the infrastructure to carry out the study.
Disclosure Statement
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the manuscript.